
Abstract
Despite an apparent increase in cases of angiostrongyliasis in humans and animals in Australia, the epidemiology of infection with the two species of Angiostrongylus that co-exist in this country, namely A. cantonensis and A. mackerrasae, is poorly understood. This knowledge gap is particularly important with respect to A. mackerrasae, a species evidently native to Australia, as its ability to cause disease in humans is unknown. Likewise, there is little information on the roles of native and introduced species of rodents and molluscs as hosts of Angiostrongylus species in Australia. This review focuses on the gaps in the knowledge about the two species, highlighting the need for epidemiological and pathogenesis studies on the native lungworm A. mackerrasae.
Introduction
N
Between 1994 and 2006, more than 300 cases of human angiostrongyliasis were reported in China (He et al. 2009). To date, over 2800 human cases have been reported worldwide, of which 28 were in Australia (Wang et al. 2008, Morton et al. 2013). Outbreaks of the disease have been reported in Tahiti (Rosen et al. 1961), American Samoa (Kliks and Palumbo 1992), Taiwan (Tsai et al. 2001), the Caribbean region (Lindo et al. 2002, Slom et al. 2002), Japan (Asato et al. 2004), China (Wang et al. 2008, Deng et al. 2011, Chen et al. 2011), and Ecuador (Dorta-Contreras et al. 2010). In such outbreaks, groups of people are usually affected by the disease in a single geographical location owing to a point source exposure attributed to contaminated foods. In contrast, reported cases from Australia have mostly been isolated incidents caused by ingestion of infected snail or slug tissues by children (Morton et al. 2013) or by misadventure, including “dares” by peers to ingest molluscs (Senanayake et al. 2003, Blair et al. 2013). A. cantonensis causes significant pathology, especially in infected children, and the prognosis of the disease is usually poor when considerable numbers of larvae are ingested (Li et al. 2001).
Other Angiostrongylus species known to cause pathology in animals and humans are A. vasorum, A. costaricensis, and A. malaysiensis. As with A. cantonensis, all three species use terrestrial and aquatic molluscs as their intermediate hosts. A. vasorum is found in Europe, northern Asia, North America, and Africa. Adults of this species infect the pulmonary arteries of dogs and wild canids, which are the definitive host. A. vasorum is of veterinary significance, but is not zoonotic (Morgan et al. 2005). A. costaricensis, found in Costa Rica, Brazil, and Venezuela (Morera and Cespedes 2002, Graeff-Teixeira et al. 2005, Incani et al. 2007), have adult worms living in the mesenteric arteries of their rodent host. Accidental ingestion of larvae by humans is known to cause severe enteritis, but not neurological disease, as seen by other species of this genus (Rebello et al. 2013). A lesser known species of rat lungworm, A. malaysiensis, occurring mainly in Malaysia, is also a cause of eosinophilic meningitis in humans (Lim and Mak 1983). Together with A. cantonensis, the former species of Angiostrongylus are classified as emerging zoonotic infections, particularly in endemic countries where terrestrial molluscs are used as a food source and rodent control is nonexistent.
Angiostrongylus Species in Australia
A few attempts have been made to define the geographical occurrence of species of Angiostrongylus within Australia, but the data are limited and specific clues as to how the parasites entered the continent are almost nonexistent. An Angiostrongylus species was first reported from Queensland as a parasite of rats by Mackerras and Sandars (1954). Although these authors identified the parasite as A. cantonensis, Bhaibulaya (1968), in his analysis of Australian angiostrongylids, re-examined the voucher specimens deposited by Mackerras and Sandars and proposed that the specimens belonged to a separate, new species, which he named A. mackerrasae. A. cantonensis is now a common parasite of feral rats in southeastern Queensland and in regions of Sydney in New South Wales (Prociv and Carlisle 2001).
Data on the northern limit of the parasite are scant. Dunsmore (1968) searched for lungworms in four species of rodents from the Sir Edward Pellew Islands (15′40″S, 136′49″E) in the Gulf of Carpentaria, northern Australia. Dunsmore did not recover any Angiostrongylus specimens from these hosts. It should be noted that Dunsmore's study focused on rat and mammal species belonging to genera other than Rattus, the natural hosts of A. cantonensis and A. mackerrasae. Although one species of the genus Rattus, R. sordidus, is found on the Edward Pellew Islands, Dunsmore did not investigate this species for parasites. The absence of positive records by Dunsmore (1968) may simply reflect his examination of mammals that are refractory to infection because of dietary preferences, natural immunity, or the host specificity of the lungworms.
