
Abstract
Rift Valley fever virus (RVFV), a member of the Bunyaviridae family, is a mosquito-borne zoonotic pathogen that causes serious morbidity and mortality in livestock and humans. The recent spread of the virus beyond its traditional endemic boundaries in Africa to the Arabian Peninsula coupled with the presence of susceptible vectors in nonendemic countries has created increased interest in RVF vaccines. Subunit vaccines composed of specific virus proteins expressed in eukaryotic or prokaryotic expression systems are shown to elicit neutralizing antibodies in susceptible hosts. RVFV structural proteins, amino-terminus glycoprotein (Gn), and carboxyl-terminus glycoprotein (Gc), were expressed using a recombinant baculovirus expression system. The recombinant proteins were reconstituted as a GnGc subunit vaccine formulation and evaluated for immunogenicity in a target species, sheep. Six sheep were each immunized with a primary dose of 50 μg of each vaccine immunogen with the adjuvant montanide ISA25; at day 21, postvaccination, each animal received a second dose of the same vaccine. The vaccine induced a strong antibody response in all animals as determined by indirect enzyme-linked immunosorbent assay (ELISA). A plaque reduction neutralization test (PRNT80) showed the primary dose of the vaccine was sufficient to elicit potentially protective virus neutralizing antibody titers ranging from 40 to 160, and the second vaccine dose boosted the titer to more than 1280. Furthermore, all animals tested positive for neutralizing antibodies at day 328 postvaccination. ELISA analysis using the recombinant nucleocapsid protein as a negative marker antigen indicated that the vaccine candidate is DIVA (differentiating infected from vaccinated animals) compatible and represents a promising vaccine platform for RVFV infection in susceptible species.
Introduction
R
RVFV belongs to the genus Phlebovirus within the family Bunyaviridae, which includes over 350 named isolates (Walter and Barr 2011). It has a tripartite single-stranded RNA genome of negative polarity consisting of small (S), medium (M), and large (L) RNA segments. The M segment encodes two structural glycoproteins, amino-terminus glycoprotein (Gn) and carboxyl-terminus glycoprotein (Gc), the 78-kDa protein, and the nonstructural protein, NSm, whereas the S segment encodes the nucleocapsid protein (N) protein and the nonstructural protein NSs. The L segment encodes an RNA-dependent RNA polymerase (Elliott 1996). The N and L proteins are required for viral RNA synthesis. The NSs protein, the major virulence factor, has been shown to inhibit host transcriptional immune response (Bouloy et al. 2001) through generalized transcription downregulation, including repression of interferon-β (IFN-β) (Billecocq et al. 2004, Le May et al. 2004, Le May et al. 2008) and degradation of protein kinase R (Habjan et al. 2009, Ikegami et al. 2009). The NSm protein functions to suppress virus-induced apoptosis (Won et al. 2007), and there is evidence that it plays a functional role in the vector competence of mosquitos for RVFV at the level of the midgut barrier (Kading et al. 2014). The glycoproteins Gn and Gc are surface proteins that play a role in virus attachment to initiate infection and have been shown to carry epitopes that elicit the production of neutralizing antibodies, a correlate of protective immunity (Besselaar et al. 1991, Besselaar and Blackburn 1992, Besselaar and Blackburn 1994).
There are currently no RVFV vaccines fully approved for commercial use outside its endemic area in Africa and the Arabian Peninsula. Given the potential for viral spread elsewhere, including the mainland United States, there is an urgent need for a safe and efficacious vaccine. Attributes essential for a vaccine for use in nonendemic areas include safety and the ability to generate a rapid (with primary vaccination) protective immune response in a susceptible host. In endemic regions, RVFV in livestock has been controlled traditionally by using the live-attenuated Smithburn strain or inactivated whole virus (Grobbelaar et al. 2011). The Smithburn vaccine is highly immunogenic, but it is teratogenic in pregnant sheep and cattle (Coetzer and Barnard 1977, Botros et al. 2006). The formalin-inactivated whole-virus vaccines are safe but less immunogenic (Lubroth et al. 2007). Clone 13, another live attenuated natural mutant, is now in commercial use in South Africa for the control of the disease in livestock (Muller et al. 1995, Dungu et al. 2010). MP12, a chemically attenuated virus derived from ZH548, an Egyptian wild-type isolate (Caplen et al. 1985, Vialat et al. 1997), is being evaluated as a potential vaccine for human and veterinary use. The immunogenicity and pathogenicity of these latter two candidate vaccines have been evaluated in various animal species (Muller et al. 1995, Morrill et al. 1997), and, although both vaccine candidates showed promising results, MP12 was reported to induce fetal malformations during the first trimester (Hunter et al. 2002); however, a recent study reported the absence of fetal malformation in pregnant ewes inoculated with the virus (Morrill et al. 2013).
