
Abstract
Bluetongue virus (BTV) and epizootic hemorrhagic disease virus (EHDV) are arthropod-transmitted viruses in the genus Orbivirus of the family Reoviridae. These viruses infect a variety of domestic and wild ruminant hosts, although the susceptibility to clinical disease associated with BTV or EHDV infection varies greatly among host species, as well as between individuals of the same species. Since their initial detection in North America during the 1950s, these viruses have circulated in endemic and epidemic patterns, with occasional incursions to more northern latitudes. In recent years, changes in the pattern of BTV and EHDV infection and disease have forced the scientific community to revisit some fundamental areas related to the epidemiology of these diseases, specifically in relation to virus–vector–host interactions and environmental factors that have potentially enabled the observed changes. The aim of this review is to identify research and surveillance gaps that obscure our understanding of BT and EHD in North America.
Introduction
B
In this review, we examine the current status and expanding distributions of these viruses in North America, discuss the role of insect vectors, and examine patterns of disease. We identify gaps in our epidemiological knowledge, and identify strategies for addressing these gaps. This is not meant to be a comprehensive review of the Orbivirus literature, and so the reader is referred to the numerous relevant reviews that contain significant sections on the epidemiology and pathobiology of BT and EHD in North America (Hoff and Hoff 1976, Hoff and Trainer 1978, Thomas 1981, Walton et al. 1984, Gibbs and Greiner 1989, Gibbs et al. 1992, Nettles et al. 1992a, Tabachnick 1996, Howerth et al. 2001, Stallknecht and Howerth 2004, Tabachnick 2004, Walton 2004, Wilson et al. 2009, Savini et al. 2011).
Current Status of Orbiviruses in North America: Summary of the Old and the New
Viruses: Virus serotypes and their North American distribution
Bluetongue virus
Worldwide, there currently are 24 recognized serotypes of BTV (Mertens et al. 2005, Maan et al. 2007), and proposed 25th and 26th serotypes were recently detected (Hofmann et al. 2008, Maan et al. 2011). In the United States, BTV was initially isolated from sheep during a 1952 outbreak (Hardy and Price 1952), and over the next 30 years, BTV-2, -10, -11, -13, and -17 were isolated from US livestock and wildlife, representing the historically endemic BTV serotypes (Barber 1979, Gibbs et al. 1983). However, the number of BTV serotypes documented in the United States has swelled since 1999 after the detection of 10 historically nonendemic serotypes (1, 3, 5, 6, 9, 12, 14, 19, 22, and 24) in the southeastern United States (Gibbs et al. 2008, Ostlund 2010). The current known distribution of BTV in the United States is shown in Figure 1.
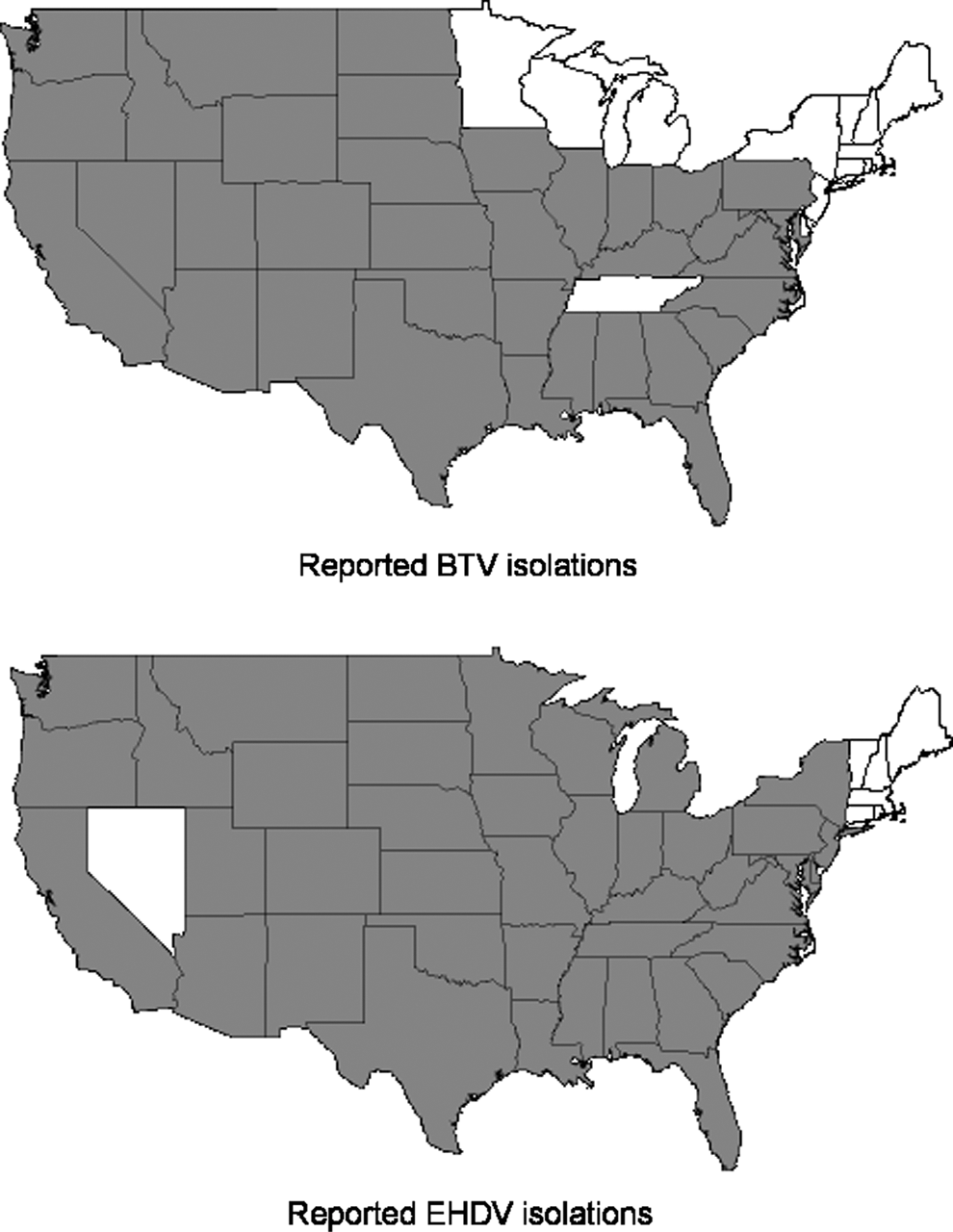
Distribution of bluetongue virus (BTV) and epizootic hemorrhagic disease virus (EHDV) isolated from ruminants by the National Veterinary Services Laboratories (USDA-APHIS-VS) and the Southeastern Cooperative Wildlife Disease Study (University of Georgia) from ruminants between 1974 and 2013. Isolations of BTV and EHDV from other laboratories are not represented here.
Since its initial detection, BTV has persisted in a stable epidemiological pattern characterized by high levels of transmission in the southern and southwestern states of the United States and reduced transmission in northern and eastern states (Metcalf et al. 1981). Canada remains free of BTV with the exception of occasional detections in southern British Columbia (Pare et al. 2012). In Montana, a low prevalence of BTV antibodies (<2.0%) has been reported in cattle (Van Donkersgoed et al. 2001, 2006). Similar results have been reported in cattle in northern Indiana and Illinois (Boyer et al. 2008) and in North Dakota and South Dakota (Green et al. 2005); in these states, antibody prevalence decreased with increasing latitude. Occasional outbreaks have occurred historically along the northern edge of the BTV distribution in the United States as early as 1966 near Billings, MT (Jones and Leudke 1969). Documented BT outbreaks have resulted in deaths of domestic sheep and numerous wild ruminant species in northern Wyoming and Montana (Thorne et al. 1988, Miller et al. 2010). These northern outbreaks are worrisome because they indicate the potential for the virus to expand its range and cause morbidity and mortality in immunologically naïve hosts, similar to recent outbreaks observed in northern Europe.
