
Abstract
Arthropod disease vectors, most notably mosquitoes, ticks, tsetse flies, and sandflies, are strongly influenced by environmental conditions and responsible for the vast majority of global vector-borne human diseases. The most widely used statistical models to predict future vector distributions model species niches and project the models forward under future climate scenarios. Although these methods address variations in vector distributions through space, their capacity to predict changing distributions through time is far less certain. Here, we review modeling methods used to validate and forecast future distributions of arthropod vectors under the effects of climate change and outline the uses or limitations of these techniques. We then suggest a validation approach specific to temporal extrapolation models that is gaining momentum in macroecological modeling and has great potential for epidemiological modeling of disease vectors. We performed systematic searches in the Web of Science, ScienceDirect, and Google Scholar to identify peer-reviewed English journal articles that model arthropod disease vector distributions under future environment scenarios. We included studies published up to and including June, 2014. We identified 29 relevant articles for our review. The majority of these studies predicted current species niches and projected the models forward under future climate scenarios without temporal validation. Historically calibrated forecast models improve predictions of changing vector distributions by tracking known shifts through recently observed time periods. With accelerating climate change, accurate predictions of shifts in disease vectors are crucial to target vector control interventions where needs are greatest.
Introduction
R
The failure of the space-for-time assumption (Blois et al. 2013) imposes strong limits on the reliability of spatial models for temporal prediction. For example, nine modeling techniques used to forecast four South African plant species' distributions in 2030 predicted between a 92% loss to a 322% gain, but all models successfully described the current spatial distributions of these species (Pearson et al. 2006). Furthermore, environmental factors that correlate positively with species' distributions spatially may show the opposite relationship temporally. For example, models of Canadian butterfly species' richness showed a positive association with human population density spatially but a negative association temporally (White and Kerr 2006). Species distribution modeling methods range from logistic regression to highly specialized machine-learning techniques (Elith and Leathwick 2009), all of which have proven useful under different circumstances. Strong tests of model accuracy are critical to forecasting how changing environmental circumstances will alter the distributions of disease vectors through time.
Accurately projecting distributions of human disease vectors, such as those transmitting malaria, Lyme disease, dengue, and trypanosomiasis, is crucial because of their human health impacts and the related economic consequences of misdirected management interventions. Yet, predictions of future vector distributions often conflict. For example, models of Anopheles mosquito distributions, the vectors of the malaria parasite, across the Caribbean did not predict northern range expansions into the Gulf coastal region under warming climates by 2080, contradicting previous studies which predicted future expansions of malaria by as early as 2050 (Fuller et al. 2012a). Conflicting results of future vector distributions mislead control programs and practices that already pose substantial financial challenges for many developing countries (Martens et al. 1999, World Health Organization 2008). Technological advances are unlikely to permit predictions of future species distributions to be tested unambiguously; the definitive, but impractical, method for testing such models is to wait to compare predictions to actual events (Rastetter 1996, Algar et al. 2009). This does not mean that forecasting future species distributions and validating these methods lacks rigor. It does, however, underscore the need for strong validation methods to increase confidence that such models are reliable (Botkin et al. 2007).
There are extensive parallels between macroecological models, which predict species distributions over broad areas (e.g., regions and continents; see Brown and Maurer 1989, Kerr et al. 2007), and epidemiological models needed to predict changing vector distributions across comparable modeled areas. Both may use correlative and mechanistic methods for projecting changing species distributions (for a comparison of these two methods in vector modeling under future climates, see Fischer et al. 2014). Correlative models can include general linear models or use machine-learning techniques, such as maxent (Phillips et al. 2006), techniques from the Biomod modeling package, such as random forest analyses (Thuiller 2003), and genetic algorithms for rule-set production (Stockwell and Peters 1999) to relate species occurrence records to environmental factors and determine the area the species actually occupies (i.e., realized niche). Mechanistic models, or stochastic models coupled with habitat suitability or population models, use lab- and field-based measures of physiological traits (e.g., behavior, temperature tolerances, energy turnover) to determine all potential areas where the species may persist (i.e., fundamental niche) (Kearney and Porter 2009). Temporal macroecological models often seek to forecast future species distributions for the purposes of conservation (Trejo et al. 2011), invasive species (Sobek-Swant et al. 2012), or agriculture (Ureta et al. 2012); their forecasting and validation methods show considerable overlap with those predicting changing disease risk. Satellite remote-sensing tools provide a common array of baseline measurements for both fields that are revolutionizing statistical modeling in epidemiology (Hay et al. 2000) and macroecology (Kerr and Ostrovsky 2003) by improving detection of spatial and temporal variation in the biophysical determinants of species distributions. Consequent syntheses of techniques from both disciplines has led to the emergence of “eco-epidemiology,” “landscape epidemiology” (Pavlovsky 1966, Kitron 1998, Peterson 2006, Reisen 2010), “medical geography” (Meade and Earickson 2000), and “disease macroecology” (Keith et al. 2012).
