
Abstract
Chikungunya virus (CHIKV), a mosquito-borne alphavirus of increasing public health significance, has caused large epidemics in Africa and the Indian Ocean basin; now it is spreading throughout the Americas. The primary vectors of CHIKV are Aedes (Ae.) aegypti and, after the introduction of a mutation in the E1 envelope protein gene, the highly anthropophilic and geographically widespread Ae. albopictus mosquito. We review here research efforts to characterize the viral genetic basis of mosquito–vector interactions, the use of RNA interference and other strategies for the control of CHIKV in mosquitoes, and the potentiation of CHIKV infection by mosquito saliva. Over the past decade, CHIKV has emerged on a truly global scale. Since 2013, CHIKV transmission has been reported throughout the Caribbean region, in North America, and in Central and South American countries, including Brazil, Columbia, Costa Rica, El Salvador, French Guiana, Guatemala, Guyana, Nicaragua, Panama, Suriname, and Venezuela. Closing the gaps in our knowledge of driving factors behind the rapid geographic expansion of CHIKV should be considered a research priority. The abundance of multiple primate species in many of these countries, together with species of mosquito that have never been exposed to CHIKV, may provide opportunities for this highly adaptable virus to establish sylvatic cycles that to date have not been seen outside of Africa. The short-term and long-term ecological consequences of such transmission cycles, including the impact on wildlife and people living in these areas, are completely unknown.
Introduction
C
In Africa, CHIKV is a zoonotic arbovirus with a life cycle that principally involves primates and Aedes mosquitoes. Although there are reports of virus or antibody being detected in nonprimates, these species likely play no significant role in the normal transmission cycle. Although the idea that birds may play a role in the transmission cycle has been mentioned in many papers, what seems to be a misconception has been based on the statement by Moore et al. (1974) that “In March 1968, an isolate was recovered from a golden sparrow (Ayipasser luteus) captured at Lake Chad in northern Nigeria (G.E. Kemp, personal communication).” Subsequent surveillance has indicated that birds are not involved in the transmission cycle, and, indeed, it is known that the primary vectors rarely if ever feed on birds (Faraji et al. 2014). On the basis of data from many recent studies, it really is time to stop perpetuating a myth, sometimes fueled by a public perception (that I have personally heard often), that the name chikungunya must be derived from an association with chickens. In Asia, no sylvatic cycle has been observed. CHIKV is similar to dengue viruses (DENV) where humans can develop a relatively high viremia. Because of this, epidemics can be sustained strictly via human–mosquito transmission. Symptoms of CHIKV often resemble those of dengue fever; in dengue-endemic regions, it may be misdiagnosed and therefore underreported (Carey 1971). However, the arthritogenic nature of CHIKV differs from the febrile illness shared with other arbovirus infections and often results in recurrent polyarthritis that persists up to several years. Such chronic clinical manifestations have been found correlated with the observation that CHIKV can often cause persistent infection in muscle and joint tissues (Hoarau et al. 2010, Hawman et al. 2013, Rohatgi et al. 2014).
During interepidemic periods, CHIKV is maintained in Africa via a sylvatic transmission cycle involving a number of species of mosquitoes including: Aedes (Ae.) aegypti, Ae. africanus, Ae. luteocephalus, and Ae. furcifer-taylori, and wild primates (Jupp and McIntosh 1988). This is in contrast to Asia where the virus is maintained in cycles between Ae. aegypti or, most recently, Ae. albopictus and humans (Jupp and McIntosh 1988). These vectors are also widely distributed. Indeed, since the 1980s, Ae. albopictus has invaded and become established in many parts of the world, including the Americas and Europe (Benedict et al. 2007, Schaffner et al. 2013). The range expansion, primarily due to human activities related to global commerce, together with viral evolution described below, has been a major contributory factor to the spread of CHIKV.