Strategies to develop RVFV vaccines include subunit (Schmaljohn et al. 1989, Naslund et al. 2009, Mandell et al. 2010a), DNA (Spik et al. 2006, Lagerqvist et al. 2009), virus-like particles (VLPs) (Naslund et al. 2009, de Boer et al. 2010, Kortekaas et al. 2012), virus replicon particles (Kortekaas et al. 2011, Dodd et al. 2012, Oreshkova et al. 2013), virus-vectored (Wallace et al. 2006, Heise et al. 2009) modified live vaccines, developed from recombinant viruses engineered using reverse genetics (Ikegami et al. 2006, Bird et al. 2008, Billecocq et al. 2008, Habjan et al. 2008, Bird et al. 2011), live attenuated (Smithburn 1949, Caplen et al. 1985, Muller et al. 1995, Dungu et al. 2010, Pittman 2012, Morrill et al. 2013), and inactivated whole virus vaccines (Pittman et al. 2000). Although subunit vaccines for RVFV are generally considered safe, and recently some progress has been made in their development, evaluation of immunogenicity and/or efficacy in a target species, sheep, has been performed for a few candidates (Kortekaas et al. 2012, Oreshkova et al. 2013). On the other hand, production of live vaccines requires high level of biosafety, and their use is associated with potential risk to personnel and reversion to virulence in vaccinated hosts. Therefore, the general availability of a safe, efficacious vaccine with DIVA (differentiating infected from vaccinated animals) compatibility will be extremely valuable to endemic and nonendemic countries outside Africa.
In this study, we describe the expression of the RVFV structural glycoproteins Gn and Gc and assess their immunogenicity in a natural target species, sheep. The results demonstrate the induction of a strong virus neutralizing antibody response with potentially protective titers induced within 2 weeks after the primary vaccination without any noticeable adverse reactions. Taken together, the findings suggest that a baculovirus-expressed RVFV glycoprotein subunit vaccine could be a potential viable candidate for development of an effective vaccine against RVFV.
Materials and Methods
Cell cultures
The African green monkey cells, Vero E6 (American Type Culture Collection [ATCC], Manassas, VA), were maintained in Eagle's minimal essential medium (Corning Cellgro, Manassas, VA) supplemented with 10% fetal bovine serum (FBS),
Construction of recombinant bacmid
The cloning and construction of the recombinant bacmid was carried out as described previously (Faburay et al. 2013). Briefly, the coding sequences of the RVFV structural proteins Gn and Gc were retrieved from the RVFV ZH548 isolate sequences (GenBank accession no. DQ380151), and the sequences were synthesized (GENEWIZ, Inc., San Diego, CA) with molecular modifications. For Gn, the transmembrane domain and cytoplasmic tail were deleted from the coding sequence and designated as the Gn ectodomain (Gne) (Fig. 1A). To ensure translocation of both Gne and Gc into the endoplasmic reticulum (ER) and processing through the cellular glycosylation pathway, a specific 54-nucleotide signal peptide (SP) sequence was identified in the M segment of the RVFV genome, as described previously (Faburay et al. 2013) (sequences are available on request). This SP sequence was fused to the 5′ end of the Gne and Gc sequences (Fig. 1B). The signal peptide contained a cleavage site, and cleavage is catalyzed by signal peptidases located in the lumen of the ER, a site for protein translation and posttranslational modification.

Creation of amino-terminus glycoprotein ectodomain (Gne) and carboxyl-terminus glycoprotein (Gc) recombinant constructs and recombinant protein expression. (
The nucleoprotein (N) nucleotide sequence was amplified by PCR from the pET30 Ek/LIC recombinant plasmid (a gift from Dr. Friedeman Weber, University of Marburg, Germany), which contained the entire coding region of the S segment of the RVFV strain ZH548. The RVFV-specific sequences were cloned into a pFastBac/CT-TOPO vector (Invitrogen–Life Technologies) in-frame with a carboxy-terminal polyhistidine-tagged protein, resulting in the donor plasmids pRF-Gne, pRF-Gc, and pRF-N. The individual donor plasmids were transformed into MAX Efficiency DH10Bac-competent E. coli to generate recombinant bacmids. Recombinant bacmids were purified and used for transfection to express the respective proteins.
Recombinant baculovirus expression and purification of RVFV proteins
Recombinant baculovirus expression of the respective RVFV proteins was carried out as described previously (Faburay et al. 2013). Briefly, purified recombinant bacmids carrying the respective coding sequences (Gne, Gc, and N) of RVFV were transfected, using Cellfectin II Reagent (Invitrogen–Life Technologies), into Sf9 cells (Invitrogen–Life Technologies) grown in Sf-900 II SFM medium (Invitrogen–Life Technologies) supplemented with 10% FBS and 100 U/mL 100 μg/mL penicillin-streptomycin. Protein expression was carried out using P2 or higher-passage recombinant baculovirus stock (>107 plaque-forming units (pfu)/mL). Western blot analysis of the lysate was performed to confirm specific protein expression as described below. The proteins were expressed with a carboxy-terminal 6xHis-tag, and purification using Ni-NTA Superflow resin (Novagen, Rockland, MA) was performed according to the protocol described previously (Faburay et al. 2013). The purified proteins were stained with Coomassie Blue, and protein concentrations were determined using the bicinchoninic acid (BCA) assay (Thermo Scientific, Rockford, IL) at an absorbance of 562 nm, using bovine serum albumin (BSA; Sigma-Aldrich, St. Louis, MO) as the protein standard. Aliquots were stored at −80°C until used.