In attempting to predict changes in species distributions through time, predictive models commonly share a straightforward approach. Generally, these models: (1) Use current environmental conditions to predict current disease vector distributions, (2) test the accuracy of the model by evaluating its fit to current spatial observations, and (3) project the model to a future time period by substituting anticipated environmental observations into the spatial model. Spatial validation, which improves confidence in model fit in the current time period, does not imply temporal reliability. To ensure the latter, analyses must be multitemporal (Kharouba et al. 2009). A validation technique that accounts for time (or historical calibration) is needed. Using this approach, models are constructed with observations from different time periods. These models are then projected either backward or forward into a time period with independent vector data to test their capacity to predict changing distributions through time. Historical calibration is likely the minimum requirement to establish confidence that these models can be used to forecast changes in vector distributions accurately (e.g., Kharouba et al. 2009, Rapacciuolo et al. 2014). This approach provides spatiotemporal tests of model reliability, rather than spatial tests followed by risky reliance on space-for-time assumptions.
Here, we review the critical field of forecasting future disease vector distributions under changing climate. First, we discuss the different methods used to validate the spatial models of present-day vector distributions. Next, we assess how these spatial models are projected into future time periods under different climatic scenarios. We then introduce the framework we call historical calibration for validation that is specific to temporal extrapolations in macroecology and may increase accuracy of forecasting models in epidemiology. While we focus on disease vectors that directly affect human health for this review, the assessment of the forecast and validation approaches can be applied across spatial scales and broadly among many focal species and hosts, such as livestock (e.g., for advances in forecast modeling of Culicoides midges and the bluetongue virus in livestock, see Acevedo et al. 2010 and Guichard et al. 2014).
Materials and Methods
We performed systematic searches in three databases for peer-reviewed literature: Web of Science (
Results and Discussion
Validating spatial models
First, it is important to distinguish between model fit—whether a model's predictions reasonably match data available within an observed time period—and true model validation—determining whether the parameters of a model are causally linked with the biological response, which it predicts accurately. The former is the domain of approaches such as the “split data” approach. The latter truly tests model predictions, rather than model fit. However, true model validation is more challenging because it requires independent disease vector data to test models' generality or temporal transferability (i.e., data not used in model training and that are spatially or temporally independent). All too frequently, such data do not exist. While the validation methods reviewed were limited by inevitable autocorrelation issues or data availability, and occasionally were not described at all, these 29 studies generally included strong validation techniques, which are categorized and described below.
The most common test of model fit we observed in these studies is a variation of the “split-data” approach (see Table 1). This approach is a subset of cross-validation, which divides datasets into two parts—training data used to calibrate the model and testing data to evaluate its predictive accuracy. The “split-data” approach randomly splits the original data into training and testing data (e.g., 70% of occurrence data are used for training the model, and the remaining 30% are used for testing). The “split-data” approach is a cross-validation method; thus, it tests model fit, not the model's temporal transferability.
The “split-data” approach is limited by autocorrelation between calibration and validation data sets (Araujo et al. 2005a). Modeling methods assume the modeled occurrences are independent and thus suitable for model validation, but this is not the case when the data used to calibrate and validate the model are collected from the same modeled region. For example, the spatial models of Anopheles mosquitoes in the Caribbean were validated by randomly selecting 36 points from the original 350-point data set, which were then treated as “independent test points” (Fuller et al. 2012a). Yet, randomly selected subsets of the original data set do not become independent because the environmental determinants of species distributions are strongly autocorrelated with the data used for model calibration.
This issue was shown when model validation using independent versus nonindependent data found the latter consistently resulted in higher estimations of model accuracy (Araujo et al. 2005a). The use of independently collected vector test data from the same modeled area in different time periods (e.g., Ogden et al. 2008) does not correct this issue but only introduces the risk of temporal autocorrelation due to possible lack of independence between data points adjacent through time (Araujo et al. 2005a, Rapacciuolo et al. 2014). Therefore, species distribution models may sometimes perform no better than random models, but they predict spatial distributions accurately purely because of underlying spatial or spatiotemporal autocorrelation in predictor variables (Bahn and McGill 2013). It then becomes possible to fit environmental factors to almost any spatially autocorrelated species distribution, making it difficult to determine whether a distribution arises because the vector responds biologically to that factor or because both the species distribution and environmental factors share a similar autocorrelative structure. Species distributions, including those of disease vectors like mosquitoes, depend strongly on environmental factors that are highly autocorrelated both spatially and temporally (e.g., Duncombe et al. 2013). As a consequence, any other variable that shows similar underlying spatial autocorrelative structure may appear, spuriously, as a causal variable in a model.