CHIKV Molecular Evolution and Selection in Mosquitoes
CHIKV and ONNV are closely related and share conserved sequences and antigenic epitopes; however, CHIKV is transmitted by Aedes spp. mosquitoes and ONNV is a unique arbovirus in that is primarily transmitted by Anopheles (An.) gambiae. Therefore, an early molecular characterization of CHIKV prior to the 2006 epidemic focused on the mechanisms governing the vector specificity of both viruses (Vanlandingham et al. 2005, 2006). Using cDNA infectious clones, the chimerization of genetic materials between CHIKV and ONNV first demonstrated that the distinction in vector specificity for Ae. aegypti by CHIKV is determined by genetic sequences in the structural genes (Vanlandingham et al. 2006). More recent evidence has suggested that the vector specificity for An. gambiae by ONNV is controlled by the genetic loci in its nsP3 gene (Saxton-Shaw et al. 2013). However, the impact of single genetic loci in both regions still remains to be characterized.
It has long been known that RNA viruses have an enormous capacity for genetic variation due to the high error rate of RNA virus polymerases. Geographically isolated lineages often evolve, occasionally changing the vector or host specificity. Quasi-species populations generated during replication may contain a variant that is selected due to a competitive advantage, such as the ability to replicate to higher titers in a mosquito or vertebrate host, or to extend its host range. This was recently demonstrated in CHIKV, where a single mutation, alanine to valine at position 226 in the E1 envelope glycoprotein gene (A226V E1), facilitated CHIKV replication in and therefore transmission by the highly anthropophilic Ae. albopictus (Tsetsarkin et al. 2007, 2011b). Not only did this mutation facilitate the explosive Indian Ocean epidemic, but also allowed substantial geographic expansion of CHIKV throughout sub-Saharan Africa, Southeast Asia, and into Europe (Thiberville et al. 2013). Phylogenetic analysis of numerous CHIKV sequences has identified three geographically associated genotypes: West African, ECSA, and Asian. The ECSA and Asian strains were calculated to have diverged within the last 150 years, with the Asian clade splitting into an extinct Indian lineage and the currently circulating Southeast Asian strains. Recent Indian Ocean isolates form a monophyletic lineage descending from the ECSA clade. The strains in this lineage contain three positively selected codons—two in the capsid protein at codons 23 and 27 and the E1-A226V mutation critical for the adaptation of some CHIKV strains to Ae. albopictus (Volk et al. 2010). The ongoing epidemic in the Caribbean sustained by Ae. aegypti involves the Asian genotype (Van Bortel et al. 2014), which does not have the A226V mutation responsible for a high infectivity for Ae. albopictus. In November, however, it was reported that the ECSA genotype of CHIKV was detected in Brazil (Maron 2014). Although isolates did not contain the A226V mutation, nature has already demonstrated multiple independent occurrences. In Brazil, only Ae. aegypti is involved in CHIKV transmission (Teixeira et al. 2015). However, if Ae. albopictus begins to play a role as a vector in the Americas, for example in Panama where it is common (Miller and Loaiza 2015), it is certainly conceivable that there may be selection for the more transmissible mutated form of CHIKV.