BT expansion in Europe has been occurring since the early 2000s. Expansion into the southern regions bordering the Mediterranean has been associated with range expansion of Culicoides imicola from Africa, possibly due to climate change (Purse et al. 2005). Coupled with this has been the recent expansion of an African serotype, BTV-8, into northern Europe during 2006–2008. This outbreak occurred due to the introduction of virus into indigenous vector species that had the competence to transmit BTV (Carpenter et al. 2006, 2008). The mechanism by which the virus was introduced into northern Europe is still unknown; however, range expansion of infected midges by airborne dispersal likely occurred and was responsible for introducing the virus to the United Kingdom (Burgin et al. 2013). The virus was able to overwinter successfully in northern Europe, possibly due to a combination of moderate winter temperatures and animal housing systems that facilitated vector survival throughout the winter, although other mechanisms are possible (Wilson et al. 2009). High animal morbidity was observed in cattle, possibly due to the virus strain or immune/susceptible status of the animals. Although BTV has been present in North America for over 60 years, the situation in Europe is a reminder of the potential for strains of BTV to expand beyond traditional boundaries, possibly infect novel vectors, and cause increased morbidity when entering susceptible animal populations.
Epizootic hemorrhagic disease virus
There currently are seven proposed serotypes of EHDV (1, 2, and 4–8) worldwide (Anthony et al. 2009). Although the global distribution of individual EHDV serotypes is not well understood, the viruses are generally considered to exist in temperate and tropical climates that support vector populations; thus, global EHDV distribution likely mirrors that of BTV (Savini et al. 2011). The known distribution of EHDV in the United Sates is shown in Figure 1, but EHDV has also been detected periodically in southern British Columbia and Alberta, Canada (Chalmers et al. 1964, Dulac et al. 1989, 1992). In North America, EHDV-1 and -2 have caused cyclical epidemics among ruminant populations for over 60 years. However, a nonendemic serotype, EHDV-6, was isolated from dead white-tailed deer (WTD; Odocoileus virginianus) in 2006 (Allison et al. 2010), and this virus has been isolated each subsequent year over a wide geographic area in the eastern and midwestern United States (Allison et al. 2012, DJ Johnson, unpublished). This virus represents a novel EHDV reassortant between serotype 2 (endemic) and serotype 6 (exotic). Phylogenetic analysis of EHDV-6 strains from the United States, Australia, South Africa, Guadeloupe, and Bahrain suggest that the US strain is most closely related to the Australian prototype strain (Allison et al. 2012). The route of introduction into the United States and the Culicoides species responsible for transmission are not known.
In addition to the recent detection of a novel EHDV serotype, there has also been an apparent northern expansion of EHDV in the eastern United States over the last decade. While EHDV has been sporadically detected in the upper midwestern and northeastern United States (Nettles and Stallknecht 1992) and southwestern Canada (Dulac et al. 1992, Pybus et al. 2014), these incursions have been infrequent. Given the susceptibility of WTD in northern areas to EHDV, incursions often result in high mortality, facilitating the recognition of virus circulation. For instance, the increasing frequency of recent epidemics in Michigan, Pennsylvania, New Jersey, and New York offer a good example of this apparent expansion (see the section Patterns of BT and EHD in North America, below) (Stallknecht et al. 2015). The mechanisms underlying these more frequent outbreaks have not been investigated, but understanding whether these represent repeated incursions or overwintering virus is important to ascertain. In addition, outside of North America, EHDV incursions have also occurred in cattle in many North African and Middle Eastern countries (Temizel et al. 2009, Yadin et al. 2008, Savini et al. 2011). These virus detections, along with the occurrence of disease in cattle and associated production loss (Kedmi et al. 2010), highlight the potential for EHDV introduction into susceptible cattle populations in Europe via infected vectors (European Food Safety Authority 2009).
Hosts: Ruminant hosts in North America
Despite the broad global distribution of BTV and EHDV, North America is the only continent where these viruses are known to consistently cause disease among wild ruminants (Howerth et al. 2001). This makes the North American situation unique, and numerous factors have made wild ruminants, namely WTD, excellent sentinel animals for Orbivirus activity. Indeed, EHDV and BTV serotypes previously exotic to the United States have been isolated from WTD, and this species is known to be susceptible to additional exotic EHDV and BTV serotypes (Johnson et al. 2006, Allison et al. 2010, Ruder et al. 2012a, Drolet et al. 2013). The clinical and pathological findings of BT and EHD are indistinguishable in WTD; thus, the term hemorrhagic disease (HD) is often used in this species. The utility of WTD as sentinels for Orbivirus activity in North America is based on the following: (1) WTD are extremely susceptible to EHDV/BTV infection and can develop severe disease, (2) the abundance and wide geographic distribution of WTD in North America, (3) passive surveillance of WTD through routine diagnostics by the Southeastern Cooperative Wildlife Disease Study (SCWDS; University of Georgia) and the National Veterinary Services Laboratories (NVSL; US Department of Agriculture), and (4) over 30 years of data from a long-term HD mortality questionnaire-based survey conducted by SCWDS. Collectively, decades of research and monitoring have revealed temporal and spatial patterns of infection and disease among WTD (see the section, Patterns of BT and EHD in North America) (Couvillion et al. 1981, Stallknecht et al. 2002).
Bluetongue virus
Domestic cattle and sheep are the primary domestic ruminant hosts of BTV in North America, although cattle (∼90,000,000) greatly outnumber sheep (5,400,000) (USDA National Agricultural Statistics Service 2014). Whereas wild ruminant species vary greatly across all continents where BTV is present, these two domestic ruminant species are a common thread, and numerous serological surveys and virus isolation data indicate BTV commonly infects these species in the United States (Pearson et al. 1992, Ostlund et al. 2004). In North America, reports of periodic BT outbreaks among domestic sheep and cattle have occurred since the 1950s (Hardy and Price 1952, McGowan 1953, Baldwin et al. 1991, Miller et al. 2010). However, these are typically localized epidemics that do not come close to approaching the magnitude and severity of the recent BT epidemics in Europe, or even EHD outbreaks in North American ruminants. Domestic goats and alpacas (Vicugna pacos) are also hosts for BTV (Ortega et al. 2010, DJ Johnson, unpublished), but their epidemiological significance in North America is not known.
Among North American wild ruminants, BT has been reported in WTD (Stair et al 1968), mule deer (Odocoileus hemionus) (Nettles and Stallknecht 1992), pronghorn (Antilocapra americana) (Thorne et al. 1988), bighorn sheep (Ovis canadensis) (Robinson et al. 1967), and American bison (Bison bison) (DJ Johnson, unpublished). Natural BT outbreaks among WTD, pronghorn, and bighorn sheep have been severe, resulting in high morbidity and mortality. Clinical disease associated with BTV infection has also been reported for several exotic ungulates in North American zoological collections, as reviewed by Howerth et al. (2001).
Epizootic hemorrhagic disease virus
Cattle are likely the primary domestic ruminant host of EHDV in North America, on the basis of serological surveys and sentinel animal studies (Odiawa et al. 1985, Shapiro et al. 1991, Aradaib et al. 2005, Boyer et al. 2008), as well as outbreak investigations (Foster et al. 1980, Metcalf et al. 1992, House et al. 1998, Shulaw and Zhang 2008) and the isolation of EHDV from diagnostic samples (DE Stallknecht, unpublished, DJ Johnson, unpublished). The primary outcome of infection in North American cattle appears to be subclinical infection or a mild-to-moderate transient febrile disease (Metcalf et al. 1992, House et al. 1998). However, there has been an apparent increase in the recognition of clinical disease in cattle associated with EHDV infection in numerous parts of the world, including the United States, for instance: Reunion Island in 2003 and 2009 (Bréard et al. 2004, Sailleau et al. 2012), Israel in 2006 (Yadin et al. 2008), Turkey in 2007 (Temizel et al. 2009), Morocco in 2004 and 2006 (European Food Safety Authority 2009), Algeria in 2006 (European Food Safety Authority 2009), Tunisia in 2006 (Savini et al. 2011), Jordan in 2006 (Savini et al. 2011), and the United States in 2007 and 2012–2013 (Shulaw and Zhang 2008, Ostlund 2008, Dudley 2013, Rodman and Johnson 2012).