Validation efforts that limit spatial and temporal autocorrelation by drawing model testing data from different areas and/or times are more likely to be effective (Araujo et al. 2005a, Botkin et al. 2007). Developing statistical techniques to measure spatial autocorrelation explicitly can also increase confidence that the model is predicting accurately and capturing critical biological processes (Kerr et al. 2011). Unraveling the relative roles of purely spatial variables, purely environmental causes, and the overlapping contributions of space and time may require additional statistical analyses, such as variance decomposition (Legendre and Legendre 1998). Moran's I or Geary's c are also useful for evaluating the strength of spatial autocorrelation in datasets used in these models (see Dormann et al. 2007).
In the absence of independent data, many studies we assembled were forced to rely on area under the receiver operating characteristic (AUC) as a surrogate for model validation, with models considered validated if they received high AUC values for their training and testing data (e.g., Moo-Llanes et al. 2013, Garza et al. 2014, Gonzalez et al. 2014). However, AUC tests model fit and does not perform true model validation. The AUC's measure of a statistical model's fit is derived from presence–absence species data, and this method can overestimate model accuracy (for details on these problems, see Lobo et al. 2008). The true skill statistic (TSS), however, is threshold-dependent (i.e., uses continuous probabilities of presence) and appears to provide more reliable tests of model fit than AUC (Allouche et al. 2006). Ideally, multiple test statistics should be used to assess model performance (Elith and Leathwick 2009); methods such as AUC and TSS should be used to assess model fit, whereas model tests of spatial and temporal transferability using independent observations should be used to test those models more strongly.
A second method of validation is the use of independent data (i.e., data not used for model training or tests of model fit), and it is the ideal validation approach for predictive models to test the model's transferability to independent events (Araujo and Guisan 2006, Botkin et al. 2007). Independent data sets can come from a geographically distinct area (Fielding and Haworth 1995, Randin et al. 2006), or from a different time period (Araujo et al. 2005a). For instance, Anopheles mosquito occurrence records collected between 1930 and 1940 from northeastern Brazil were used to validate present-day spatial models of these mosquito species in sub-Saharan Africa (Tonnang et al. 2010). Yet, correlative disease vector forecast models rarely use temporally independent data for model validation (e.g., Brownstein et al. 2005), usually due to data scarcity.
We found that studies using mechanistic models to predict changing disease vector distributions were more likely to employ independent field observations for validation, with six of the nine mechanistic model studies using this approach (Table 1). Studies employing correlative models (e.g., Beebe et al. 2009, Drake and Beier 2014) did so more rarely. Another variation of independent validation involved comparison of the study's spatial models with predicted suitability maps from other sources from that same time period. For example, the Tsetse Ecological Distribution model (see Fig. 1), used to predict the sleeping sickness vector Glossina morsitans in Kenya, was validated by comparing it to a combination of tsetse fly habitat suitability maps from other sources (DeVisser et al. 2010, Messina et al. 2012). However, agreement between the model's output and available tsetse habitat suitability maps is low, likely due to variation in how each map was constructed and what each map represents (e.g., maps of historical ranges, species distribution models, etc.).