Laboratory studies have found that Ae. albopictus is a competent vector for CHIKV (Tesh et al. 1976, Turell et al. 1992); however, it had not been implicated as a major vector in previous CHIKV epidemics. Tesh et al. (1976) infected 16 different geographic strains of Ae. albopictus with the prototype ECSA Ross strain and the Barsai strain of CHIKV. Turell et al. (1992) infected 10 strains of Ae. albopictus and seven strains of Ae. aegypti with the 15561 CHIKV strain (originally isolated in Thailand from human serum in 1962). This study found that mosquitoes collected at various locations possessed similar characteristics for vector competence when fed 5.3 Log10 plaque-forming units (pfu)/mL CHIKV: 70–100% of Ae. albopictus and 14–28% of Ae. aegypti were infected at day 14 postinfection (Turell et al. 1992). Laboratory-based vector competence studies have demonstrated transmission by other species, including Ae. fulgens, Ae. furcifer, Ae. togoi, Ae. triseriatus, Ae. vexans, Ae. vittatus, Eretmapodites chrysogaster, and Opifex fuscus (for review, see Coffey et al. 2014)
The absence of Ae. aegypti on Reunion Island led to the hypothesis that a change in the viral genotype may have affected infectivity for Ae. albopictus during the 2005 Indian Ocean epidemic (Enserink 2006, Reiter et al. 2006, Schuffenecker et al. 2006). This was confirmed by sequence analysis and vector competence testing of strains collected during the epidemic (Tsetsarkin et al. 2007, 2011a) and collection and testing of vectors from affected areas. A total of 240 mosquito pools were collected by the Direction Régionale des Affaires Sanitaires et Sociales (DRASS) and processed by the Service de Santé des Armées (SSA), Marseille. On the basis of CHIKV-positive pools of Ae. albopictus, it was suspected that Ae. albopictus was the main vector of CHIKV on the island. During this time, Ae. albopictus was also identified as a vector in places where Ae. aegypti was also prevalent, including Madagascar, India, and Gabon (Thiberville et al. 2013).
Multiple factors may have contributed to recent epidemics, including exposure of a susceptible naïve population, the abundance of larval habitats, and climate. In the Americas, the relatively low incidence of infections involving other alphaviruses, for example Mayaru (Zuchi et al. 2014), may also facilitate spread of CHIKV, because any potential for cross-protective immunity is low. Human epidemics of CHIKV in Africa historically coincided with rainy periods that increased sylvatic vector densities (Lumsden 1955), but outbreaks along the coast of Kenya in 2004 followed a period of unusually warm and dry conditions, during which time improper water storage may have facilitated vector breeding, and increased temperatures could have enhanced CHIKV transmission by mosquitoes breeding in close proximity to people (Chretien et al. 2007, Gould and Higgs 2009). The CHIKV strain implicated in the large Indian Ocean epidemic was related to previous ECSA isolates, and the Indian Ocean epidemic followed the outbreaks in Kenya, Comoros, and Seychelles, as well as some regions of Madagascar in 2004 (Schuffenecker et al. 2006, Chretien et al. 2007, de Lamballerie et al. 2008). The A226V E1 mutation occurred during this outbreak and facilitated the rapid spread of CHIKV via Ae. albopictus. In Ae. albopictus, CHIKV infectivity was significantly higher and viral dissemination and transmission to suckling mice was more efficient for CHIKV with valine at E1-226, but this mutation had no significant effect on CHIKV transmission by Ae. aegypti (Tsetsarkin et al. 2007).
Interestingly, the A226V E1 mutation has arisen independently at least four times in response to a requirement of transmission by Ae. albopictus (i.e., in areas without Ae. aegypti or areas populated by Ae. albopictus and Ae. aegypti). The result is that Ae. albopictus has become a major CHIKV vector (de Lamballerie et al. 2008, Volk et al. 2010, Tsetsarkin et al. 2011a). Although the A226V E1 mutation itself modulates CHIKV infectivity and transmission by Ae. albopictus, this mutation is limited to CHIKV ECSA strains, and other mutations in E1 and E2 have been found to block the A226V E1-mediated adaptation to Ae. albopictus: E2-T211I in most ECSA strains and E1-A98T in all endemic Asian strains (Tsetsarkin et al. 2009, 2011a). An additional mutation, E2-L210Q, was subsequently found to increase CHIKV dissemination in Ae. albopictus but had no effect on CHIKV fitness in Ae. aegypti (Tsetsarkin and Weaver 2011). Recent work suggests that additional adaptive mutations have arisen in the envelope protein of CHIKV strains that have increased fitness in Ae. albopictus. It has been predicted that combinations of these additional adaptive mutations would evolve in endemic strains in India and Southeast Asia, mediating even greater fitness in Ae. albopictus, and that these strains would spread globally (Tsetsarkin et al. 2014). Interestingly, an outbreak in 2013 involving the Asian clade of CHIKV on Yap Island in the Federated States of Micronesia involved Ae. hensilli (Savage et al. 2014). Virus detection in one pool of male mosquitoes suggested vertical transmission. A laboratory study demonstrated that Ae. hensilli was also highly susceptible to infection with a Comoros 2005 strain of the ECSA clade (Ledermann et al. 2014). With respect to vertical transmission, although there have been occasional reports from surveillance and laboratory confirmation that CHIKV may be transmitted transovarially (Agarwal et al. 2014), this seems to be such a rare phenomenon that it likely has no significance in the natural transmission cycle.