Most recently in the United States, during the summer and fall of both 2012 and 2013, there were EHD outbreaks involving numerous cattle herds throughout the Midwest (Dudley 2013, Rodman and Johnson 2012). Unlike EHD epidemics in susceptible WTD populations, reported mortality rates in cattle are typically low, and commonly reported clinical signs include fever, inappetence, lethargy, coronitis with lameness, oral erosions/ulcers, salivation, and reduced milk yield (Yadin et al. 2008, Temizel et al. 2009, Dudley 2013). Although not appreciated historically, decreased production and significant economic loss were well documented during a recent EHD outbreak in dairy cattle (Kedmi et al. 2010). In addition to these reports in cattle, clinical EHDV infection has also been reported in other domesticated or captive-wild species in recent years, including yaks (Bos grunniens) (Van Campen et al. 2013), captive American bison (Dudley 2013), alpaca (Vicugna pacos) (Stallknecht, pers. commun.), and captive pygmy brocket deer (Mazama nana) (Favero et al. 2013).
EHDV has long been recognized as a significant pathogen of North American wild ruminants (Shope et al. 1960). Morbidity and mortality have been documented in WTD, mule deer, pronghorn, elk (Cervus elaphus), mountain goat (Oreamnos americanus), and bighorn sheep, as reviewed by Howerth et al. (2001). Of these species, epidemics impact WTD more than any other. The initial isolation and characterization of EHDV was made in 1955 when EHDV-1 was isolated from dead WTD in New Jersey (Shope et al. 1960), and it was feared that this disease would threaten the conservation of this once rare species. Given the growth of free-ranging WTD populations over the past 60 years in the face of cyclical EHD outbreaks, there are no indications that EHD will limit long-term population growth (Stallknecht et al. 2002). However, the potential for negative impact on wild ruminant populations is understudied (Fischer et al. 1995, Beringer et al. 2000, Gaydos et al. 2004), and given the dynamic pressures on wild populations in a changing environment, the possibility should not be dismissed.
Duration of BTV and EHDV viremia
The duration and titer of BTV and EHDV viremia is an important factor to consider when evaluating the potential infectivity of viremic hosts to Culicoides vectors. The duration and titer of viremia can vary among ruminant species and among individuals of the same species. Viremia, as determined by virus isolations from BTV-infected ruminants, can be prolonged and is well characterized in cattle. The maximum duration of BTV viremia by virus isolation is 63 days postinfection (dpi) based on the analysis of over 500 naturally or experimentally infected cattle (Singer et al. 2001). Viremia in BTV-infected sheep can last up to 54 dpi, although it is typically much shorter in duration (Richards et al. 1988, Koumbati et al. 1999). The maximum duration of viremia in BTV-infected cattle and sheep that is infectious to C. sonorensis was 21 dpi in both cattle and sheep, although the next time point evaluated was 49 dpi (Bonneau et al. 2002). Whereas only three of 10 midge pools were positive by virus isolation after an incubation period, blood virus titers were low (<100.5–102.4 50% tissue culture infective dose [TCID50]/mL) at the time of feeding (21 dpi), indicating potential for ruminant-to-vector BTV transmission in animals with prolonged low-titer viremia.
Unlike domestic ruminants, there has been limited experimental characterization of BTV in wild ruminant hosts, and reported viremia (by virus isolation) has typically been transient compared with cattle. For instance, duration of viremia was ≤10 days in black-tailed deer (Work et al. 1992), ≤17 days in WTD (Hoff and Trainer 1974), ≤8 days in elk (Murray and Trainer 1970), and ≤24 days in American bison (Tessaro and Clavijo 2001). However, experimental infections are scarce in these species and virus isolation techniques and study design varied among studies.
Viremia can also be prolonged in both WTD and cattle infected with EHDV, up to 59 and 50 dpi, respectively (Gibbs and Lawman 1977, Gaydos et al. 2002b). However, duration of viremia is variable and is often more transient in both species (for review, see Savini et al. 2011). Experimental infections in other wild ruminants are rare and limited to observations of 12 and 26 days in two elk experimentally infected with EHDV (Hoff and Trainer 1973). Virus could not be recovered from experimentally infected black-tailed deer (Work et al. 1992).
Virus titer in the blood of EHDV-infected ruminants varies not only between individuals, but also within an individual over the course of infection. Blood virus titers in EHDV-infected cattle and WTD generally peak 5–10 dpi before either ceasing or decreasing to a very low titer. Although studies are limited, animals with high titer viremia are more infectious to Culicoides sonorensis than those individuals with low-titer viremia. EHDV recovery was 5% from C. sonorensis 10–16 days after feeding on WTD with a blood virus titer of 103.1–3.9 TCID50/mL, compared with 50% after feeding on WTD with a high-titer viremia (107.0–7.6 TCID50/mL) (Ruder et al. 2012b). Virus could not be recovered from 617 C. sonorensis 10 days after feeding on EHDV-infected calves with low-titer viremia (<102.3 TCID50/mL) (MG Ruder, unpublished). Additional studies are needed to better understand how virus kinetics over the course of infection in the different ruminant hosts impacts infection rates in Culicoides spp.
Transmission of BTV and EHDV
Transmission of BTV and EHDV is seasonal in North America, with infection primarily occurring in the late summer and fall (Couvillion et al. 1981). In northern areas, transmission ends once adult vectors cease activity with the onset of winter. Late summer infection also is the norm in southerly locations (Gerry and Mullens 2000, Gerry et al. 2001) where adult midges may be present in low numbers year round. Virus overwintering remains a mystery, as recently reviewed by Wilson and others (2008), particularly in northern areas where freezing temperature precludes persistence in adult Culicoides and evidence of transovarial transmission and overwintering in larval populations is lacking. Virus overwintering in southern locations may be possible in adult vectors capable of surviving cooling temperatures for extended durations, and transmission may occur at levels too low for detection, although this potential phenomenon has not been well studied (Losson et al. 2007, Mayo et al. 2014). Other possibilities include overwintering in the host or alternate vectors (for review, see Wilson et al. 2008), as well as annual reintroductions from endemic to nonendemic areas through animal movement or wind-borne dispersal of the vector. The latter deserves attention in relation to annual reintroduction of viruses into nonendemic areas.
Culicoides vectors in North America
Early literature on virus transmission can be confusing if the vector subspecies is not mentioned or if the historical context of the species name is not considered. The principal vector of BTV in North America is now considered to be C. sonorensis (Tabachnick 1996). This species was originally considered one of five subspecies of the Culicoides variipennis complex (Wirth and Jones 1957) that included C. v. variipennis, C. v. occidentalis, C. v. sonorensis, C. v. albertensis, and C. v. australis. Subsequent workers proposed the synonymy of C. v. sonorensis and C. v. australis (Atchely 1967) as well as C. v. sonorensis and C. v. albertensis (Downes 1978). Jorgensen (1969) elevated C. v. occidentalis to full species status, and this was also proposed by Downes (1978), with C. occidentalis occidentalis and C. occidentalis sonorensis as subspecies. These were renamed C. variipennis variipennis, C. v. occidentalis, and C. v. sonorensis (Wirth and Morris 1985) and were later elevated to full species status (Holbrook et al. 2000). The only other confirmed BTV vector is C. insignis in Florida and the Caribbean (Tanya et al. 1992). Culicoides debilipalpis (formerly C. lahillei) has been considered a likely vector (Mullen et al. 1985a, b, Becker et al. 2010), although the studies required to elevate this species to a confirmed vector are lacking. Additionally, numerous other Culicoides spp. have been implicated as suspected vectors in certain regions of the United States (Gibbs and Greiner 1989, Mullens and Dada 1992) and targeted field and laboratory studies are needed. Figure 2 shows the historical range of confirmed and two suspected Orbivirus vectors in the United States. In addition to Culicoides spp., BTV transmission might also be possible via other vectors, such as hard and soft ticks (Stott et al. 1985, Bouwknegt et al. 2010) and sheep keds (Luedke et al. 1965). The role these and other hematophagus arthropods play in natural transmission cycles and virus overwintering is unknown.