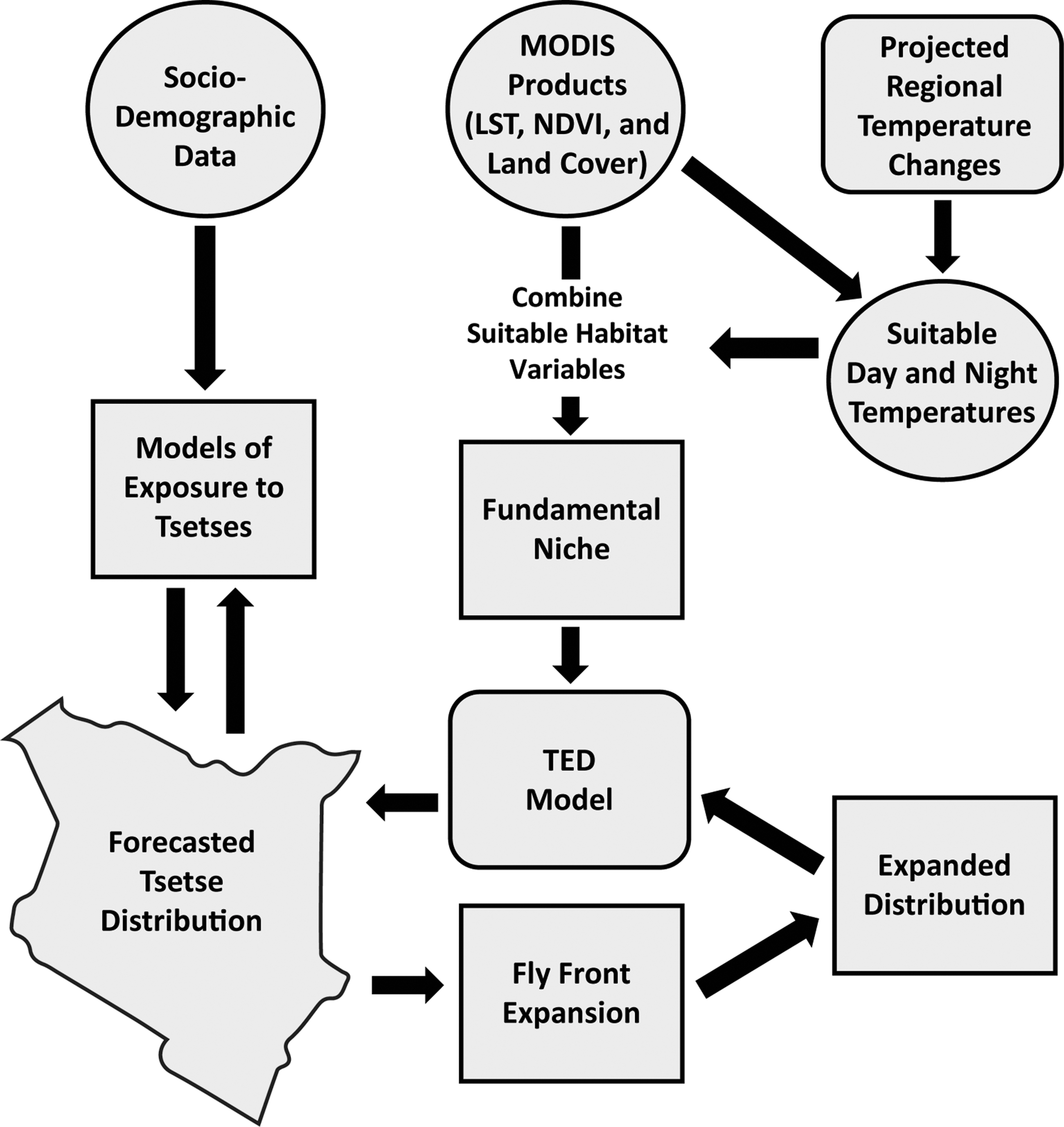
The African Trypanosomiasis Forecasting System (ATcast) modeling outline. Incorporated in this modeling framework is the Tsetse Ecological Distribution (TED) Model, which accounts for spatio-temporal dispersal dynamics of tsetse fly distributions, among various other climatic and nonclimatic factors. Projected regional temperature changes are calculated using the Regional Atmospheric Modeling System (RAMS). MODIS, Moderate Resolution Imaging Spectroradiometer; LST, land surface temperature; NDVI, Normalized Difference Vegetation Index. (Adapted from Messina et al. 2012.)
Many of the studies we reviewed claimed insufficient data for validation (e.g., Martens et al. 1999, Levine et al. 2004, Estrada-Pena and Venzal 2007, Gonzalez et al. 2010, Tonnang et al. 2010, Messina et al. 2012). While mechanistic modeling of disease vectors is one way to counter this problem, independent occurrence data are still needed to validate the models. Paleoecological observations have value for model validation for some species (Martinez-Meyer and Peterson 2006, Willis and Birks 2006, Roberts and Hamann 2012), but their potential for arthropod vectors is limited because some specimens are rarely preserved and because environmental conditions at the time of their preservation can be too uncertain to provide insight into present-day environmental dependence (Borkent and Grimaldi 2004). This also reveals the added challenge of modeling most emerging vector species (e.g., mosquitoes carrying West Nile virus; e.g., Yoo 2014), especially with the historical calibration approach: Historical records will be largely lacking for places of concern. For these cases, historical calibration might not be possible until sufficient vector records have been collected.
A third validation approach was the comparison of occurrence data of arthropod vectors with the locations of infected reservoir species, and it holds significant promise (Table 1). For example, models of phlebotomine sand flies were validated using disease case data on leishmaniasis, for which dogs are a reservoir (Galvez et al. 2011). Leishmaniasis case data were collected from veterinary clinics scattered across 32 villages in Madrid. The number of dogs infected with leishmaniasis, and the locations of the clinic in which they were diagnosed, were recorded. Their locations were then geocoded into a map layer and compared with the map of phlebotomine sand fly vectors and disease presence and predicted phlebotomine distributions were found to be strongly related.
Although the presence of a disease suggests the nearby presence of the vector that transmits it, such as leishmaniasis in dogs carried by the phlebotomine fly, areas where the disease is detectable are commonly subsets of a vector's geographical distributions (Kulkarni et al. 2010). Furthermore, disease cases may be aggregated around the health centers where they were diagnosed and recorded, offering biased measurements of the geographical distribution of disease incidence and, consequently, vector distributions. For example, the human African trypanosomiasis (HAT) vector G. morsitans still inhabits many regions of Tanzania, but HAT is now mainly found in the western and northern regions of the country (Matemba et al. 2010, Malele 2011).