An increased vector range further increases the risk of importing CHIKV into new ecological niches through infected travelers returning from destinations experiencing CHIKV epidemics. Countries in Europe, the Caribbean Basin, and the United States, where Ae. albopictus and/or Ae. aegypti are established and widely distributed, are at risk of CHIKV establishment if a viremic person is fed upon by these vectors. There have been numerous imported cases of CHIKV into Europe (Belgium, France, Spain, Germany, Switzerland, United Kingdom, and the Czech Republic), with as many as 1000 imported cases were reported in western Europe in 2006 alone (Thiberville et al. 2013). In addition, autochthonous CHIKV transmission with Ae. albopictus as the vector was documented in Italy in 2007 (Angelini et al. 2007) and France in 2010 (Gould et al. 2010). The first cases of autochthonous CHIKV transmission in the Americas were documented in December, 2013, in Saint Martin (Vega-Rua et al. 2014). By the end of December, 2014, greater than 800,000 confirmed or probable CHIKV cases had been reported in 30 Caribbean countries/territories (Pan American Health Organization, no date). CHIKV cases have also been documented in Argentina, Belize, Bermuda, Bolivia, Brazil, Chile, Costa Rica, Ecuador, El Salvador, French Guiana, Guatemala, Guyana, Honduras, Mexico, Nicaragua, Panama, Paraguay, Peru, Suriname, and Venezuela (Pan American Health Organization, no date). Between May and December of 2013, over 2000 cases were imported into the United States, with 11 cases of autochthonous transmission documented in Florida (Fig. 1; Centers for Disease Control and Prevention 2015b). Symptoms of cases and characteristics of patients in the United States have recently been described by Lindsey et al. (2015). The realization that CHIKV could become established in Europe and/or the Americas has caused considerable concern; CHIKV was added to the list of diseases requiring mandatory notification in France and was designated as a Category C agent by the National Institute of Allergy and Infectious Diseases (NIAID), emphasizing the need for more research on the virus.
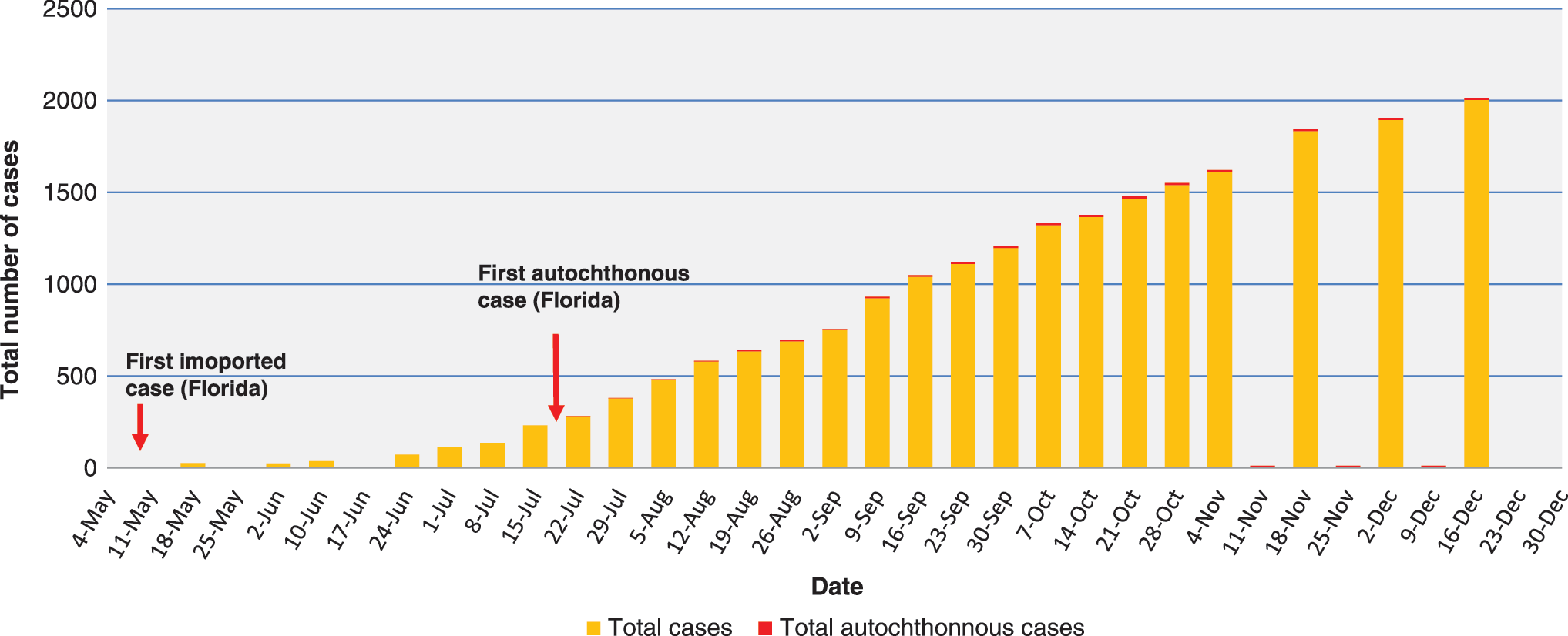
The total number of imported and autochthonous cases of chikungunya based on data from ArboNET. Note: Data were not available for June 17, November 18 and 25, and December 9, 23, and 30.