Alicata (1988) inferred that Angiostrongylus arrived in Australia through the eastern part of the continent. There are no reports of Angiostrongylus from western and southern parts of Australia, and it has only been within the last decade that the parasite was found as far south as Jervis Bay in New South Wales (35.09′S 150.80′E) (Stokes et al. 2007).
Following Alicata, it appears that A. cantonensis may have been initially introduced to Queensland with infected rats, as indicated by the discovery of the parasite in rodents and early reports of infection in dogs and humans. The more recent discovery of infections in animals and humans in Sydney and the southern coast of New South Wales could suggest a recent southerly expansion in disease distribution of this parasite in Australia. This expansion of A. cantonensis distribution in Australia was possibly enhanced by low host specificity and the fact that most terrestrial molluscs and rats species in Australia can potentially serve as natural hosts for this nematode (Prociv et al. 2000). However, much remains to be discovered about host preferences of angiostrongylids in Australia, the feeding biology of native and introduced slugs, the distribution of the parasites throughout their known range, and the actual geographical limits of both species in Australia.
Angiostrongylus species have been reported from both feral and native rat species of Australia (Bhaibulaya 1968). Understanding the epidemiology of Angiostrongylus species in Australia is complicated by the presence of two nearly identical species, A. cantonensis in introduced Rattus species and A. mackerrasae in the native rat, Rattus fuscipes. Despite the similar life cycle of the two species, A. mackerrasae can be differentiated from A. cantonensis using morphological criteria of the adult worms. Bhaibulaya (1968) distinguished the two species on the sizes of the spicules in adult males, which in A. cantonensis are 1.24 mm in length and in A. mackerrasae are 0.49 mm in length (Fig. 1). In addition, the length of the vagina in the adult female parasites differs between the species, with that of A. cantonensis being longer. Because the primary points of morphological distinction between the two species are in the copulatory apparatus, it might be expected that there exists mechanical isolation between the two species. Species interbreeding has been achieved, however, when experimental hybridization of the two species produced male F1 hybrids that were sterile, whereas females remained fertile (Bhaibulaya 1974). Bhaibulaya (1974) also observed that the morphology of F1 hybrids were intermediate between the two species and, on the basis of this, suggested that the two species are distinct. This leads to the speculation that the two parasite species have the potential to become sympatric within the same host, possibly leading to the evolution of a hybrid species.