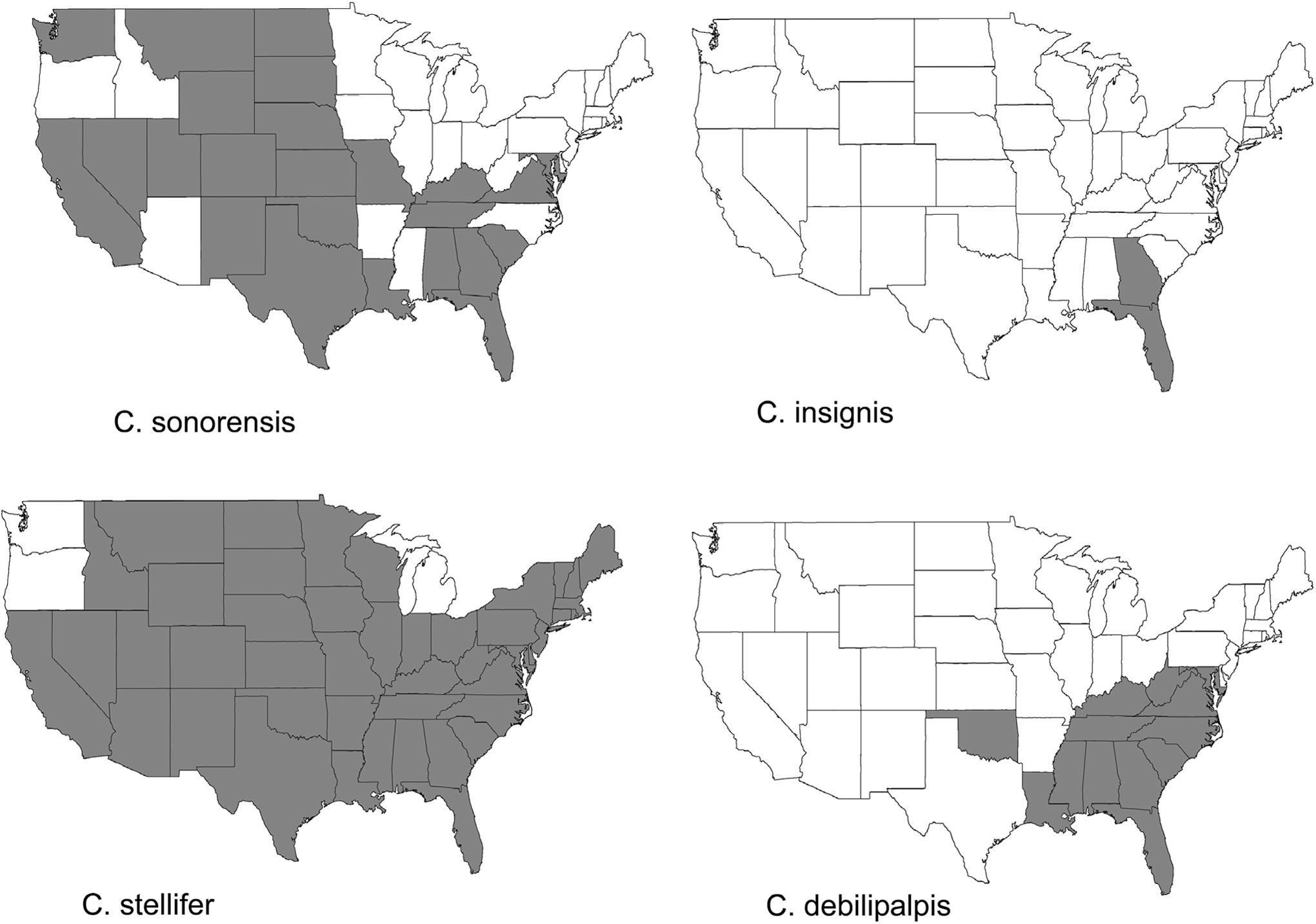
Maps showing the known distribution of confirmed (C. sonorensis, C. insignis) and suspected (C. debilipalpis, C. stellifer) vectors of bluetongue virus (BTV) and/or epizootic hemorrhagic disease virus (EHDV) in the United States. These distributions are not based on uniform, coordinated entomological surveys but rather the coalescence of smaller unrelated studies, as compiled by Blanton and Wirth (1979). More recent Culicoides surveillance is needed to determine if the known distributions of certain Culicoides spp. have changed.
C. sonorensis is the only confirmed vector of EHDV (Foster et al. 1977, Jones et al. 1977), but other species are likely to act as vectors. Expansions of EHDV into parts of the northern and eastern United States include areas that are not considered part of the historic range of C. sonorensis (Fig. 2). It is unclear if the ranges of vectors are expanding in addition to the range of the virus, if changing climate is altering the controls of vectorial capacity, or if the virus is being transmitted by other Culicoides species. Additionally, in both endemic and epidemic regions of the southeastern United States, C. sonorensis has been rare in both trap collections and animal aspirations. Based largely on midge abundance, seasonal occurrence, and host preference, C. debilipalpis, C. stellifer, C. spinosus, C. obsoletus, C. biguttatus, C. niger, and C. paraensis (Mullen et al. 1985a, b, Smith and Stallknecht, 1996, Smith et al. 1996a, b) all warrant further investigation as vectors. Of these suspected EHDV vectors, current evidence suggests C. debilipalpis and C. stellifer likely play a role, but additional work is needed to confirm these species as vectors. The role, if any, of alternate arthropod vectors in EHDV transmission has not been investigated.
Vector incrimination
Criteria for incriminating an arthropod as a vector of a disease agent include: (1) Repeated virus recovery from wild-caught specimens free of visible blood, (2) demonstration of ability to become infected by feeding on a viremic vertebrate host or an artificial substitute, (3) demonstration of the ability to transmit biologically by bite, and (4) accumulation of field evidence confirming the significant association of the infected arthropods with the appropriate vertebrate population in which disease or infection is occurring (World Health Organization 1967). Items 2 and 3 allow estimating vector competence, i.e., the genetic capability of a vector species to become infective after ingesting a viremic blood meal. This type of work has been focused on C. sonorensis, but should be expanded to other Culicoides species and other hematophagus arthropods that have a high host affinity for ruminant hosts of BTV and EHDV. There should be recognition of the potential for regional transmission cycles whereby vector species may serve slightly different functions, such as “maintenance” vectors and “epidemic” vectors.
The ability of a vector species to transmit a pathogen can be quantified using vectorial capacity. The vectorial capacity equation incorporates more information than vector competence that is listed as items 2 and 3 in the preceding paragraph. Vectorial capacity also considers field and life history components of the vector species that can influence its potential to transmit a pathogen (Lysyk and Danyk 2007). The field components are functions of the vector population's biology and abundance and include the host biting rate (bites/host per day), host preference (proportion), and vector competence (proportions). The life history component includes aspects of the vector's life history in relation to the extrinsic incubation period of the virus. These include the frequency of feeding (one/gonotrophic cycle duration), survival over the virus extrinsic incubation period, and the expectation of life of the vector. Many of the components of the life history factor are temperature dependent and show a latitudinal correlation with incidence of BTV (Mullens et al. 2004). The life history component can vary eight-fold as temperatures increase from 10°C to 30°C (Lysyk and Danyk 2007). Vector competence is the proportion of infected insects that will support virus replication and become infective. Host preference indicates the proportion of blood meals taken from a particular host species. It can range from a value of 1 (indicating strict preference for one host) or have a lesser value spread among the host species in a region. Host biting rate can show substantial variation, and, in fact, high levels of vector feeding can more than compensate for low vector competence or host preference. Unfortunately, these parameters have only been defined for C. sonorensis in a few geographic locations. Many of the risk assessments and modeling exercises for Orbivirus transmission use parameters defined for C. sonorensis, even in Europe where the species does not occur.
Alternate transmission routes
Fecal and oral shedding of EHDV have been documented in experimentally infected WTD (Gaydos et al. 2002a), but contact transmission is not suspected to play a significant role in free-ranging populations. However, artificially high animal densities, such as those encountered in farmed WTD operations, may create conditions where this route becomes important, if it occurs at all. Oral and fecal shedding of EHDV has not been investigated in cattle. Contact transmission of BTV has been suspected in WTD (Thomas and Trainer 1970) and cattle (Menzies et al. 2008). Transplacental transmission of BTV has been demonstrated in Europe in cattle (Backx et al. 2009) and sheep (Worwa et al. 2009), but the epidemiological importance of these alternate routes has not been well investigated.