Mosquito Responses to Infection
CHIKV is associated with a high case-to-infection ratio, with most people who have been infected developing symptoms. However, as with other mosquito-borne viruses, the vectors display no overt signs of infection and no obvious effects on longevity or reproductive capacity despite the presence of viral titers in excess of 10,000,000 virions. As described below, although mosquitoes process the components of antiviral responses, given the large number of human cases, these would seem not to effectively control viral infection of mosquitoes in nature. Because these responses and other factors have been discussed as potential new approaches to control mosquito-borne pathogens, they are described below.
The most well-characterized invertebrate antiviral defense system is RNA interference (RNAi), which can control arbovirus replication and transmission in mosquitoes or invertebrate culture through the production of 21-nucleotide small interfering RNAs (siRNAs) from double-stranded RNA replicative intermediates recognized by the Dicer protein Dcr-2. siRNAs are used to target the viral genome for degradation in a sequence-specific manner. The modulation of the mosquito innate immune response to arbovirus infection by RNAi has been demonstrated previously for alphaviruses related to CHIKV, including Semliki Forest virus (SFV) (Fragkoudis et al. 2008, Siu et al. 2011), Sindbis virus (Myles et al. 2008, Khoo et al. 2010), and ONNV (Keene et al. 2004). CHIKV infection of Vero cells was inhibited by siRNAs against nsP3 and E1 (Dash et al. 2008) and E2 and nsP1 (Parashar et al. 2013); CHIKV replication in mice was inhibited by siRNAs against E2 and nsP1 (Parashar et al. 2013); and plasmid-based small-hairpin RNAs inhibited CHIKV replication in HeLa cells and prevented CHIK disease onset in suckling mice (Lam et al. 2012), demonstrating that CHIKV infection can be modulated by RNAi in mammalian cells. Viral siRNAs matching the CHIKV genome were produced by infected Ae. albopictus and Ae. aegypti, and higher levels of viral RNA resulted in more viral siRNA molecules and, in Ae. albopictus, more robust modulation of virus infection (Morazzani et al. 2012).