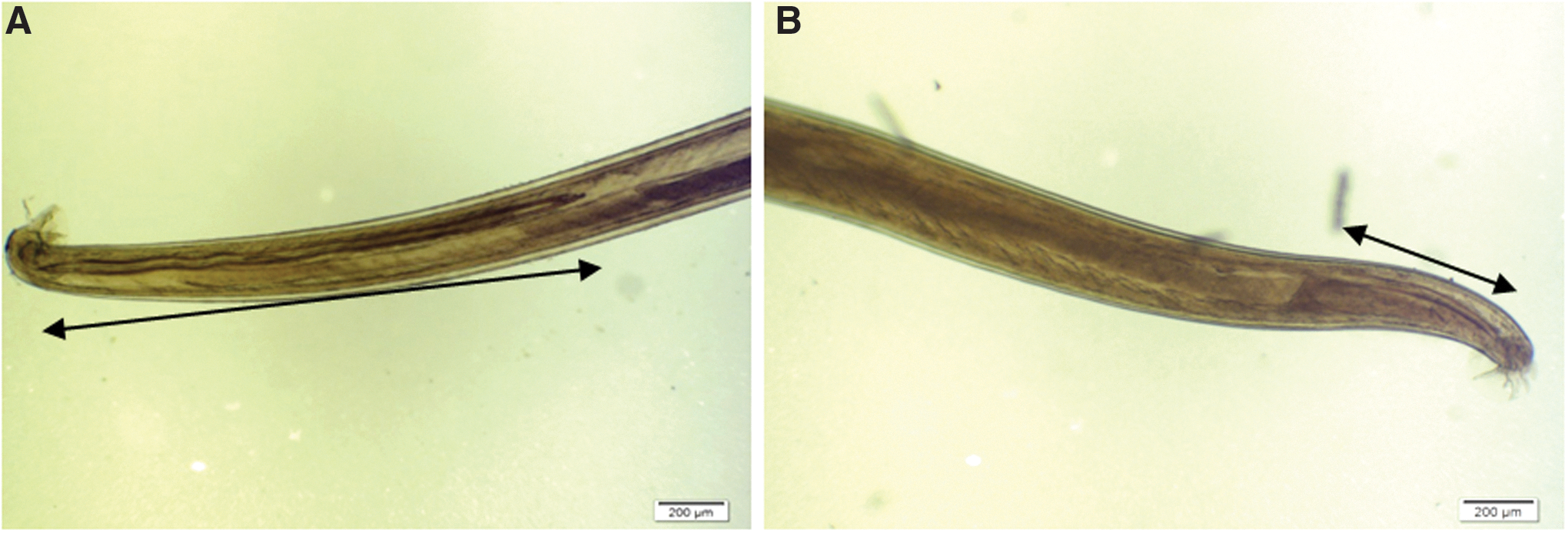
Difference in spicule size in adult male (arrows): The posterior end of adult male A. cantonensis (
Whereas A. cantonensis is known to be responsible for eosinophilic meningitis in a wide range of hosts, the role of A. mackerrasae in animal and human disease is entirely unknown. Forty years ago, Bhaibulaya (1974) showed that A. mackerrasae was neurotropic and migrated to the brain of rats in the same manner as A. cantonensis (Bhaibulaya 1974). Given the similarity in larval migratory pathways within the host of these two species, it is plausible that A. mackerrasae may also be neurotropic in humans as well as other animal hosts. Because morphological separation of the two species is almost impossible at all stages bar the fully developed adult worms, we hypothesize that recovered worms from humans and animals at post mortem may have a proportion that are A. mackerrasae. Recovered nematodes from human cases of angiostrongyliasis in Australia to date have not attempted to identify the etiological agent by genetic analysis.
Although angiostrongyliasis is clearly enzootic in Australia (Prociv and Carlisle 2001), there have been relatively few human cases reported, raising the question as to whether human infections are either misdiagnosed or underreported. At worst, infection with A. cantonensis can lead to eosinophilic meningoencephalitis, pulmonary abscesses, blindness, profound mental retardation, spasticity, epilepsy, and even death in infants who ingested infected planarians or snails (Cooke-Yarborough et al. 1999, Morton et al. 2013). However, the most frequent symptom in humans is severe chronic headache for which patients are rarely hospitalized (Prociv et al. 2000), and therefore many cases may be missed.
There exists only scattered epidemiological information of Angiostrongylus spp. along the eastern regions of Australia. Local surveys in the state of Queensland (Yong et al. 1981) showed that A. cantonensis occurred in populations of feral rats in pockets along the Brisbane River, which runs through the capital city of Brisbane. In addition, retrospective case studies in dogs provide evidence that A. cantonensis is widespread in metropolitan areas of Brisbane and Sydney (Mason et al. 1976, Lunn et al. 2012). According to Yong and colleagues (1981), virtually all terrestrial molluscs in that region could be infected experimentally. In another survey done by Stokes et al. (2007), the prevalence of rats infected with A. cantonensis in coastal forests of the regions surrounding Jervis Bay, New South Wales, in southeastern Australia was reported to be 11.3% in summer, with a prevalence of up to 14.7% in selected sites. Unfortunately, neither study used molecular methods to identify of Angiostrongylus species, although the authors indicated that both Australian species were present in the survey site.