Patterns of BT and EHD in North America
Understanding the incidence of disease in a zone
EHD and BT occur seasonally from mid-summer to late autumn. However, there is latitudinal and physiographic variation in the duration of transmission season that remains poorly defined (Stallknecht and Howerth 2004). Our present understanding of the delineation of epidemiological zones has been aided by decades of research and monitoring of HD in wild ruminants (Stallknecht et al. 2002). Within the distribution of wild ruminant HD mortality shown in Figure 3 are endemic and epidemic disease patterns. In endemic regions, such as the coastal plain in the southeastern United States where midge activity can persist much of the year and viruses circulate nearly annually, many infections are subclinical or result in mild disease (Nettles et al. 1992, Stallknecht et al. 2002). Moving inland in the Southeast and in portions of the Midwest, epidemics occur less frequently and high levels of mortality can be observed (Stallknecht et al. 2002). Epidemics are even more infrequent (historically) in more northern latitudes and in the western states and often result in high mortality (Nettles and Stallknecht 1992, Stallknecht et al. 2002). In addition to these endemic and epidemic cycles, a pattern of endemic stability exists in WTD in certain parts of the United States where WTD are thought to co-exist with orbiviruses and their vectors (Stallknecht et al. 1996, Stallknecht et al. 2002, Flacke et al. 2004). In these areas, prevalence of antibodies to EHDV and BTV among free-ranging WTD can approach 100% but reports of disease are extremely rare (Stallknecht et al. 1996). This endemic stability may be the result of acquired immunity via frequent exposure to the viruses, high subsequent passive immunity in fawns, and unknown mechanisms of innate immunity via co-evolution of the host and pathogen (Stallknecht et al. 2002). These mechanisms of immunity are potentially not present when captive WTD from nonendemic regions are imported to endemic regions, possibly explaining the high rates of morbidity and mortality encountered among some captive herds (Roughton 1975). Evidence to support some of the mechanisms mentioned above have been demonstrated during previous controlled experiments (Shope et al. 1960, Quist et al. 1997, Gaydos et al. 2002b, c, d). However, a better understanding of the seasonality, abundance, and competence of regional vector populations, and their interaction with the environment, is necessary to better understand these apparent patterns of infection and disease observed on the landscape.
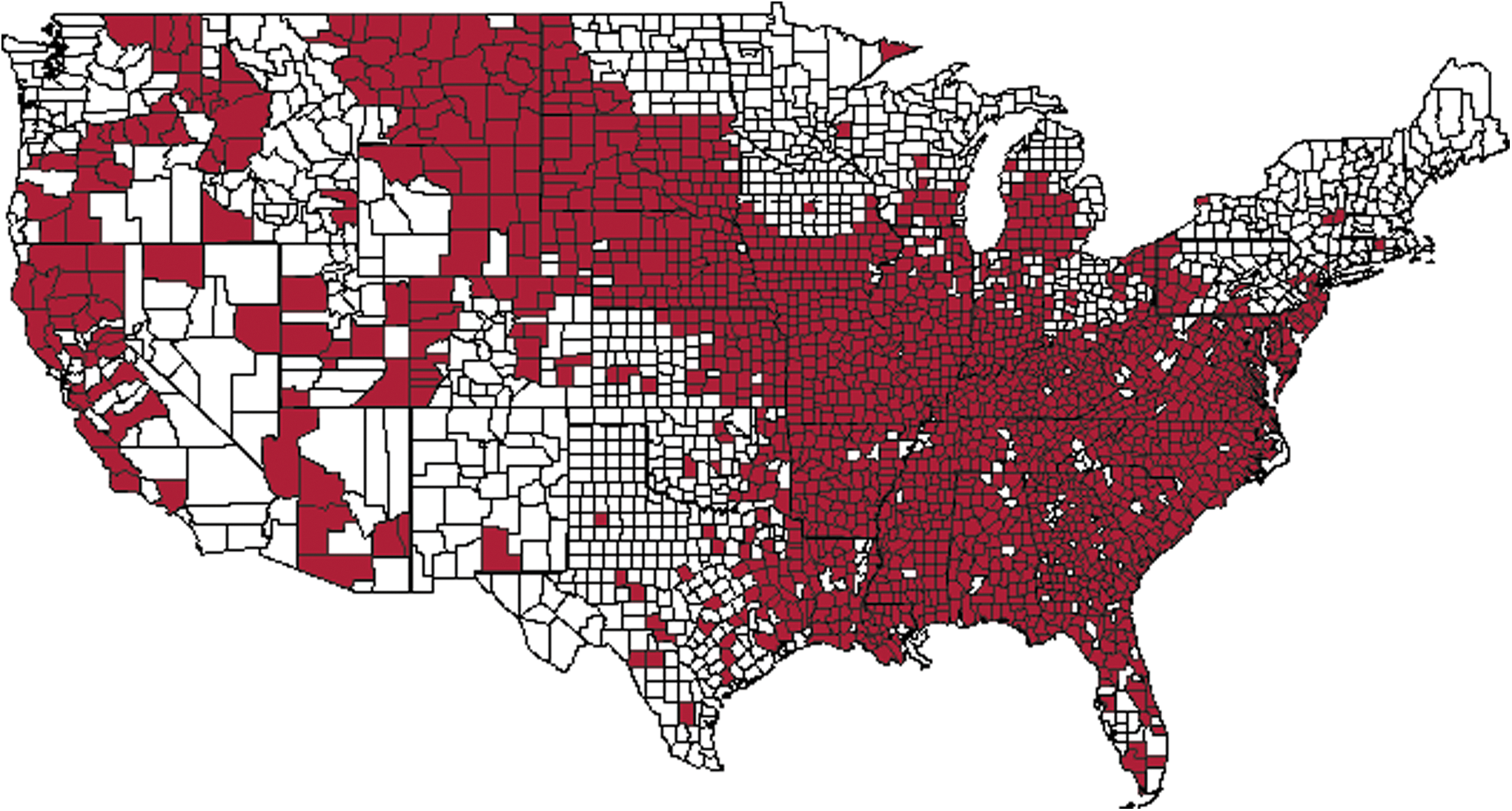
United States distribution of hemorrhagic disease (epizootic hemorrhagic disease [EHD] and/or bluetongue [BT]) morbidity and/or mortality in free-ranging wild ruminants reported to the Southeastern Cooperative Wildlife Disease Study from 1980 to 2012. Results are based on a long-term annual survey of all state wildlife management agencies in the United States to report HD occurrence on a county basis. The four case criteria for HD reporting include: (1) Sudden and unexplained deer mortality during the late summer and early autumn, (2) necropsy-based diagnosis of HD by a trained professional based on gross lesions, (3) isolation or molecular-based detection of EHDV or BTV from on affected animal, and (4) detection of deer with sloughing hooves (two or more feet), oral ulcers, or scars on rumen mucosa. This figure only represents free-ranging wild ruminants. Morbidity and mortality in domestic livestock and captive cervid herds are not represented. Furthermore, this data was compiled by the Southeastern Cooperative Wildlife Disease Study (University of Georgia) and does not include reports from other institutions. Color images available online at
The distribution of HD mortality among wild ruminants has been relatively stable (Fig. 3), and much of the upper midwestern and northeastern states have been largely free of EHDV, aside from infrequent incursions (Nettles et al. 1992b, Stallknecht et al. 2002). Historically, HD outbreaks in wild ruminants have occurred in a 2- to 3-year cycle in endemic regions, and an 8- to 10-year cycle in epidemic regions (Couvillion et al. 1981, Nettles et al. 1992b). However, over the past decade, there are indications these patterns are changing and reported outbreaks in northern areas have become more frequent. For example, confirmed EHD outbreaks have occurred in New Jersey in 1955, 1975, 1999, 2007, and 2010–2012; Michigan in 1955, 1974, 2006, 2008–2013; and in New York in 2007, 2010, and 2012. These observations are striking, yet the ecological drivers are yet to be investigated. Two of the most widespread and intense epidemics observed since the monitoring of this disease began occurred in 2007 and 2012 (Figs. 4 and 5). Both of these epidemics coincided with drought and high temperature and resulted in not only widespread mortality among WTD populations, but also disease in cattle. During the interepidemic period from 2008 to 2011, EHDV was isolated during most years in multiple northern states: Michigan (4 years), Indiana (4 years), Pennsylvania (3 years), and New Jersey (4 years) (DE Stallknecht, unpublished). More recently, EHDV has been isolated from ruminants in South Dakota each year 2011–2013, as well as in Nebraska and Iowa during 2012 and 2013 (DJ Johnson, unpublished). This frequency of isolation of EHDV from northern latitudes in the United States highlights our lack of understanding of the epidemiology of EHD, especially in regard to surveillance, overwintering mechanisms, and competent vector populations.