A similar RNAi pathway, the PIWI-interacting RNA (piRNA) pathway, has also been shown to have antiviral activity through a Dicer-independent mechanism that generates 25- to 30-nucleotide piRNAs (Morazzani et al. 2012, Schnettler et al. 2013). SFV infection of U4.4 (Ae. albopictus) and Aag2 (Ae. aegypti) cell lines resulted in the production of piRNA-like molecules, and knockdown of proteins involved in the piRNA pathway enhanced SFV replication (Schnettler et al. 2013). piRNA-like, viral, small RNAs were produced in the soma of CHIKV-infected Ae. albopictus and Ae. aegypti, and these small RNAs modulated the pathogenesis of a recombinant CHIKV in dcr-2 null mutant C6/36 (Ae. albopictus) cells (Morazzani et al. 2012).
Mc Farlane et al. (2014) recently evaluated the role of the RNAi pathway in mosquitoes infected with CHIKV. Knockdown/silencing of specific pathway components in vitro permitted CHIKV to replicate to higher levels, suggesting that when expressed to natural levels, these components would control replication. Infection of cells with CHIKV resulted in a suppression of the Toll signaling pathway via host cell shut-off, although antiviral activity was not mediated by either the JAK/STAT, IMD, or the Toll pathways. In vivo silencing of Ago-2 by inoculation of specific double-stranded RNA (dsRNA) resulted in a significant increase in the number of virus particles in the midgut at 4 and 7 days postinfection and more particles in the head. These effects were, however, time dependent and transitory.
MicroRNAs (miRNAs) are 22-nucleotide, endogenous, noncoding RNAs that regulate gene expression in many biological processes, including the host defense response against pathogens, at the posttranscriptional level. Jain et al. (2014) recently demonstrated that miRNA expression was differentially up- or downregulated in An. stephensi after blood feeding with or without Plasmodium parasites. The miRNA pathway appears to be important in Ae. aegypti as well, because miRNA levels were significantly modulated in mosquitoes infected with the flavivirus dengue type 2 (Campbell et al. 2014). Although no work has been published on miRNAs produced in response to CHIKV infection, replication of the alphavirus eastern equine encephalitis virus (EEEV) was restricted in myeloid-lineage cells by a host-cell miRNA that bound to a region essential for efficient infection of mosquitoes (Trobaugh et al. 2014).
In mosquitoes, phenoloxidases (PO) play a role in several key physiological functions, including cuticular sclerotization, wound healing, and melanization of parasites (Shiao et al. 2001) The PO cascade may also play a role in mosquito antiviral immunity. This cascade was activated in U4.4 cells by SFV infection, and SFV replication was enhanced when PO activity was blocked by a recombinant virus encoding an inhibitor of the PO cascade. Replication of the same recombinant SFV was enhanced in Ae. aegypti, suggesting the PO cascade also functions in vivo (Rodriguez-Andres et al. 2012).
In addition to the well-characterized RNAi response, it is noteworthy that heat shock proteins (HSPs) can also be another important physiological response of infected insects against CHIKV and closely related ONNV. Due to their function as chaperones that assist in the folding of proteins, the upregulation of HSPs has been found advantageous for the establishment of CHIKV infection in Ae. aegypti, which has been previously heat shocked in its larval stage (Mourya et al. 2004). Presumably, the upregulation of the expression level HSPs can promote the transport of nascent viral proteins in the intracellular trafficking process into the endoplasmic reticulum (ER). Such a hypothesis can also be supported by the evidence derived from the proteomic analyses of infected mosquitoes, showing the more abundant expression of cytoskeleton and ER-associated translocator genes (Tchankouo-Nguetcheu et al. 2012, Rider et al. 2013). On the other hand, because of its role in the stress response of organisms, HSP70B, for example, was also found to suppress the infection of the ONNV in An. gambiae (Sim et al. 2005, 2007).