Projecting vector distributions into the future
Studies we reviewed projected future changes in disease vector distributions by substituting future environmental changes into their current spatial models in one of three ways—using GCMs, adding a constant increment of environmental change uniformly throughout the region of interest, or extrapolating recent environmental trends linearly into the future (see Table 1). First, 24 of the 29 papers we reviewed used GCMs to model climate on the basis of its underlying physical processes and used these models to predict geographical variation in future climates under different emissions scenarios. For instance, Lyme disease risk in North America was predicted by modeling Ixodes scapularis tick distributions with current climate data and tick occurrence records and then projecting the current spatial model to 2020, 2050, and 2080 using GCMs in combination with emissions scenarios (Brownstein et al. 2005). While GCMs are believed to remain the most advanced tools available for future projections of climate change (Randall et al. 2007), their outputs still fail to address changes in biological processes through time when the GCMs are projected from only one spatial model.
Second, the use of constant environmental increments set by the study, such as the input of 2°C, 0.1°C, or 4°C increases in temperature combined with 10% increases or decreases in seasonal rainfall across Africa to predict future Anopheles mosquito distributions (Tonnang et al. 2010), assumes that the spatial environmental patterns will remain constant through the modeled period. Retaining constant spatial trends in environmental factors using this approach will lead to discontinuities in predicted vector distributions if modeled environmental differences exceed thresholds of tolerance among vector species in some areas. For this reason, detailed knowledge of the vector's ecology is imperative to these modeling approaches. Information on temperature thresholds for survival is gathered through field observations, such as for the Asian tiger mosquito, Aedes albopictus (Fischer et al. 2014). Upper and lower critical temperatures can also be tested in laboratory settings and then incorporated in future climate scenarios. However, results for survival thresholds can vary substantially between field and laboratory conditions. Because ecological complexity, such as short-term temperature fluctuations, can affect results of survival thresholds in the field, both field and laboratory results should be taken into account when modeling geographic limitations (e.g., Brady et al. 2013). While such measurements are likely to correlate with species' realized thermal niche limits beyond laboratory confines, species' capacities for behavioral thermoregulation are variables among species and among environments (Sunday et al. 2014).
Third, extrapolating from recent environmental trends by adding a constant change increment (e.g., +1°C) is an improvement to setting theoretical increments because its future projections are grounded on observed trends through time. For example, change in irrigated cropland in Sudan and Upper Egypt was first calculated between 2001 and 2009 and then extrapolated to 2050 to predict future Anopheles arabiensis mosquito distributions (Fuller et al. 2012b). However, this method assumes these trends will only continue linearly.
Determining which projection method is most realistic will remain uncertain because the future is uncertain, but ensemble forecasting is a promising solution to increasing forecast robustness (Araujo and New 2007). Multiple model outputs are analyzed for their range of projections. Forecast models derived from different future scenarios are combined using consensus methods to produce outputs and are predicted to be more accurate than forecasts from single models (Forester et al. 2013). Ensemble forecasts can and are already being used in epidemiological forecast models. For instance, projections of Ae. albopictus in Europe used 10 regional climate models and achieved climate projections consistent with the Intergovernmental Panel on Climate Change (IPCC) projections and future hot spots consistent with other published studies (Caminade et al. 2012). But regardless of the method used to project future environmental change, a forecast model will lack credibility unless the magnitude of species' temporal response to environmental change can be approximated.
Each of the projection modeling frameworks we reviewed requires the space-for-time assumption (Pearson et al. 2006, Fisher et al. 2010). That is, observed spatial environmental gradients within the species' current range are assumed to be identical to how it would respond to the temporal environmental changes of similar magnitude. Models are then extrapolated to spatial and temporal domains different from those in which the models were trained (Dobrowski et al. 2011, Kerr et al. 2011). However, spatially accurate models may not be temporally transferrable. Slight differences in the current distributions of species predicted by different models become magnified when those distributions are then projected into future climate scenarios, resulting in a variety of predicted range sizes for the same species and region and increasing projection uncertainty (Kharouba et al. 2009).
If species distributions are at or near equilibrium with environmental conditions (i.e., the current areas where the population is found represent the full possible extent of the population's distribution), it is more likely that models will correctly predict their distributions through time (Botkin et al. 2007, Broennimann and Guisan 2008, Elith and Leathwick 2009, Araujo and Peterson 2012, Pagel and Schurr 2012, Sax et al. 2013, Wenger et al. 2013). Yet, disease vectors are often likened to invasive species in their ability to spread rapidly into new environments through human transport and changes in land use and climate, and equilibrium assumptions are likely to be unreliable (Crowl et al. 2008, Fuller et al. 2012b). The equilibrium assumption may mislead predictions over broad areas, depending on species' traits (e.g., dispersal), so species' capacities to colonize previously unoccupied areas may affect reliability of model predictions (Araujo and Pearson 2005, Leroux et al. 2013).