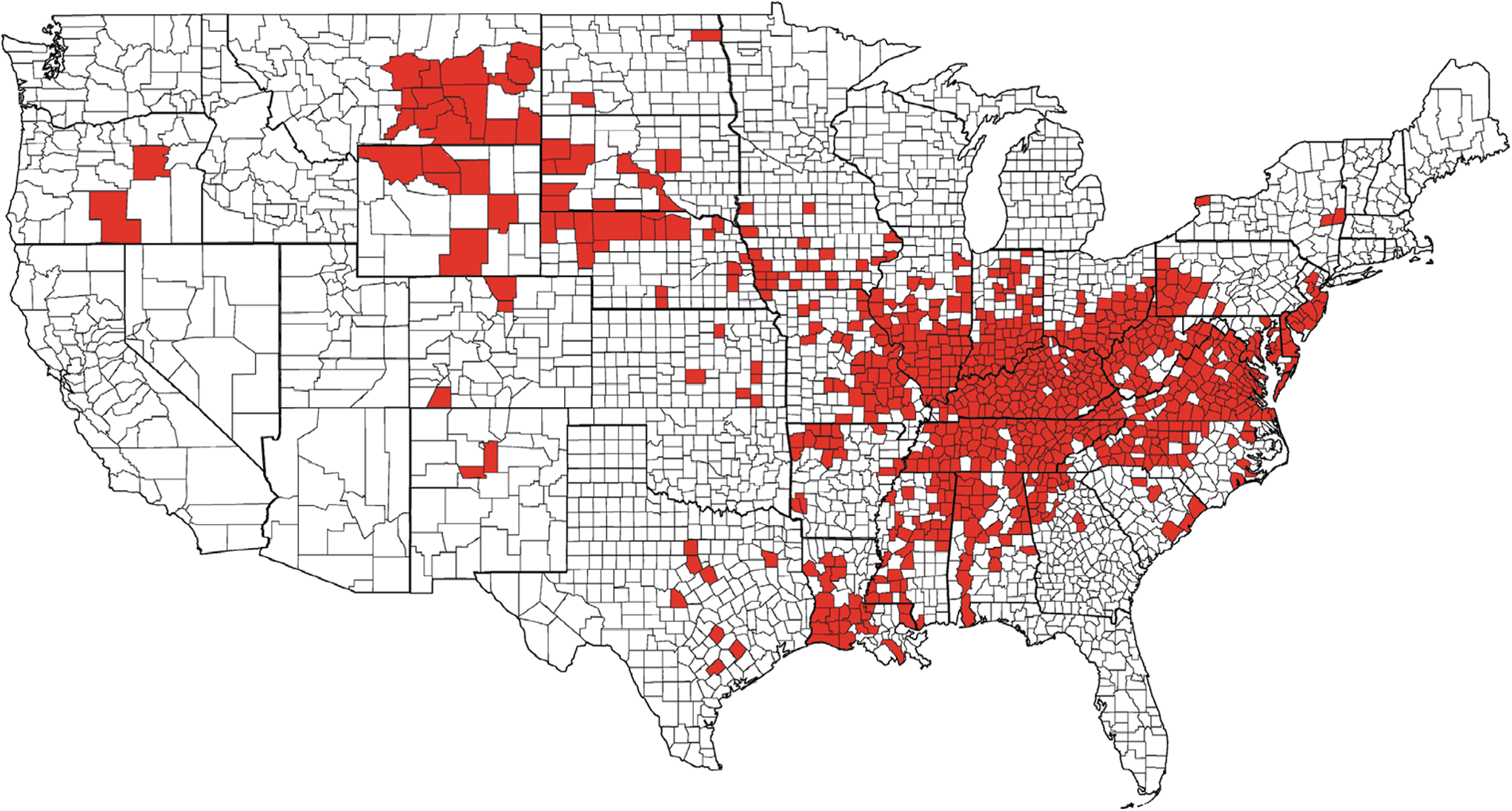
United States distribution of hemorrhagic disease (epizootic hemorrhagic disease [EHD] and/or bluetongue [BT]) morbidity and/or mortality in free-ranging wild ruminants reported to the Southeastern Cooperative Wildlife Disease Study during the 2007 epidemic. The outbreak was geographically expansive and appeared to be two distinct epidemics. The western focus was dominated by BTV-17, whereas EHDV-2 was the primary isolate in the eastern outbreak. During this epidemic, 11 midwestern and mid-Atlantic states had counties with more than 100 dead white-tailed deer (WTD) reported, with many counties reporting estimates of >1000 mortalities. This figure only represents free-ranging wild ruminants. Morbidity and mortality in domestic livestock and captive cervid herds are not represented. Furthermore, this data was compiled by the Southeastern Cooperative Wildlife Disease Study (University of Georgia) and does not include reports from other institutions. See Figure 3 legend for description of how data were collected. Color images available online at
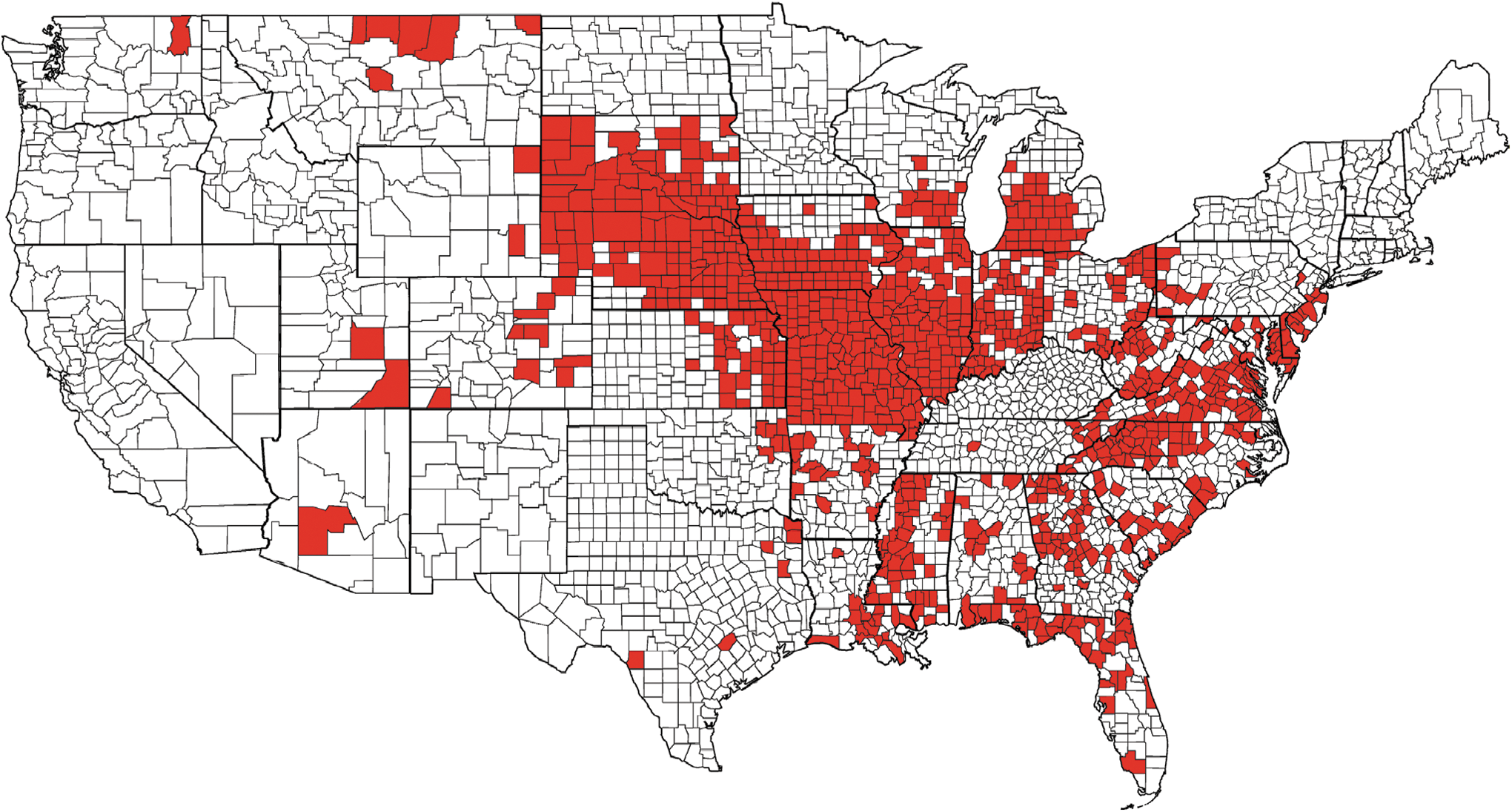
United States distribution of hemorrhagic disease (epizootic hemorrhagic disease [EHD] and/or bluetongue [BT]) morbidity and/or mortality in free-ranging wild ruminants reported to the Southeastern Cooperative Wildlife Disease Study during the 2012 epidemic. The geographically expansive outbreak in the Midwest was dominated by EHDV-2 and EHDV-6. This figure only represents free-ranging wild ruminants. Morbidity and mortality in domestic livestock and captive cervid herds are not represented. Furthermore, this data was compiled by the Southeastern Cooperative Wildlife Disease Study (University of Georgia) and does not include reports from other institutions. See Figure 3 legend for description of how data were collected. Color images available online at
Compared to wild ruminants, reports of EHD or BT in livestock in the United States are less frequent, and large outbreaks typically coincide with large-scale epidemics in sympatric wild ruminants (e.g., 2007 and 2012). Thus, when attempting to understand epidemiological patterns in livestock, there is more reliance on traditional serological surveys, sentinel animal studies, import/export diagnostic sample submissions, and individual animal or herd level clinical case submissions. Although domestic cattle are not as susceptible as WTD to EHD and BT morbidity and mortality, serological surveys in North America have generally supported the presence of endemic and epidemic disease zones observed in wild ruminants. For instance, serological surveys of cattle for antibodies against EHDV and BTV (Metcalf et al. 1981, Odiawa et al. 1985, Shapiro et al. 1991, Ostlund et al. 2004, Green et al. 2005, Boyer et al. 2008) suggest that seroprevalence rates are latitudinally/regionally stratified. For instance, during 1977–1978 and 1983–1985, regional surveys of cattle for BTV antibodies were performed yielding seroprevalence rates of <1% in 18 northern and northeastern states, 18–53% in nine central and southwestern states, 6–39% among 13 southeastern states, and 80% in Puerto Rico (Metcalf et al. 1981, Pearson et al. 1992). Subsequent follow-up surveys of the northern and northeastern states performed in the 1990s and early 2000s helped to further define the distribution of BTV in the northern United States (Ostlund et al. 2004). Collectively from 1977 to 2002, market cattle surveys in New England, Michigan, Minnesota, New York, and Wisconsin revealed a seroprevalence of <2.0%.
Although it is clear that there is a gradation of BTV/EHDV activity and that endemic, epidemic, and incursive disease zones exist in North America, these regional zones are yet to be systematically investigated and defined. The BTV-free zone in parts of the northern and northeastern United States has been well investigated; however, given the changes observed globally over the past decade, we should not assume this BTV-free status will remain unchanged. Much of our current understanding of the North American zones is based on disease reports, surveys, and the distribution of C. sonorensis. These zones can be more clearly defined not only through additional serological surveys and strategic sentinel animal studies, but also investigation of additional Culicoides species. There is a need to better understand the ecology and life history of Culicoides species in different physiographic regions. For instance, the seasonal abundance, vector competence, host preference, larval habitat, and adult habitat utilization of different Culicoides species will provide insight into patterns of infection and disease. At the same time, it will be important to consider that the disease zones in the United States may be different for EHDV and BTV, reflecting potential differences in transmission that may exist between these closely related viruses.
Factors influencing BT and EHD incidence