The potential inclusion of genetically engineered mosquitoes to augment traditional mosquito control measures to combat vector-borne diseases, for example, the release of insects carrying a dominant lethal gene (Release of Insects with Dominant Lethal [RIDL®] mosquitoes), has been widely discussed (Higgs 2013, Alphey 2014). Although RIDL Ae. aegypti mosquitoes have already been released in several countries, Miller and Loaiza (2015) recently expressed concern that a consequence of the release of genetically modified Ae. aegypti in Panama may be that Ae. albopictis populations expand. The repercussions of replacing Ae. aegypti with Ae. albopictus could impact CHIKV transmission and might even drive the selection of mutant viruses. Clearly, these concerns need to be evaluated. The recent announcements that the US Food and Drug Administration is considering a proposal to release millions of RIDL Ae. aegypti, in Florida to control dengue (
Mosquito Saliva: Its Effects on the Vertebrate Immune System and Arboviral Infections
Increasing evidence demonstrates that vector saliva, including that of mosquitoes, modulates the vertebrate immune system and the course of pathogen infections (Belkaid et al. 1998, Kamhawi et al. 2000a,b, Morris et al. 2001, Schneider et al. 2004, Billingsley et al. 2006, Styer et al. 2006). Mosquitoes inject saliva into the skin during probing (Ribeiro and Francischetti 2003), and thus virus may be delivered intradermally (Turell and Spielman 1992, Turell et al. 1995). Mosquito saliva contains numerous components, many of which may be pharmacologically active (Racioppi and Spielman 1987, Ribeiro 1987, Ribeiro 1989, Kerlin and Hughes 1992, Ribeiro 1992, Ribeiro and Nussenzveig 1993, James 1994, Ribeiro et al. 1994, Champagne et al. 1995a,b, Stark and James 1996, Ribeiro and Valenzuela 1999, Valenzuela 2002, Valenzuela et al. 2002, Ribeiro and Francischetti 2003, Calvo et al. 2004, Ribeiro et al. 2004, Wasserman et al. 2004). Saliva secreted into the feeding site is often associated with hypersensitivity reactions commonly seen after mosquito feeding. Types I and III hypersensitivity reactions reflect an antibody response to salivary proteins, and type IV is initiated as a cellular response. Enhancement of arboviral infections by mosquito saliva has now been demonstrated for several viruses, including Cache Valley virus (Edwards et al. 1998), La Crosse virus (Osorio et al. 1996), Sindbis virus (Schneider et al. 2004), vesicular stomatitis virus (Limesand et al. 2000, 2003), and West Nile virus (Schneider et al. 2006). Saliva has been identified as affecting various cells and cytokines (Zeidner et al. 1997, Wanasen et al. 2004), and direct links have been established for salivary-induced effects on the immune system as a mechanism to explain enhancement of viral infections (Limesand et al. 2000, 2003, Schneider et al. 2004, 2006). Additionally, Thangamani and Wikel (2009) found the expression of various Ae. aegypti salivary gland genes was altered in response to blood feeding. Of 463 studied transcripts, 2.8–11.6% of genes were upregulated and 8–20% of genes were downregulated. Not surprisingly, known upregulated genes included defensins, mucins, and other immune-related proteins, whereas odorant-binding protein was significantly downregulated.