Apart from rats and dogs, A. cantonensis has also been reported in many different animal species in Australia, including captive Bennett's wallabies (Macropus rufogriseus) (McKenzie et al. 1978), horses (Wright et al. 1991), captive rufous bettongs (Aepyprymnus rufescens) (Higgins et al. 1997), captive tamarins (Sanguinus spp.) (Carlisle et al. 1998), wild black- and grey-headed flying foxes (Pteropus species) (Barrett et al. 2002), Yellow-Tailed Black Cockatoos (Calyptorhynchus funereus) (Monks et al. 2005), and, most recently, Tawny Frogmouths (Podargus strigoides) (Monks et al. 2005, Gelis et al. 2011, Ma et al. 2013) and brushtail possums (Trichosurus vulpecula) (Ma et al. 2013). The presence of A. cantonensis has also been suggested to be a threat to Australian endangered bird species, such as the Eastern Bristlebird, Dasyornis brachypterus, which prey on gastropods (Stokes et al. 2007).
Interestingly, there is a recent report of a patent infection of the flying fox Pteropus alecto with the native angiostrongylid A. mackerrasae (Mackie et al. 2013). This finding has important implications for our understanding of animal and human angiostrongyliasis. Despite the serious risk of Angiostrongylus spp. to wildlife and public health, no systematic survey of this nematode has ever been conducted in Australia. A national survey of this nematode has been carried out in China (Zhang et al. 2009), and studies such as these are advocated in Australia to determine the prevalence, geographical distribution, and climatic and ecological drivers of this parasite in definitive and intermediate hosts to identify populations at risk.
Rats As Definitive Hosts in Australia
Since its first description from R. norvegicus in China (Chen 1935), A. cantonensis has been reported from other native and introduced rat species from Southeast Asia and Australia (Prociv et al. 2000) such as R. rattus (Ash 1962), R. fuscipes (Bhaibulaya 1968), R. exulans (Carney et al. 1978, Uchikawa et al. 1984), R. flavipectus (Zhang et al. 2009), R. argentiventer (Vitta et al. 2011), and R. losea (Deng et al. 2012). In Australia, A. cantonensis has been reported from R. rattus and R. norvegicus, whereas A. mackerrasae has only been reported from native rat species, R. fuscipes (Bhaibulaya 1968) and R. lutreolus (Stokes et al. 2007).
Stokes et al. (2007) described complete geographical separation of A. cantonensis and A. mackerrasae in Jervis Bay forests. In that study, it was found that A. cantonensis was only present in R. rattus populations in northern forests of the region and A. mackerrasae was only observed in R. fuscipes in southern forests. Because the life cycles of A. cantonensis and A. mackerrasae are very similar, there is a potential for mixed infections in native rat species as well as in introduced rats.
The close proximity of populations of introduced black rats and native rat species (e.g., bush rats, R. fuscipes) (Banks and Hughes 2012) and their shared habit of mollusc predation raises some interesting possibilities. Members of the genus Rattus are thought to have invaded Australia from New Guinea about 1.4 million years ago, with species descended from the original invaders belonging to the R. fuscipes group of species (Robins et al. 2010). More recent invasions (approximately 0.5 million years ago) from the R. sordidus and R. leucopus lineages led to greater diversification of members of the genus on mainland Australia. Despite the large radiation of Rattus species in Australia, it appears that A. mackerrasae is the only species found in native Australian rats. The geographic range of this species appears to be limited by the natural range of its preferred host, R. fuscipes. The presence of only a single native Angiostrongylus is intriguing because there is a diverse range of native Rattus species in Australia and an even more diverse range of species in New Guinea and South East Asia countries (Robins et al. 2014). Several other native rodent species occur in this country that frequently eat molluscs or other invertebrate prey (e.g., Hydromys chrysogaster and Uromys caudimaculatus).
Introduced rodents that occur on the Australian mainland and on some offshore islands include the house mouse (Mus musculus), the brown rat (R. norvegicus), the black rats (R. rattus) and Asian rat (R. tanezumi). Another inhabitant of islands, the Pacific rat, R. exulans, appears to have extended its range with prehistoric human migration and trading (Matisoo-Smith et al. 1998). These introduced rodents occur as commensal species (that is, they live within close proximity to humans), with the house mouse and Pacific and black rats also occurring in natural environments. Populations of these introduced rodents do not fluctuate as much as those of the native rats, because of their proximity to human habitations and the consequent static environment in which they live.