Expression and purification of RVFV N protein
Recombinant Escherichia coli expression of RVFV N protein was carried out using an expression construct kindly provided by Dr. Friedeman Weber (University of Marburg, Germany). The E. coli cells Bl21(DE3)pRARE harboring the expression vector pCDNA3.1 were grown on Luria–Bertani (LB) agar plates containing 100 μg/mL ampicillin and 34 μg/mL chloramphenicol. A single clone was picked and inoculated into 10 mL of LB medium for overnight growth. Ten milliliters of the overnight culture was then inoculated into 1 liter of LB medium, and expression was induced with the addition of 0.4 mM isopropyl β-
Detection of recombinant protein expression and analysis of immunoreactivity
The procedure for detection of recombinant protein expression by western blot has been described previously (Faburay et al. 2013). Expression of the recombinant proteins N and Gne was further confirmed using primary antibodies, mouse anti-N (R3-ID8), and the mouse anti-Gn monoclonal antibody 4D4 (a gift from Dr. Connie Schmaljohn, United States Army Medical Research Institute for Infectious Diseases), respectively, at a dilution of 1:2000. To probe immunoreactivity of antisera obtained from sheep vaccinated with recombinant RVFV Gne and Gc glycoprotein subunit vaccine, the blots were incubated with individual sheep sera at 1:100 dilution for 1 h at room temperature. After washing, the membrane was incubated for 1 h at room temperature with Protein G-HRP (Abcam, Cambridge, MA) diluted 1: 25,000. After the final washing steps, specific reactivity was detected using 3-amino-9-ethyl-carbazole (AEC) peroxidase substrate (Sigma-Aldrich, St. Louis, MO) or an enhanced chemiluminescent (ECL) detection system.
Preparation of the vaccine, animal immunization, and MP12 virus antiserum
The purified glycoproteins were formulated in montanide ISA25 water-in-oil adjuvant (Seppic, France) to obtain a concentration of 50 μg of each immunogen per vaccine dose according to the manufacturer's instruction. Six adult Rambouillet breed sheep (#9, #36, #163, #169, #170, #179) were each immunized subcutaneously with a primary dose of 50 μg of purified Gne and 50 μg of purified Gc. At day 21 after primary vaccination, hereafter referred to simply as postvaccination (pv), each sheep was given a booster with the same amount of vaccine (a second dose). At different time points after vaccination, the injection sites were inspected for possible adverse reaction. Blood samples were collected from the jugular vein of each sheep for the separation of sera prior to vaccination (day 0) to establish a baseline prevaccination immune response status. Thereafter, serum samples were collected from each sheep weekly on days 7, 14, 21, 28, 35, 42, and 49 pv. All sera were stored at −80°C until used. The RVFV MP12 control antiserum was obtained from a sheep at day 28 postinfection in a previous experiment conducted at the Kansas State University Biosecurity Research Institute (BRI) (Faburay et al. 2013). Animal experiments complied with institutional guidelines of South Dakota State University and were approved (Committee approval no. 12-037A) by South Dakota State University Institutional Animal Care and Use Committee (IACUC).
Immunogen-specific antibody response
Specific antibody response in serum was measured by enzyme-linked immunosorbent assay (ELISA) using nickel column–purified baculovirus-expressed RVFV Gne, Gc, and E. coli-expressed RVFV N proteins. The N protein was expressed in E. coil using an expression construct kindly provided by Friedeman Weber (University of Marburg, Germany) and was used as a negative diagnostic marker antigen to assess the DIVA compatibility of the recombinant subunit vaccine. To perform an ELISA, each well in a 96-well plate (Nunc, Maxisorp) format was coated overnight at 4°C with approximately 100 ng of each purified recombinant protein in 100 μL of Dulbecco's coating buffer (pH 7.4) (Invitrogen–Life Technologies). The ELISA assays were performed as described previously (Faburay et al. 2013). The cutoff point was determined for each sheep in each specific ELISA by the addition of two standard deviations to the corresponding mean OD value of the prevaccination serum. Mean OD values equal to or greater than the cutoff value were considered positive.
Anti-RVF virus plaque-reduction neutralization 80% titers
The stock of MP12 RVFV was diluted to 50 plaque-forming units (pfu) in 250 μL of 1×minimum essential medium (MEM) containing 4% BSA (Sigma-Aldrich, St. Louis, MO). Separately, aliquots of serum from each vaccinated sheep were diluted as follows: 1:10, 1:20, 1:40, 1:80, 1:160, 1:320, 1:640, and 1:1280 in 1×MEM containing 2% BSA and 1% penicillin-streptomycin. Diluted serum (250 μL) was mixed with an equal volume of diluted MP12 virus and incubated at 37°C for 1 h. Thereafter, each mixture of serum plus RVFV was used to infect confluent monolayers of Vero E6 cells in 12-well plates.
After 1 h adsorption at 37°C and 5% CO2, the mixture was removed, and 1.5 mL of nutrient agarose overlay (1×MEM, 4% BSA, and 0.9% SeaPlaque agarose) was added to the monolayers. After 5 days of incubation, the cells were fixed with 10% neutral buffered formalin for 3 h prior to removal of the agarose overlay. The monolayer was stained with 0.5% Crystal Violet in phosphate-buffered saline (PBS), and plaques were enumerated. The calculated plaque reduction neutralization 80% titers (PRNT80) corresponded to the reciprocal titer of the highest serum dilution, which reduced the number of plaques by 80% or more relative to the virus control. As positive neutralizing serum control, a 1:40 dilution of day-28 serum obtained from a sheep previously immunized with RVFV GnGc VLP was used. The serum showed a PRNT80 titer of 1280 (data not shown). Studies in humans and nonhuman primates show that a PRNT80 titer of ≥40 is protective against virulent RVFV challenge (Pittman et al. 2000, Papin et al. 2011); thus, in this study a PRNT80 titer of ≥40 was considered potentially protective.