Thangamani et al. (2010) compared the host (murine) immune response to CHIKV delivered by mosquito bite and by needle inoculation by examining the cutaneous cytokine responses using quantitative RT-PCR. TH2 cytokines were significantly upregulated and TH1 cytokines were significantly downregulated in the skin after the bite of uninfected and CHIKV-infected Ae. aegypti. Expression of interleukin-1 (IL-2) and IL-4 were both significantly upregulated in the skin exposed to CHIKV-infected mosquito bites compared to uninfected mosquito bites, and CHIKV inoculation-induced interferon-γ (IFN-γ and Toll-like receptor 3 (TLR-3) upregulation was suppressed by the presence of mosquito saliva. Eosinophils were observed at the sites of mosquito bites but not needle inoculation, and more cells were recruited to the sites of CHIKV-infected mosquito bites than to the sites of uninfected mosquito bites. Overall, this study on CHIKV is consistent with previous studies of other viruses that demonstrated a significant role for mosquito saliva in the early infection events and immune response to arbovirus transmission (Thangamani et al. 2010). A recent study (Her et al. 2014) demonstrated a key role of TLR3-mediated antibodies with respect to CHIKV infection, replication, and pathology in the vertebrate host. This observation may explain the importance of mosquito-delivered CHIKV impacting TLR-3 expression.
Conclusions
Clearly, CHIKV is an increasing global health concern due to the recent explosive emergence of the virus in and out of Africa and Asia to naïve regions of Europe and the Americas. Diagnosis and treatment efforts are complicated by the concomitant circulation of CHIKV with the sometimes clinically indistinguishable DENV. The role of this overlap in epidemiology and transmission of both viruses remains to be seen. Considerable research efforts have been directed to understanding the epidemiology, molecular biology, and evolution of CHIKV, as well as the virus–vector interactions driving CHIKV transmission and worldwide spread. However, this trend must continue because large gaps in our knowledge about key driving factors behind the geographic expansion of CHIKV still exist. The virus is now encroaching into tropical South and Central American countries and the United States (Staples and Fischer 2014), and an important question is whether it will become permanently established? The successful adaptation of CHIKV for increased infectivity to Ae. albopictus that was mediated by a single amino acid substitution demonstrates the plasticity of this virus to exploit new niches. Therefore, one wonders if we will see sylvatic transmission cycles involving New World primates (Higgs and Vanlandingham 2015), as has occurred with the yellow fever virus. Despite enormous efforts to control yellow fever by multinational mosquito eradication programs and a highly efficacious vaccine, the virus has maintained a presence in the Americas and continues to cause rural infections and human fatalities. The zoonotic West Nile virus became rapidly established throughout the United States and in other countries largely because of its capacity to infect many species of vertebrates and mosquitoes. As we know it, CHIKV has more limited zoonotic potential; however, with new territory comes new opportunity.
With regard to controlling the spread of CHIKV and reducing numbers of CHIKV infections, at present we must rely on traditional, proven approaches, including chemical-based mosquito control, source reduction, and personal protection with repellents or behavioral avoidance. Efforts are ongoing to produce efficacious vaccines, and new strategies to suppress competent mosquito populations are becoming a reality. Given the very large number of human infections with various arboviruses that result from millions of mosquitoes being infected, one cannot help but conclude that, although laboratory studies have demonstrated the existence of mechanisms that can apparently confer some level of resistance to infection, in nature they are simply not effective in preventing viral infection. This may be because they are not as effective as laboratory experiments would seem to suggest, or it may be because the virus has evolved to suppress the mosquito innate immune responses or to be unaffected by these responses. Further studies are required to provide a better understanding of the complex interplay between viruses and their mosquito vectors (Zouache et al. 2014) so that we can develop a realistic view of whether or not manipulation of the mosquito physiological/immunological processes that are involved in the virus–vector relationship could ever really be applied as a strategy for reducing the impact of arboviruses. Effective implementation of current and new approaches to maximal effect will depend on timely surveillance and basic entomological knowledge, for example, of feeding behaviors and host preferences to allow targeting of vector species appropriately.
Clearly, CHIKV is a virus that we need to monitor and focus on so that we can better understand it and anticipate things to come. Perhaps then we can develop integrated approaches to prevent future epidemics and begin eradication in the regions of the world that it has recently invaded.
Footnotes
Author Disclosure Statement
Stephen Higgs is the Editor-in-Chief of Vector-Borne and Zoonotic Diseases.