It is not clear whether the introduced rat species can act as reservoirs for A. mackerrasae. Experimental work performed by Bhaibulaya (1974) suggests that laboratory rats can support this species, but whether transmission of the parasite to different hosts occurs in nature has not been determined. Furthermore, there is some confusion surrounding the genetic identity of rats in Australia. Genetic analysis indicates that the Australian black rat population is comprised of two species R. rattus and R. tanezumi, which cannot be distinguished on morphological grounds (Dickman and Watts 2008). Any investigation of A. cantonensis in black rats in Australia has failed to consider this complicating factor. Finally, as stated before, an understanding of the epidemiology and host ranges of the two species of Angiostrongylus in Australia is limited by the discrete morphological differences between the two parasite species, and the complete absence of genetic data for A. mackerrasae.
In view of the dearth of useful genetic and morphological data of these parasites and their hosts, and the continuing infections of humans and domestic and native animals, there is need for better epidemiological data on these parasites. There has been no systematic survey of the angiostrongylids in Australia to identify the geographical range or hotspots of epidemiological significance. Identification of hotspots for the transmission of A. cantonensis, and possibly A. mackerrasae is one of the most effective ways of controlling the disease occurrence. For instance, application of rodent control tools at Taronga Zoo, Sydney, New South Wales, has dramatically reduced the cases of neural angiostrongyliasis in zoo animals (Dr. K. Rose, personal communication).
Sentinel Hosts for Hotspots of Human Infection
Apart from the indication of a broad rat survey, it has been proposed that dogs can be used as sentinels for disease transmission in the urban and periurban areas (Lunn et al. 2012). Lunn and colleagues noted that the majority (64%) of dogs diagnosed with neural angiostrongyliasis in Sydney occurred between the autumn and winter months from April to July. Presumably, the cool and damp nights of this season allow terrestrial molluscs to emerge and seek food. Pups and adolescent dogs are more commonly infected and show some attraction to slugs (Mason 1987). The combination of this exploratory behavior by younger dogs as well as accidental ingestion of slugs that may contaminate dog food left outside the house are likely sources of infections of domestic dogs (Lunn et al. 2012). In addition, the infection of the two toddlers in Sydney also occurred during the autumn and winter period (Morton et al. 2013), thus supporting a seasonal prevalence. Therefore, more information on the distribution of both species found in Australia and environmental risk factors influencing infection to humans and animals are required. For such a comprehensive study to be effective, better detection methods in both the warm-blooded host and the terrestrial molluscs must be developed.
A relatively common Australian bird, the Tawny Frogmouth (Podargus strigoides) has also been suggested as a sentinel to identify risk areas for human and animal infections (Ma et al. 2013). The diet of these birds consists mostly of large nocturnal invertebrates. Juvenile birds are prone to hunting and ingesting snails, and a series of case reports identified a cluster of infections in this species (Gelis et al. 2011). A recent survey showed that 80% of cases of Tawny Frogmouths with neurological disease had nematodes in the lesions, and, in the majority of those cases, the infection was attributed to A. cantonensis infection (Ma et al. 2013). Most cases of diseased birds occurred in a period from February to June, a time, as stated, when terrestrial molluscs are likely to be active.
Significance of Human Disease in Australia
Terrestrial molluscs are not commonly eaten as food in Australia. Due to their behavior, young children and animals are most at risk of infection with this insidious parasite. Because of the widespread presence of Angiostrongylus spp. in urban environments of eastern Australia, the diagnosis of this potentially fatal disease should be considered in any patient with severe headache, paraesthesia, or cranial nerve palsy (Alto 2001).