Electron microscopy
Co-expression of the RVFV glycoproteins Gne and Gc in cell culture has been shown to result in the formation of VLPs (de Boer et al. 2010, Mandell et al. 2010b). Thus, to rule out the possibility of assembly of recombinant Gne and Gc upon mixing of the proteins into vaccine formulation, an examination by transmission electron microscopy (TEM) was performed. Briefly, equal amounts of purified Gne and Gc were mixed together in a single tube and incubated for 30 min at room temperature. As controls, aliquots of purified Gne and Gc were also made in separate tubes and incubated for 30 min as described above. Subsequently, the proteins were nebulized on copper Formar carbon-coated grids (Ted Pella Inc., Redding, CA), dried at room temperature for 30 min, and stained with phosphotungstic acid (PTA). Images were recorded at a calibrated magnification of 30,000× or 60,000× using an electron microscope (FEI Tecnai G2 Spirit BioTwin, Hillsboro, OR).
Statistical analysis
We used a t-test of independent samples with equal or unequal variances for statistical analysis. To determine the significance of observed differences in the OD values of prevaccination and postvaccination sera, a serum reactivity index (SRI) for each sheep, defined as the ratio of postvaccination serum OD value to the prevaccination serum OD value, was determined.
Results
Expression of RVFV N, Gn, and Gc recombinant proteins
Recombinant baculovirus expression of the target proteins was analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting. Using an anti-hisHRP monoclonal antibody, expression of recombinant N, Gne, and Gc glycoproteins of RVFV was detected. The expression of the recombinant proteins is a continuation of our previous work on the expression of RVFV structural and nonstructural proteins, which confirmed the expression of Gne and Gc in glycosylated form. A ∼31-kDa N protein, ∼52-kDa Gne protein, and a ∼60-kDa Gc protein were detected (Faburay et al. 2013). Expression of the N protein and Gne glycoprotein was confirmed using mouse anti-N and anti-Gn monoclonal antibodies, respectively (Faburay et al. 2013). Reactivity of recombinant Gc was confirmed by western blotting using antisera obtained from RVFV-infected sheep (Faburay et al. 2013). A Coomassie Blue stain of purified Gne and Gc used as vaccine immunogens and in ELISA showed significant enrichment of the recombinant proteins with a substantial amount of the purified total protein (estimated at ≥80%) consisting of the target protein (Fig. 1C). Additional western blotting analysis showed specific immunoreactivity of recombinant Gne and Gc with day-28 pv sheep sera, whereas recombinant N protein showed no specific reactivity (data not shown).
Vaccination and Immunogenicity of Gne and Gc glycoproteins
To examine vaccine-induced seroconversion and the kinetics of antibody response, sera collected from the vaccinated sheep at various time points pv (day 0, 7, 14, 21, 28, 35, 42, and 49) were tested in Gne and Gc-specific indirect ELISAs (Fig. 2A, B). The vaccine immunogens, as well as antigens used in the ELISA assays, were prepared from purified proteins enriched for the recombinant glycoproteins (Fig. 1C). Thus, it was considered that the overwhelming antibody responses detected in the ELISAs were vaccine induced and highly specific to Gne and Gc. Antibody reactivity with Gne antigen was detected in three of the sheep at day 7 pv (Fig. 2A). At day 14 pv, all sheep seroconverted, showing reactivity in the Gne and Gc-specific ELISAs, with Gne-specific antibodies showing comparatively stronger early-onset reactivity (Fig. 2A, B). A second vaccine dose at day 21 pv significantly (p<0.05) increased specific reactivity with both antigens at day 28 pv (Fig. 2A, B). SRI, a metric for vaccine-induced antibody response in vaccinated animals, showed an increase in OD values ranging from 4- to 9.6-fold to the Gne antigen (Fig. 2C) and 8- to 22.4-fold to the Gc antigen (Fig. 2D). For both antigens, peak induction of antibody response was observed at day 28 pv (Fig. 2A, B). Inspection of the injection sites revealed no adverse reaction (granuloma, skin ulceration, etc.) to vaccine administration.