Epidemiological studies examining the relationship between BTV and/or EHDV infection and various environmental and climatic variables in North America are limited (Sellers and Maarouf 1989, 1991, Sleeman et al. 2009, Boyer et al. 2010, Mayo et al. 2012, Xu et al. 2012, Berry et al. 2013) but have revealed some interesting associations that warrant further investigation. Recent spatial statistical modeling of HD in WTD from 1980 to 2007 revealed that wetland cover is likely a critical driver of HD morbidity (Berry et al. 2013). The factors underlying this association were not investigated, but likely relate to WTD habitat utilization and larval development sites for Culicoides species. Other studies have found significant clustering, or patchiness of infection, or outbreaks (Boyer et al. 2010, Xu et al. 2012). In a 3-year study by Boyer et al. (2010), temperature was associated with EHDV and BTV seropositivity in cattle during all 3 years of the study, although there were differing spatial distributions between viruses. Furthermore, forest patchiness was associated with EHDV seropositivity in cattle for two of three seasons, and heavy spring rainfall was hypothesized to play a role in the lack of association during the third year, which is supported by other studies (Sleeman et al. 2009). Interestingly, there was no association observed between BTV seropositivity and forest patchiness.
Vector distribution and larval habitat likely play a significant role in the occurrence of infection and disease. For instance, C. sonorensis is often found closely associated with livestock and organically loaded water sources, and populations are distributed throughout the southern and western United States yet absent in the upper Midwest east of the Missouri River and in the northeastern United States (Holbrook et al. 2000). It has been suggested that soil type and climate likely help determine the distribution of the C. variipennis complex in the United States, and these findings may help to explain the absence of BTV in the northeastern United States (Schmidtmann et al. 2000, 2006, 2011). However, BTV is occasionally detected in these regions and EHDV commonly circulates in these areas (Fig. 1), indicating the need for additional entomological surveillance. Variation in research findings and field observations, such as those mentioned above, suggest that arbitrarily lumping EHDV epidemiology with BTV epidemiology should be done cautiously, as there may be significant differences in the ecology of these diseases. Additional studies will be necessary to better understand the ecology of these diseases in a changing environment and climate.
Molecular epidemiological investigations of field strains of EHDV and BTV in North America are limited (Pritchard et al. 1995, Cheney et al. 1995, 1996, Wilson et al. 2000, Cheney et al. 2003, Mecham et al. 2003, Murphy et al, 2005, 2006, MacLachlan et al, 2007, Balasuriya et al, 2008, Allison et al, 2010, 2012, Anbalagan et al. 2014). Many of these previous studies have relied on partial gene, full gene, or multiple gene segment sequence for analysis and have revealed significant findings regarding novel viruses, outbreak dynamics, and evolution of viruses in North America. However, with the cost and speed of full-genome sequencing decreasing, great potential exists for additional phylogenetic studies. The long-term storage of virus isolates, such as those at the SCWDS and NVSL, offer an invaluable resource. Phylogenetic analysis of field isolates at various temporal or spatial scales can provide insight into a diversity of epidemiological questions.
Current sources of BT and EHD epidemiological information
At present, neither BT nor EHD are considered or treated as reportable diseases (state or federal) in the United States. Epidemiological information is not routinely collected through national surveys. What data are collected are maintained at the Center for Epidemiology and Animal Health via the National Animal Health Reporting System (NAHRS). This system is an aggregation point for reports from state veterinarians on the presence of World Organisation for Animal Health (OIE)-listed diseases in the United States. There is some uncertainty associated with reporting at the state level because reports are often given as presence or absence of the disease. For wild ruminants, a long-term virus isolation and HD morbidity/mortality dataset is maintained by SCWDS. There are also a number of veterinary diagnostic labs that operate throughout the country that capture information based on diagnostic submissions. These include the NVSL, various State veterinary diagnostic laboratories, university laboratories, and Newport Laboratories (Worthington, MN) and other private labs. A more coordinated effort by the various agencies to collect information could provide a starting point for increasing our understanding of the epidemiology of BT and EHD in the United States.
Summary of Orbivirus Epidemiology Research Gaps
Research gaps presented below were classified into two broad categories posed as questions. The first category includes gaps related to the overall pattern of BTV and EHDV transmission in North America, whereas the second category includes gaps related to the expression and incidence of clinical disease within the epidemiological zones. Within these two broad categories, we have identified a number of more specific research gaps that are distinct, yet interconnected. Following this, we propose a framework for several coordinated research and surveillance strategies that could begin to resolve these gaps.
