
Abstract
Trypanosoma and Leishmania are obligate parasites that cause important diseases in human and domestic animals. Wild mammals are the natural reservoirs of these parasites, which are transmitted by hematophagous arthropods. The present study aimed to detect the natural occurrence of trypanosomatids through serological diagnosis, PCR of whole blood and blood culture (hemoculture), and phylogenetic relationships using small subunit ribosomal DNA (SSU rDNA), cytochrome b, and glycosomal glyceraldehyde 3-phosphate dehydrogenase (gGAPDH) genes. Samples from 131 wild animals, including rodents, marsupials, and bats, were sampled in six areas in the state of Maranhão, in a transition zone of semiarid climates northeast of the equatorial humid Amazon. Serological analysis for Leishmania (Leishmania) infantum chagasi was performed in opossums by indirect fluorescent antibody test (IFAT), and all animals were serologically negative. Nine positive hemocultures (6.77%) were isolated and cryopreserved and from mammals of the Didelphimorphia and Chiroptera orders and positioned in phylogenies on the basis of sequences from different genes with reference strains of Trypanosoma cruzi marinkellei and T. cruzi. From primary samples (blood and tissues) only one bat, Pteronotus parnellii, was positive to SSU rDNA and gGAPDH genes and grouped with the L. infantum chagasi branch. The studies conducted in Maranhão State provide knowledge of parasite diversity. It is important to determine the presence of trypanosomatids in wild mammals with synanthropic habits.
Introduction
A
Trypanosoma cruzi is a digenetic trypanosomatid that is characterized by significant genetic variability due to the intricacy of its population, which is composed of multiple clones displaying distinct biological properties (Buscaglia and Di Noia 2003). This species has recently been classified into six discrete typing units (DTUs) ranging from T. cruzi I (TcI to TcVI) (Zingales et al. 2009, 2012), largely distributed in humans, reservoirs, and vectors, with the emergence of one new genotype associated with bat species, designated TcBat, in Brazil and Panama (Marcili et al. 2009, Pinto et al. 2012, Zingales et al. 2012). This parasite is usually transmitted by a triatomine insect vector, and the main hosts are marsupials, edentates, rodents, chiroptera, and carnivores in sylvatic habitats, and humans, dogs, cats, and rodents in domestic or peridomestic transmission cycles (World Health Organiziation 2002, Schofield et al. 2006).
The genus comprises Leishmania protozoan parasites with a digenetic life cycle (heteroxenic), living alternately in vertebrate hosts and insect vectors (Gontijo and Carvalho 2003). In Brazil, Leishmania (Leishmania) infantum chagasi is the etiological agent primarily transmitted by the sand fly Lutzomyia longipalpis (Dantas-Torres and Brandão-Filho 2006). A great number of Leishmania species have been characterized as parasites of wild animals and are less frequently found in domesticated animals (Lainson 1988). Several studies have reported dogs as the main domestic reservoir, and skin parasitism in dogs has been associated with transmission of L. (L.) infantum chagasi to humans (Lainson et al. 1998, De Queiroz et al. 2011).
Other animals have also been investigated with regard to their role in the transmission of this disease, with major attention given to synanthropic animals, especially opossums (Didelphis spp.) (Travi et al. 1998). Bats are known to house several trypanosomatid parasites; however, few studies have investigated their potential involvement in Leishmania life cycles (Rotureau 2006). L. (L.) infantum chagasi was detected and isolated from bats in Venezuela and Brazil through PCR or indirect immunofluorescence, and positive bats were detected (De Lima et al. 2008, Savani et al. 2010).
The State of Maranhão located in a transition zone between the Amazon forest and a semiarid zone and is constantly modified by actions of human beings, whose main effect is an imbalance that can facilitate the transmission of many pathogens. Therefore, research in a wild reservoir is important for determining the presence of trypanosomatids and the risk of humans contracting the disease. The present study evaluated infection by T. cruzi and L. (L.) infantum chagasi in wild mammals in Maranhão through parasitological, serological, and molecular diagnosis and phylogenetic relationships.