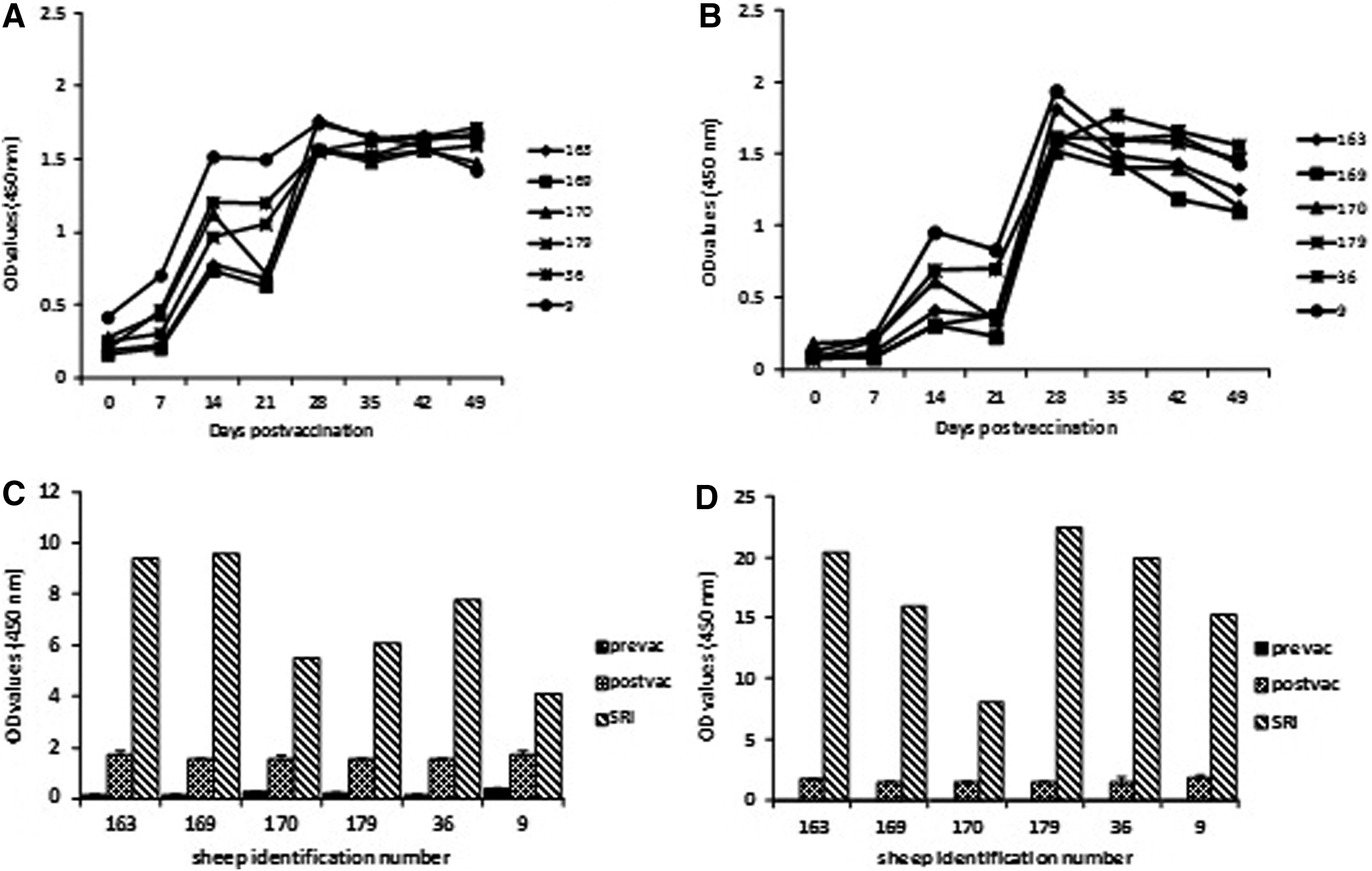
Analysis of vaccine-induced immunoglobulin G (IgG) host antibody response by antigen-specific indirect enzyme-linked immunosorbent assays (ELISAs), amino-terminus glycoprotein ectodomain (Gne)-ELISA (
DIVA compatibility of recombinant RVFV GnGc glycoprotein subunit vaccine
A DIVA concept analysis by indirect ELISA was carried out using the Gne protein, as a positive diagnostic antigen, and N protein, as a negative marker, to detect specific antibodies in vaccinated sheep. Using sera from vaccinated sheep, an increase in immunoreactivity with Gne antigen was observed from day 7 pv to day 28 pv, which later plateaued until the end of the experiment (Fig. 3A). In contrast, immunoreactivity with N antigen was barely detectable, remaining at baseline levels throughout the experiment (Fig. 3A). To confirm the specific immunoreactivity of the N protein as a marker antigen in ELISA, a day-28 MP12 postinfection antiserum (positive control) was tested alongside sera obtained from sheep vaccinated with the glycoprotein-based vaccine (Fig. 3B). Reactivity with these sera remained at baseline negative levels at all the time points, whereas the MP12 control serum showed strong reactivity as indicated by a high OD value (p<0.05).