Understanding dispersal limitations can be crucial. For example, correlative modeling of the potential spread of Ae. albopictus to higher altitudes in the Alps was assessed by relating presences and absences of the mosquito to high-resolution (200-meter) land surface temperature data (Roiz et al. 2011). A temperature gradient–based model was created to fill the no-data areas and, combined with threshold conditions governing egg and adult overwintering survival, the models agreed with previous studies' results of annual and January mean temperature as the most crucial to mosquito dispersal. The authors did not discuss their validation method but do admit that model validation with empirical observations is needed and that they will validate with independent field data as they conduct new vector collections in the coming years. Mechanistic modeling of G. morsitans tsetse flies in Kenya, for example, considers both environmental factors and fly dispersal abilities (Messina et al. 2012). The modeling framework accounts for fly dispersal by setting a “fly front” (i.e., tsetse population movement as a whole, instead of individual tsetse movement rates), where interactions between tsetse distributions and predicted environmentally suitable areas would be considered tsetse presence. The end result consists of 250-meter spatial and 16-day temporal resolution presence–absence tsetse maps (see Fig. 1), validated with tsetse habitat suitability maps from other sources. A cost analysis estimates that if such a predictive modeling system is implemented it will, over an 8-year tsetse management campaign, save Kenya's control programs over $19 million or approximately 58% of normal tsetse control expenditures (McCord et al. 2012).
Historical calibration
A validation method in macroecology that relies on historical calibration of model predictions (Kharouba et al. 2009) is achieving general acceptance in conservation and global change research (Rapacciuolo et al. 2014; described in Fig. 2). We found no studies that have yet used this framework for predicting changing vector distributions through time, so we describe its utility by drawing on examples from macroecological and global change research. The key benefit of this framework is to evaluate the reliability of the space-for-time assumption and provide a direct test of whether spatial models are temporally transferrable in the context of a natural, or pseudo-, experiment (Kerr et al. 2007).
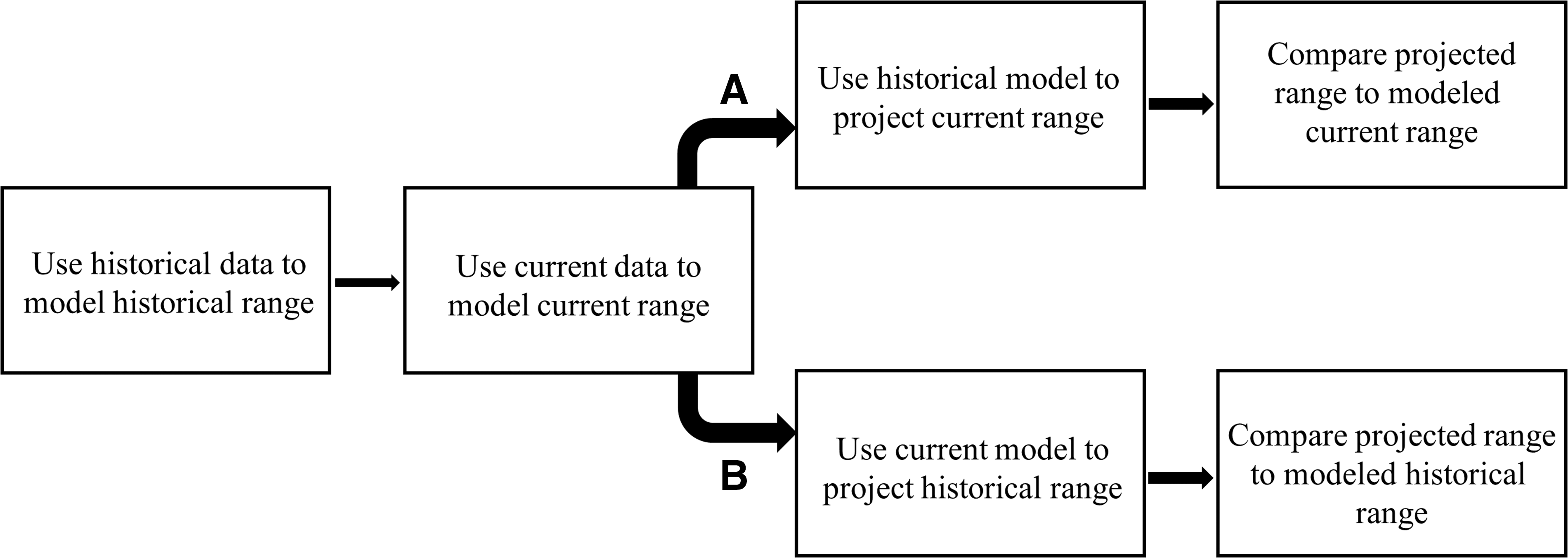
Methodology applied when using historical species occurrence data to validate species niche models through time (adapted from Kharouba et al. 2009), where models can be forecasted (
Historically calibrated distribution models employ the usual tests of model fit, based on cross-validation and statistical accuracy assessment (e.g., AUC and TSS). However, predictions of how a species' distribution will change through time are tested through a period during which such changes can be observed directly. For example, butterfly distributions across Canada were examined in a historical period (1900–1930; Kharouba et al. 2009). Each of these species' range responses to subsequent climate change was tested