Materials and Methods
Study area and small mammals caught
The study was conducted in Maranhão State, (01°01′ to 10°21′ S and 41°48′ to 48°40′W) located in northeastern Brazil. This state is located in a transitional zone of semiarid climates of the northeast to the equatorial humid area of Amazon, which is reflected in plant formations—transitioning savanna (Cerrado) in the south to seasonal forests in central and eastern areas and tropical rain forest in the northwestern area of the state. The samples were caught at six localities in seven different municipalities (Caxias/Chapadinha, Cururupu, São Domingos, São Bento, Açailandia e Barreirinhas) in distinct geographical locations in Maranhão State (Fig. 1).

Geographical origin of small mammals caught in the Maranhão state, Brazil. 1, Cururupu; 2, São Bento; 3, Barreirinhas; 4, Açailândia; 5, São Domingos; 6, Caxias/Chapadinha.
To capture the small terrestrial mammals, 20 traps (Tomahawk and Sherman) were located in longitudinal transects in forests along roads and streams, starting from the peridomestic area of the houses to the entrance into the extradomestic area or forest; bats were caught using five mist nets (7 × 3 meters). The captures were made in two 15-day campaigns in August and September of 2012 and August of 2013. The small mammals and bats that were caught were anesthetized with ketamine, and blood samples were collected by means of heart puncture.
Animals were identified using identification keys and original descriptions (Vizotto and Taddei 1973, Emmons and Feer 1997). The animals were caught and handled in accordance with the recommendations of the Brazilian Institute for the Environment and Renewable Natural Resources (ICMBio, license no. 32253-1). All procedures for handling animals were performed according to the Ethics Committee on Animal Use of the Faculty of Veterinary Medicine, University of São Paulo, Brazil.
Wild animals and blood sample collection
Animals were sedated with 1–5 mg/kg of ketamine and made aseptic with a solution of 70% ethanol before collecting samples. Two aliquots of blood were collected by cardiac puncture; part of the blood was transferred to sterile tubes containing alcohol for PCR analysis and the other for blood culture. Regarding marsupials, three aliquots of blood were collected for PCR, isolation, and indirect fluorescent antibody test (IFAT). The volumes of blood samples varied according to the body size of each animal species. After blood collection, animals were euthanized (two of each species, according the license) and tissues (spleen and liver) were harvested. Portions of each tissue were removed with the aid of single-use forceps, scissors, and scalpel blades, placed in sterile tubes containing 100% ethanol, and stored at −20°C until PCR was completed.
Serological diagnosis of L. (L.) infantum chagasi
For the blood samples collected from Didelphis, the serum was separated and stored at −20°C until tested. An IFAT was used for detection of anti–L. (L.) infantum chagasi antibodies, with a cutoff value of 1:40. Promastigote forms of the L. (L.) infantum chagasi CBT 96 strain were used as the antigen, as described by Ferrer et al. (1995). Positive and negative control sera were added to each slide.
Blood culture (hemoculture)
To detect trypanosomatid parasites, blood samples obtained from the wild were inoculated into Vacutainer tubes containing a biphasic medium consisting of 15% sheep red blood cells as the solid phase (blood agar base), overlaid by liquid liver infusion tryptose (LIT) medium supplemented with 20% fetal bovine serum (FBS) (Marcili et al. 2013, 2014). The culture was incubated at 28°C and grown in LIT medium for DNA preparation. The isolates were cryopreserved in liquid nitrogen at the Brazilian Trypanosomatid Collection (Coleção Brasileira de Tripanossomatídeos [CBT]), in the Department of Preventive Veterinary Medicine and Animal Health, Faculty of Veterinary Medicine, University of São Paulo, Brazil. Blood samples were fixed in ethanol (primary samples) for molecular detection.