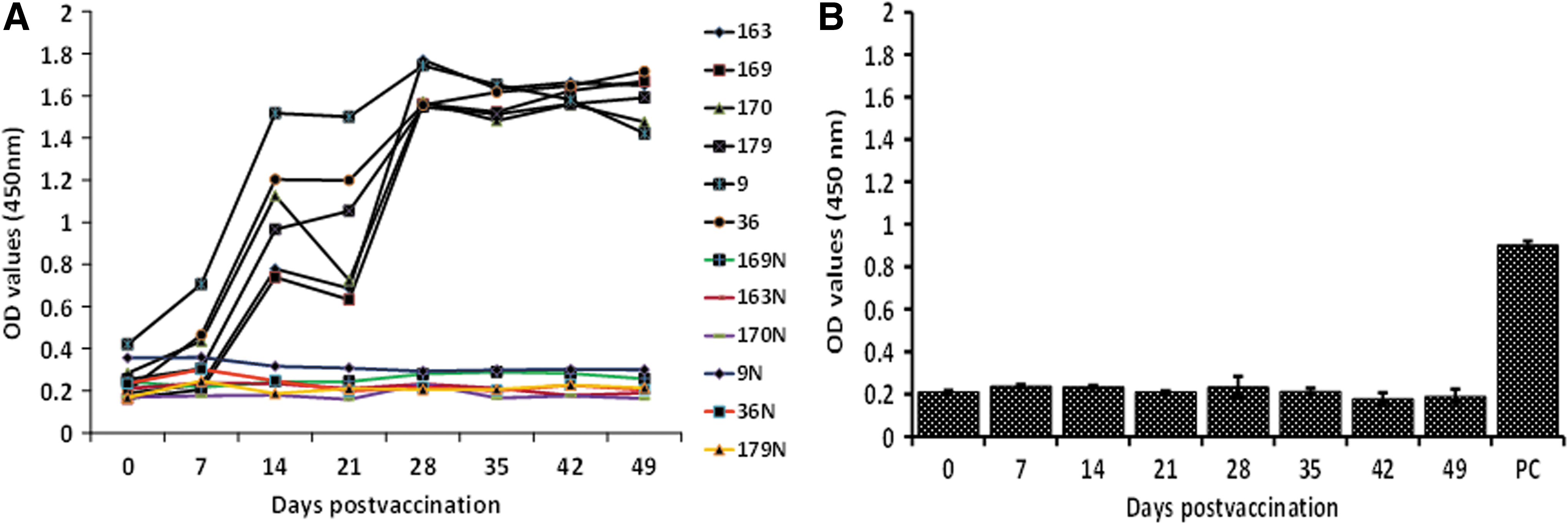
Differentiating infected from vaccinated animals (DIVA) analysis of the subunit shows indirect immunoglobulin G (IgG) enzyme-linked immunosorbent assay (ELISA) of Rift Valley fever virus (RVFV) anti-amino-terminus glycoprotein (Gn) and anti-nucleoprotein (N) antibody response in the vaccinated sheep. (
Recombinant RVFV glycoproteins elicit a neutralizing antibody response
To examine a vaccine-induced neutralizing antibody response, a PRNT assay was performed using an attenuated RVFV virus strain, MP12. A representative sample of a negative and positive PRNT80 assay using prebled and day-28 pv sera (sheep #170), respectively, is shown (Fig. 4A, B). In this study, a serum neutralizing antibody titer of 40 or greater is considered potentially protective (Pittman et al. 2000, Papin et al. 2011). Five of the six vaccinated sheep showed protective neutralizing titers at day 14 pv in response to the primary vaccination, with antibody titers ranging from 40 to 160; one sheep, #9, showed a protective neutralizing titer of 40 as early as day 7 pv (Table 1). Protective levels of virus neutralizing titers were maintained in all the sheep until day 21 pv, with three of the five sheep (#170, #179, #36) showing a titer increase. A second vaccine dose administered day 21 pv significantly boosted response in all six sheep above the 1280 titer at day 28 pv (Fig. 4C) (Table 1). Neutralizing antibody titers remained high in all sheep until day 49 pv (Fig. 4C), and all animals were positive for RVFV neutralizing antibodies when tested at day 328 pv with titers ranging from 80 to 160 (Table 1).
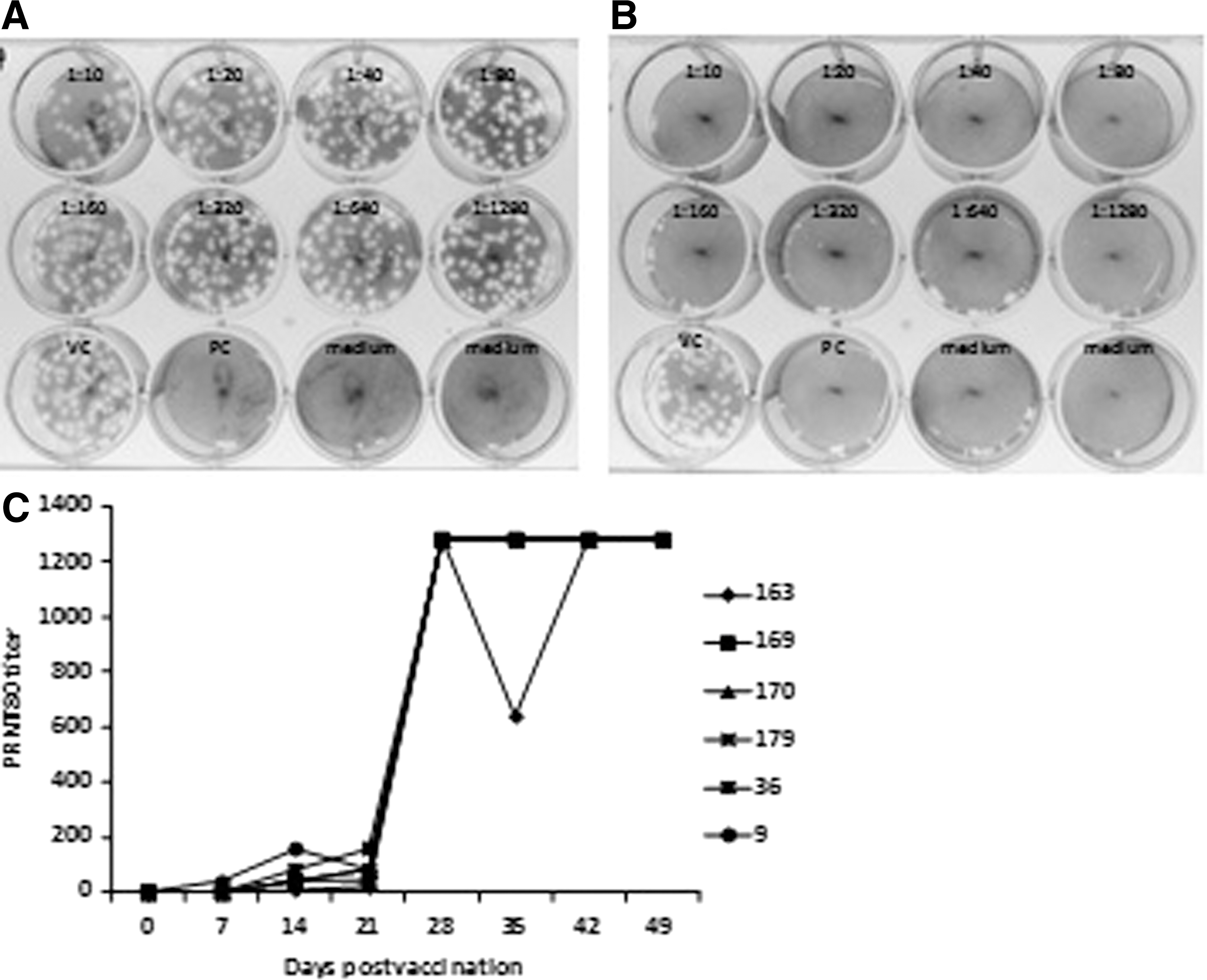
An illustration of a plaque reduction neutralization test (PRNT80) shows negative (
RVFV, Rift Valley fever virus; Gn, amino-terminus glycoprotein; GC, carboxyl-terminus glycoprotein; PRNT80, plaque reduction neutralization 80% titers; prevac, prevaccination; pv, postvaccination; nd, not determinable.
Electron microscopy
The purpose of TEM was to rule out the formation of VLPs following mixing of the recombinant glycoproteins Gne and Gc into the vaccine formulation. Images revealed by TEM showed clumps of protein aggregates that showed no resemblance to VLPs (Fig. 5).

Electromicrograph of purified recombinant Rift Valley fever virus (RVFV) proteins used in the vaccine formulation. Recombinant amino-terminus glycoprotein ectodomain (Gne), carboxyl-terminus glycoprotein (Gc), and mixed GnGc show clumps of protein aggregates that are structurally distinct from RVF VLPs, shown as control (de Boer et al. 2010). Gne and Gc are mixed in equal amounts and incubated at room temparture to allow possible assembly of proteins into VLPs. This figure illustrates the nonformation of VLPs.
Discussion