by substituting observed climate measures from 1960 to 1990 into this historical model and predicting how their ranges should have changed if historical relationships between climate and species' occurrences remained constant. Predictions for the latter time period (1960–1990) could then be tested in two ways. The first involves recalculating model accuracy statistics based on new occurrence points from that latter time period. The second involves constructing new models from latter observations for comparison with historical model predictions. Predictions of butterfly species distribution changes over the 20th century across Canada were strongly correlated with observed changes in those distributions. These studies assume that species do not adapt to novel environmental conditions through the observed or anticipated time periods. While that assumption appeared reasonable for butterflies in Canada, which often have one generation per year, uncertainties regarding disease vectors' potential to adapt to new environments (particularly since many breed extremely rapidly in far less seasonal environments than Canada) are substantial (Fischer et al. 2014).
The use of multitemporal species observations and environmental data reduces the uncertainty imposed by the space-for-time assumption. Minimally, using historical data to test (or validate) model predictions through time will reveal clear violations of the assumption, such as instances when models appear spatially accurate but predict nonsensically through time (e.g., Pearson et al. 2006). Yet, predicting future conditions will always require this assumption: Changes observed recently will continue to reflect changes anticipated through the future period that is being modeled. Both forecasting (Fig. 2A) and backcasting (or hindcasting; Fig. 2B) can provide some measure of historical calibration for model predictions. These historical calibration methods are finding increased application, especially in conjunction with advancing measures of species' dispersal capacities, population growth, and even species' particular behavioral thermoregulatory strategies under continuously varying environmental changes (e.g., Monahan and Tingley 2012, Sunday et al. 2012, Leroux et al. 2013, Sunday et al. 2014). Opportunities to use this framework to gain biological insights into vector responses to global change appear substantial, particularly given the increasing availability of historical species observations.
Advanced modeling techniques, rapidly evolving satellite remote-sensing tools, and increased vector surveillance provide the tools necessary for building historically calibrated vector models. For instance, forecast models of Anopheles mosquitoes are advancing with both mechanistic (e.g., Yamana and Eltahir 2013) and correlative (e.g., Tonnang et al. 2010) approaches at macroscales. Satellite-borne remote-sensing data sources, such as the Moderate Resolution Imaging Spectroradiometer (MODIS;
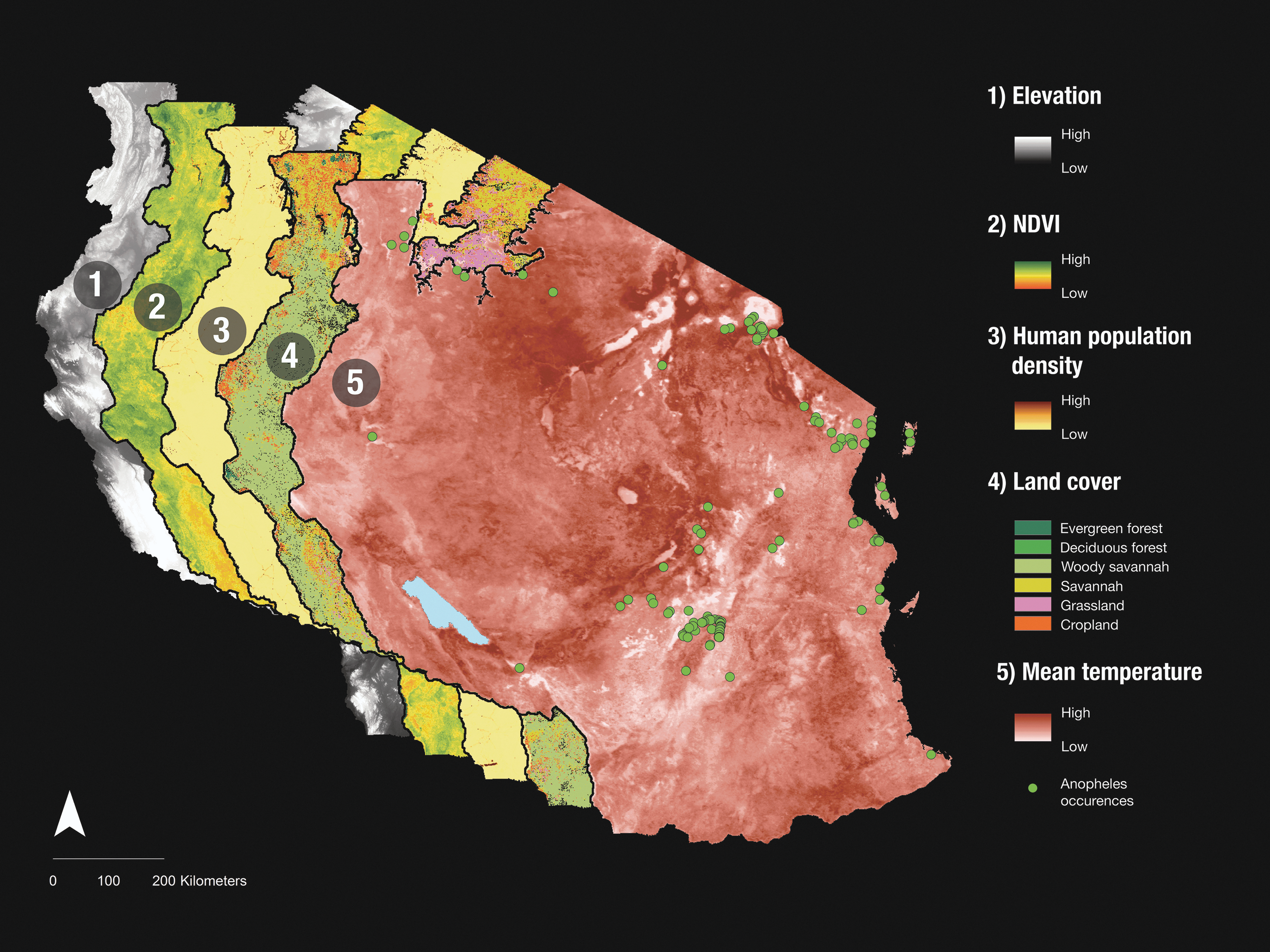