The recently reported human cases in New South Wales raises concern about the emergence of neural angiostrongyliasis in eastern Australia. These infections occurred in two toddlers, aged 10 and 14 months, from two relatively close neighborhoods in Sydney. One of these children subsequently died from the infection (Morton et al. 2013). Other cases reported in Sydney where two young men who developed eosinophilic meningitis (Senanayake et al. 2003, Blair et al. 2013). One patient recovered after 5 months after infection but the other was left with permanent central nervous system damage 18 months later. The cases highlight the need for ongoing awareness of A. cantonensis as an infectious risk in urban areas of eastern Australia and a need for rapid diagnosis of infection with this species. With continued incursion of human habitation onto forested areas in periurban regions, the presence of the sylvatic A. mackerrasae is an additional potential risk. The recent report of patent lung infection from a flying fox (Pteropus alecto) caused by A. mackerrasae indicated that this species has the potential to cause cross-species infections and to cause similar pathogenesis to that caused by A. cantonensis (Mackie et al. 2013). The findings of Mackie and colleagues (2013) emphasizes the need for a better understanding of the identity of agents of angiostrongyliasis in Australia and the potential of the two Australian members of the genus to cause diseases in humans and their pets.
Diagnosis of Angiostrongylus infections in humans and domestic animals is difficult due to low specificity and sensitivity of available tools (Lunn et al. 2012). Serological diagnosis has been made difficult by cross-reactivity with other helminth infections such as Gnathostoma species, Strongyloides stercoralis or filarial nematodes (Chye et al. 2000, Senthong et al. 2013). Enzyme-linked immunosorbent assay (ELISA) and western blot analysis of cerebrospinal fluid shows more promise, with higher positivity in cases in dogs and humans (Lunn et al. 2012). Nonetheless, some humans do not seroconvert in the early stages of infection, leading to failure to detect infection by serological methods (Morton et al. 2013); the late detection by antibody tests will lead to delayed administration of crucial anti-inflammatory medication. This delay will subsequently allow further pathological changes to occur as the parasite grows and continues to migrate through the central nervous system. Research should now focus on early detection of parasite antigen and/or genetic material in serum or in cerebrospinal fluid.
A number of laboratories have developed tests that have been applied to the investigation of A. cantonensis infections in snails. The tests target the ITS1 region of ribosomal DNA of the nematode (Qvarnstrom et al. 2007, Qvarnstrom et al. 2010, Jarvi et al. 2012) and showed relatively high sensitivity. However, the reliability of the test in human cerebrospinal fluid or serum depends on the availability of parasite DNA in active or resolving infections. Because the parasites do not replicate in the host, DNA abundance is entirely related to the numbers of parasites that enter the meninges or migrate through the ventricles.
Conclusion
Neural angiostrongyliasis in Australia remains a sporadic infection with potential lethal consequences in warm-blooded animals. Its introduction and spread into the eastern regions of Australia remain speculative. A. cantonensis is likely to have been brought to Australia with the introduction of R. rattus. The sharing of intermediate and definitive hosts for A. cantonensis and A. mackerrasae may provide opportunities for a hybrid species to develop that could have a different pathogenicity in native animal populations. Thus, there is a need to understand the population genetics of the two sympatric Angiostrongylus spp. present in Australia and determine whether the two species have potential to develop hybrid progeny that could establish and be more pathogenic.
Angiostrongylus infections appear to be emerging in many countries, and the association of several species with human diseases is becoming increasingly recognized. This is also the case in Australia where two similar species occur and both are neurotropic in their definitive hosts. Furthermore, the availability of snail intermediate hosts is dependent on environmental factors such as rainfall and temperature; this would indicate that Angiostrongylus spp. are likely to be seasonal in their prevalence.
Currently, available diagnostic tools for angiostrongyliasis in Australia are built solely around recognition of A. cantonensis. The absence of reagents for identifying A. mackerrasae raises two problems. The first is that the current probes might not detect the potentially pathogenic A. mackerrasae in serum or cerebrospinal fluid samples. This lack of diagnostic specificity and sensitivity could lead to misdiagnosis and underestimation of the true incidence of angiostrongyliasis in Australia. Second, because Australia is an island continent, the isolation of Angiostrongylus in this country may result in genetic divergence. Identification of transmission hotspots in Australia and other endemic areas is essential to aid in clinical diagnosis of neural angiostrongyliasis in humans and other animals.
Footnotes
Acknowledgments
Mahdis Aghazadeh is supported by a University of Queensland Postgraduate Award. Work leading to this review was supported by grants from the ANZ Queensland Community Foundation–Peter and Mary Ellen Stone Memorial Fund awarded to Malcolm Jones.
Author Disclosure Satement
No competing financial interests exist.