The impact of RVF outbreaks in Africa and the Arabian Peninsula, and the potential for viral spread to nonendemic areas, makes the development of safe and efficacious vaccines urgent. RVFV is a suitable candidate for a one health–focused approach to prevent both livestock and human disease through animal vaccinations (Oreshkova et al. 2013). However, there are currently no fully licensed or commercial vaccines for human or livestock use outside endemic areas, despite numerous potential vaccine candidates. Essential attributes for a RVFV vaccine include safety and high immunogenicity, DIVA compatibility, and the ability to induce a rapid onset of protective response with single vaccination, at most within 2 weeks of administration in susceptible host species.
Herein, the immunogenicity of a recombinant baculovirus-expressed RVFV Gne and Gc glycoprotein-based vaccine candidate in a natural host species, the sheep, is reported. Gn and Gc are presented as glycosylated proteins on the surface of RVF virions and have been shown to carry epitopes that elicit neutralizing antibodies, the only established correlate of protective immunity against virus infection (Besselaar, et al. 1991, Besselaar and Blackburn 1992). They are also used by the virus for attachment to target cells (Schmaljohn and Nichol 2006). Thus, the surface glycoproteins represent ideal targets for vaccine development, and we hypothesized that antibodies targeting epitopes on both structural glycoproteins in a subunit vaccine would generate a strong virus neutralizing effect. To produce vaccine immunogens, Gne and Gc expression constructs were designed to include a signal peptide at their amino terminus containing a signal peptidase cleavage site that ensures processing through translocation into the ER and the cellular glycosylation pathway and as well enhancing protein expression (Faburay et al. 2013).
Subsequently, sheep were immunized with the purified baculovirus-expressed Gne and Gc proteins with the adjuvant montanide ISA25 resulting in induction of virus neutralizing antibody response, followed by a strong response after the booster dose, in all vaccinated animals. However, due to lack of approval at the time of this study to perform challenge experiments with wild-type RVFV, it was not possible to conduct further studies to test the efficacy of the vaccine. The vaccine induced potentially protective (i.e., 1:40), virus neutralizing titers with single vaccination in five of the six animals within 2 weeks pv (Table 1). These results compared favorably with the outcome of recently reported vaccinations using vaccines based on RVFV glycoproteins, such as GnGc VLPs and Gne (Naslund et al. 2009, de Boer et al. 2010, Mandell et al. 2010a, Kortekaas et al. 2012, Oreshkova et al. 2013), as well as a Newcastle disease virus-vectored vaccine (NDFL-GnGc) (Kortekaas et al. 2010a, Kortekaas et al. 2010b) and virus replicon particles (Dodd et al. 2012, Oreshkova et al. 2013), some of which have also been reported to elicit neutralizing antibodies with single vaccination in sheep (Kortekaas et al. 2010a, Kortekaas, et al. 2012, Oreshkova, et al. 2013).