Different satellite imagery layers for mainland Tanzania in 2001 from the Shuttle Radar Topography Mission (SRTM, elevation) and Moderate Resolution Imaging Spectroradiometer (MODIS) used in conjunction with Anopheles mosquito occurrence records for predictive modeling. These satellite data can often be acquired for multiple years and even in areas where environmental data sampling is sparse.
Although historical calibration of species distribution models does not eliminate shortcomings intrinsic to predicting an uncertain future, it reduces or resolves more challenges than it creates. This historical calibration and testing framework reduces the risk that spatial autocorrelation may create misleading fits between environmental predictors and species distributions, but it cannot eliminate potential impacts of autocorrelation, which exists temporally as well as spatially. Yet, a key benefit is a direct test of the temporal transferability of a species distribution model that might otherwise provide only a snapshot of a species' reliance on environmental conditions over broad areas. When accompanied by more reliable projection methods, like ensemble forecasting, historically calibrated forecast models increasingly address the current dilemma of determining predictive model accuracy over areas where vector range changes have actually been tracked through time.
Conclusions
Validation through historical calibration is increasingly used for macroecological and global change research and could be adapted readily for epidemiological models forecasting disease vector distributions. Tests of model transferability, used with other statistical tests of model fit, will increase confidence in model predictions both spatially and temporally. Availability of reliable species observations over broad areas and at different times is a significant impediment here, as it has been for macroecological and global change research. Increased and consistent disease vector occurrence data collections are urgently needed. Forecasting disease vector distributions is further complicated by inadvertent or careless introductions of vectors into new areas (Sutherst 2004, Fischer et al. 2014). For example, chikungunya was recently found in St. Maarten in December of 2013, the first recorded instance of local transmission of the disease in the Western Hemisphere (Centers for Disease Control and Prevention 2014). Despite continuous improvements in techniques that assess how vector species' distributions and abundances depend on environmental conditions, predicting their future distributions will remain a challenging endeavor. Nevertheless, interdisciplinary opportunities to improve biological understanding of how and when vector species will respond to environmental changes could improve prospects for successful interventions to reduce their impacts on human health. Historical validation is one such opportunity.
Footnotes
Acknowledgments
We thank Andrew Plowright, Alana Taylor-Pindar, Laura Coristine, and the two anonymous reviewers for their helpful edits with the manuscript. E.S.A. acknowledges the Natural Sciences and Engineering Research Council of Canada (NSERC) for support, and J.T.K. is grateful for NSERC Discovery Grant support and to the University Research Chairs program at the University of Ottawa.
Author Disclosure Statement
No competing financial interests exist.