What is the pattern of North American ecosystems supporting the arthropod transmission of BTV and EHDV in different climatic/geographic zones?
Previous studies clearly demonstrate variable patterns of infection and disease among North American ruminants. A fundamental step toward understanding these patterns is determining the distribution and diversity of EHDV and BTV serotypes, molecular subtypes, and subgroups within North America. This, along with serological and virological surveys of sentinel herds and wild ruminants, will begin to elucidate the true endemic, epidemic, and incursive zones, as well as areas of transition between epidemiological zones. Understanding how changing land-use patterns, habitat fragmentation and augmentation, and climatic variables impact the delineation of the endemic, epidemic, and incursive zones will be important considerations in a landscape dramatically influenced by human activity and climate change. A better understanding of the defining traits of these epidemiological zones will facilitate the targeted study of potential virus overwintering mechanisms, as well as aid in the identification of mechanisms that allow viruses to expand their range. These mechanisms of expansion might include natural or human-influenced movement of infected vectors or ruminants, but we must also be open to consideration of alternate host–virus or vector–virus pathways to enable virus expansion and subsequent persistence in different ecological areas. Understanding complex interactions between hosts, viruses, vectors, and the environment may enable the identification of likely points of virus introduction into an area and subsequent risk factors for persistence in a particular ecosystem. Whereas numerous directed studies, surveys, and diagnostic investigations of varying scope and intensity have been performed and have laid the foundation for our present understanding of Orbivirus epidemiology in North America, there is a need to integrate this information and to ultimately coordinate future research and surveillance efforts.
There is also a need to re-examine the current distribution of competent vectors of BTV and EHDV in the United States, especially considering the recent changes in virus distribution and disease incidence mentioned previously. However, this effort should go beyond simply documenting the distribution of C. sonorensis and C. insignis (the only confirmed Orbivirus vectors in North America), and must also explore suspect and potential vector species. Although the midge distributions presented in Figure 2 are based on sound information, these data are decades old and we should not assume these to be static. Field and laboratory studies need to be conducted to confirm the vector status of additional species, including the identification of hematophagous arthropods feeding on susceptible hosts in various regions of the United States, entomological investigations of outbreaks, and laboratory studies on vector competence. Improved sampling and vector surveillance strategies should be developed and used to identify and characterize areas of adult abundance and larval development sites. This work will advance our understanding of Culicoides ecology and will facilitate the development of novel control strategies, identification of risk factors, and inform the development of ecological niche models. Long-term studies on larval and adult population dynamics of vector species in various geographic regions will fill gaps in our understanding of the biology and ecology of vector species in relation to patterns of infection and disease on the landscape.
A critical component of an active and productive entomological surveillance and research network in North America will rely heavily upon providing training and support for arthropod identification. Regional or national Culicoides identification guides need to be created based on collections in the Florida State Arthropod Collection. New technologies, such as molecular barcode identification, should be developed to help with identification; however, the invaluable skill of classical morphological identifications among North American entomologists must be maintained.
What are the factors influencing the expression and incidence of clinical disease within these epidemiological zones for all potential hosts?
For all of these epidemiological zones, one common theme is the absence of knowledge about how these viruses are maintained between seasonal transmission periods. Obtaining a better understanding of the defining characteristics of each of the epidemiological zones mentioned above, such as seasonal vector abundance and diversity, virus serotype diversity, and antibody prevalence, is essential to determining the underlying factors that influence disease incidence within a zone. Weather patterns and climatic trends may impact disease incidence, in addition to the previously mentioned factors of virus transmission, animal movement, and host factors (e.g., breed and species variation in susceptibility and adaptive and innate immunity).
Wild ruminants can move across the landscape, but they encounter tremendous obstacles in most regions due to habitat fragmentation and variable land-use patterns. This drastically limits long-distance migrations among wildlife; however, the interstate shipment of livestock and captive WTD is routine in the United States. The impact of animal movement between different epidemiological zones in relation to virus movement and diversity, as well as disease distribution, has not been examined. The role of innate and adaptive immunity in the incidence of disease in different ruminant breeds and species should be explored, because numerous studies in WTD suggest host immunity is critical (Stallknecht et al. 1996, Gaydos et al. 2002a, b). A relatively recent change has been the large growth of the captive WTD industry in the United States. In many operations, natural selection has been removed and breeding is managed for the exploitation of antler development in most herds. The impact of this selective breeding on disease susceptibility is unknown and should be investigated, as should the potential role of dense accumulations of highly susceptible hosts on outbreak dynamics in adjacent wildlife and livestock. Furthermore, while some studies have identified an effect of local and regional weather patterns and climatic conditions on outbreaks and disease incidence, these areas of study have been left largely unexplored in North America. These studies have identified associations between virus exposure or disease incidence and weather/climate patterns; however, the mechanisms underlying these associations remain unknown.
Management of arthropod-borne infections is difficult, especially in a multihost system involving both livestock and free-ranging wildlife. Management of these diseases in wild ruminant populations is extremely difficult at best. Depending on the nature of the livestock operation, the goal of any control strategy will be to either prevent virus exposure or prevent disease, but it is important to consider that a high prevalence of EHDV or BTV are not necessarily associated with morbidity. To develop novel Culicoides control strategies or consider vaccine programs against EHD and BT, we must first understand some fundamental aspects of the epidemiology. The gaps outlined above begin to address many of those gaps that will enable the informed development of future control strategies.
Strategies toward filling research gaps
It has long been recognized in the North American Orbivirus research community that there is a need to develop a coordinated, comprehensive active Orbivirus surveillance program for livestock and wildlife (Gibbs et al. 2008). This is potentially an expensive and logistically complex endeavor that becomes more practical if an incremental approach is considered. For instance, if we embrace the present network of diagnostic, surveillance, and research efforts across the country and find ways to form linkages, we can reshape and then grow the system. This will rely heavily upon open communication and being able to recognize and take advantage of opportunities for association and collaboration in order to facilitate the layering of existing programs. Future programs that fill voids can be developed and incorporated into the network.
There is a need to coordinate disease surveillance efforts that currently consist of observations and records of clinical disease from state and federal diagnostic labs, other research labs, the annual SCWDS HD surveillance data, as well as other investigations of disease outbreaks. Similarly, there is potential to coordinate virus surveillance in the form of observations of serological distribution within states or regions. This should be augmented with regional sentinel cattle and captive WTD herds to better understand virus distribution and more clearly define epidemiological zones. Initially, the focus should be to determine the transitional front between epidemic and incursive zones, as well as the southern United States to recognize potential virus introductions. Expansion of sentinel herds into other regions can follow.
Utilizing captive ruminants at land grant schools, government (federal and state) research herds, herds maintained by correctional facilities, and other research herds, as well as captive WTD farms, meat processors, taxidermists, and the Deer Management Assistance Program are all potential avenues to explore to enhance surveillance and reporting efforts. Such a network would facilitate molecular epidemiological investigations to understand complexities of virus movement, overwintering, and evolution. Vector surveillance could be standardized with gridded, uniform trapping systems to determine the presence/absence of known and potential vector species in association with sentinel cattle and WTD herds. Live animal baited aspirations should be performed and compared to different trap data in different physiographic regions of North America. Finally, coordinated efforts need to be made to analyze and model current data and evaluate future scenarios to determine the impact of BT and EHD in North America and predict future events. These efforts could include assessments of the impact of EHD and BT on livestock and wildlife population abundance, health, welfare, and productivity, retrospective and prospective studies to evaluate the economics of the problem and the potential cost–benefit of control strategies, and the creation of predictive models of the expansion of disease incidence and outbreaks in response to weather events and climate change.
Footnotes
Acknowledgments
We acknowledge the organizing committee and the US Department of Agriculture (USDA) Agricultural Research Service (ARS), National Program for Animal Health for sponsoring and conducting an Orbivirus Research Symposium during May, 2013, in Manhattan, Kansas, at the Center for Grain and Animal Health Research (CGAHR, USDA-ARS). The authors were grateful to participate and thank all of the participants for donating their time and talent. This paper will be part of the report from that symposium. The authors also thank the staff of the Arthropod-Borne Animal Diseases Research Unit and the CGAHR for on-site support of the symposium.
Author Disclosure Statement
No competing financial interests exist.