It is significant that the GnGc-based recombinant protein vaccine candidate tested in this study induced potentially protective neutralizing antibody titers with single vaccination in 80% (5/6) of sheep within 2 weeks of vaccination and 100% (6/6) sheep at three weeks pv (Table 1). The robust neutralizing antibody response elicited by the RVFV vaccine candidate could be attributed to the concurrent use of Gne and Gc proteins as vaccine immunogens. Gn is known to contain virus-neutralizing epitopes, whereas inclusion of Gc in the vaccine is suggested to provide an additional target for neutralizing antibodies (Besselaar et al. 1991, Besselaar and Blackburn 1992). Neutralizing antibody titers increased sharply in all animals following the booster, and this high anamnestic response was maintained in all animals for more than 3 weeks, i.e., up to day 49 pv. Remarkably, neutralizing antibody titers, ranging from 80 to 160, above the potentially protective threshold, were detected in all sheep at day 328 pv (Table 1). This is against the backdrop that in the United States, the withdrawal time for the adjuvant used in the current vaccine is 90 days. An early-onset vaccine-induced immunoglobulin G (IgG) antibody response to Gne occurred in half of the sheep within 7 days pv followed by seroconversion in 100% of the animals at 2 weeks pv for both Gne and Gc (Fig. 2A–D). It is noteworthy that high ELISA background was detected in some sheep. Although this is speculated to be due to a host of unknown factors, including the physiological status of individual animals, it nonetheless indicates the need for further development of the assay. Taken together, these results support the conclusion that the RVFV recombinant GnGc glycoprotein-based vaccine candidate is highly immunogenic, eliciting a strong immune response in sheep, the natural and most susceptible species to RVFV.
Differentiating infected from vaccinated animals during RVF disease outbreaks is of fundamental epidemiological importance. Therefore, DIVA compatibility of a vaccine with accompanying diagnostic tests represents an important factor for consideration when designing vaccines, especially for use in countries or regions nonendemic for RVFV. Using the RVFV glycoproteins and the nucleocapsid protein as diagnostic antigens, it was possible to distinguish vaccine-induced antibody responses (Fig. 3A) from RVFV MP12 infection in sheep (Fig. 3B). The increase in international trade in livestock coupled with the potential for RVFV outbreaks in nonendemic areas provides strong incentives for the development of DIVA vaccines. The absence of the nucleoprotein in the vaccine offers the possibility of developing a DIVA vaccine with a companion diagnostic assay using the recombinant N and Gne/Gc ELISA.
The N protein represents a suitable diagnostic antigen because it is the most abundant viral protein and is highly immunogenic, inducing antibodies within the first days after infection (Swanepoel et al. 1986, Paweska et al. 2008, Faburay et al. 2013). Furthermore, the recombinant GnGc glycoprotein subunit vaccine candidate elicited strong neutralizing and IgG antibody responses in the natural host, which can be easily detected by ELISA assays. Analysis of the structural morphology of the vaccine immunogens by electron microscopy confirmed that the proteins upon reconstitution formed into clumps or aggregates that were very much distinct from VLPs (Fig. 5). To date, RVFV VLP assembly has been reported to occur only by simultaneous production of Gn and Gc in both mammalian (Mandell et al. 2010a,b) and insect cells (Liu et al. 2008, de Boer et al. 2010) and in all cases involved the co-expression of nonhistidine-tagged proteins. These results are anticipated, for unlike with naked viruses, where VLPs are reassembled proteins, formation of an enveloped virus such as RVFV requires live cells.
RVFV represents a threat to human and animal health, and there is no commercially available vaccine or effective antiviral therapeutic agent for human use. Efforts to produce live attenuated vaccines (Bouloy and Flick 2009) are tempered by safety concerns; however, such safety concerns are comparatively lower for recombinant protein–based vaccines. However, unlike live-attenuated vaccines, recombinant protein–based vaccines generally require boosters to elicit long-term protective immune response. Therefore, future research will be directed at further improving the duration and onset of protective neutralizing antibody responses. Additionally, the fact that RVFV has low genetic diversity and consists of a single serotype suggests that the recombinant Gne and Gc glycoprotein vaccine would likely confer protection against all strains of the virus. Further evaluation of the efficacy of the vaccine in livestock and nonhuman primates is the next step toward developing a safe and efficacious vaccine for livestock and human use.
Footnotes
Acknowledgments
The authors wish to thank Dr. Connie W. Schmaljohn of the United States Army Medical Research Institute for Infectious Diseases (USAMRIID) for providing the monoclonal antibodies, and Dr. Friedman Weber, University of Marburg, for providing the RVFV N protein construct. We also thank Lloyd Willard and Joe Anderson of Kansas State University for their support in electron microscopy and Robert E. Kahn for the helpful comments. This work was supported by grants from the Department of Homeland Security Center of Excellence for Emerging and Zoonotic Animal Diseases (CEEZAD) (grant no. 2010-ST061-AG0001) and the Kansas Bioscience Authority (KBA).
Author Disclosure Statement
No competing financial interests exist.