Extraction and amplification of DNA from blood, organs, and culture of wild animals
The extraction of DNA from tissue was performed according to the protocol established by the Wizard SV Genomic DNA Purification Kit (Promega, Madison, WI). DNA for culture samples was extracted from the trypanosome cultures using the phenol–chloroform method. The DNA samples were subjected to the conventional PCR for trypanosomatids with barcoding on a fragment of ∼900 bp of the V7V8 small subunit ribosomal DNA (SSU rDNA) (Marcili et al. 2013, 2014), glycosomal glyceraldehyde 3-phosphate dehydrogenase (gGAPDH) (Hamilton et al. 2004), and cytochrome b (Marcili et al. 2009). PCR products of the expected size were purified and sequenced in an automated sequencer (ABI Prism 310, Applied Biosystems, Foster City, CA).
Phylogenetic analysis
The sequences obtained were aligned with sequences previously determined for other trypanosomatid species available in GenBank using ClustalX (Thompson et al. 1997) and were adjusted manually using GeneDoc (Nicholas et al. 1997). The barcoding sequences were used to construct a phylogenetic tree using maximum parsimony, as implemented in PAUP version 4.0b10 (Swofford 2002) with 500 bootstrap replicates. Bayesian analysis was performed using MrBayes version 3.1.2 (Huelsenbeck and Ronquist 2001) with 1,000,000 replicates. The first 25% of the trees represented burn-in, and the remaining trees were used to calculate Bayesian posterior probability.
Endogenous control for PCR
Ten percent of the samples that were negative for each tissue were randomly selected and subjected to PCR, as described by Ferreira et al. (2010), to amplify the cytochrome b gene of mammals, which is highly conserved.
Results
A total of 131 mammals, between rodents (n = 5), marsupials (n = 10), and bats (n = 116), were sampled in the area and examined. Regarding the rodents captured, there were six specimens of six different species, five species of marsupials, and 22 bats species (Table 1).
Hemoculture.
Serological analysis for L. (L.) infantum chagasi was performed only in opossums because it was not possible to withdraw small amounts of blood from the other small mammals. A total 10 opossums were screened and all were seronegative for L. (L.) infantum chagasi infection antibodies. Furthermore, all animals exhibited good health and no symptoms of visceral leishmaniasis (VL) at the time of clinical screening.
The overall rate of infection in wild mammals was evaluated by blood culture; 8.27% (11/131) were positive, but only 6.77% (9/131) of positive cultures were established. The positive blood cultures were from animals classified in order Didelphimorphia (Philander opossum, Gracilinanus spp., and Didelphis albiventris) and Chiroptera (Artibeus lituratus and Phyllostomus hastatus) and with a prevalence of 55.6% (5/9) and 44.44% (4/9) positive and established cultures, respectively. Positive blood cultures (two) from Pteronotus parnellii have not been established.
The morphology of all positive blood cultures was compatible with the genus Trypanosoma and metacyclic trypomastigotes of culture, similar to subgenus Schizotrypanum, except the cultures from P. parnellii, which were compatible with the subgenus Megatrypanum. The established cultures (isolates) were cryopreserved at the CBT (Table 1).
A total of 315 primary samples from different tissues were analyzed by PCR, and only one bat was positive for trypanosomatid barcoding SSU rDNA and gGAPDH genes. The sequences from primary samples and nine isolates of Trypanosoma parasites from terrestrial and flying mammals were positioned in phylogenies based on trypanosomatid barcode, gGAPDH, and cytochrome b genes and were compared with sequences retrieved from GenBank (Table 2). Congruent topologies were obtained by maximum parsimony and Bayesian analysis. The isolates were grouped as T. cruzi marinkellei and T. cruzi. T. cruzi was the most prevalent species (78% of isolates) from bats and marsupials followed by T. cruzi marinkellei (22%) in bats. All isolates of T. cruzi obtained belonged to the TcI group (100% bootstrap/100% a posteriori probability) (Fig. 2). The sequences obtained from primary samples was grouped with L. (L.) infantum chagasi (Fig. 3).
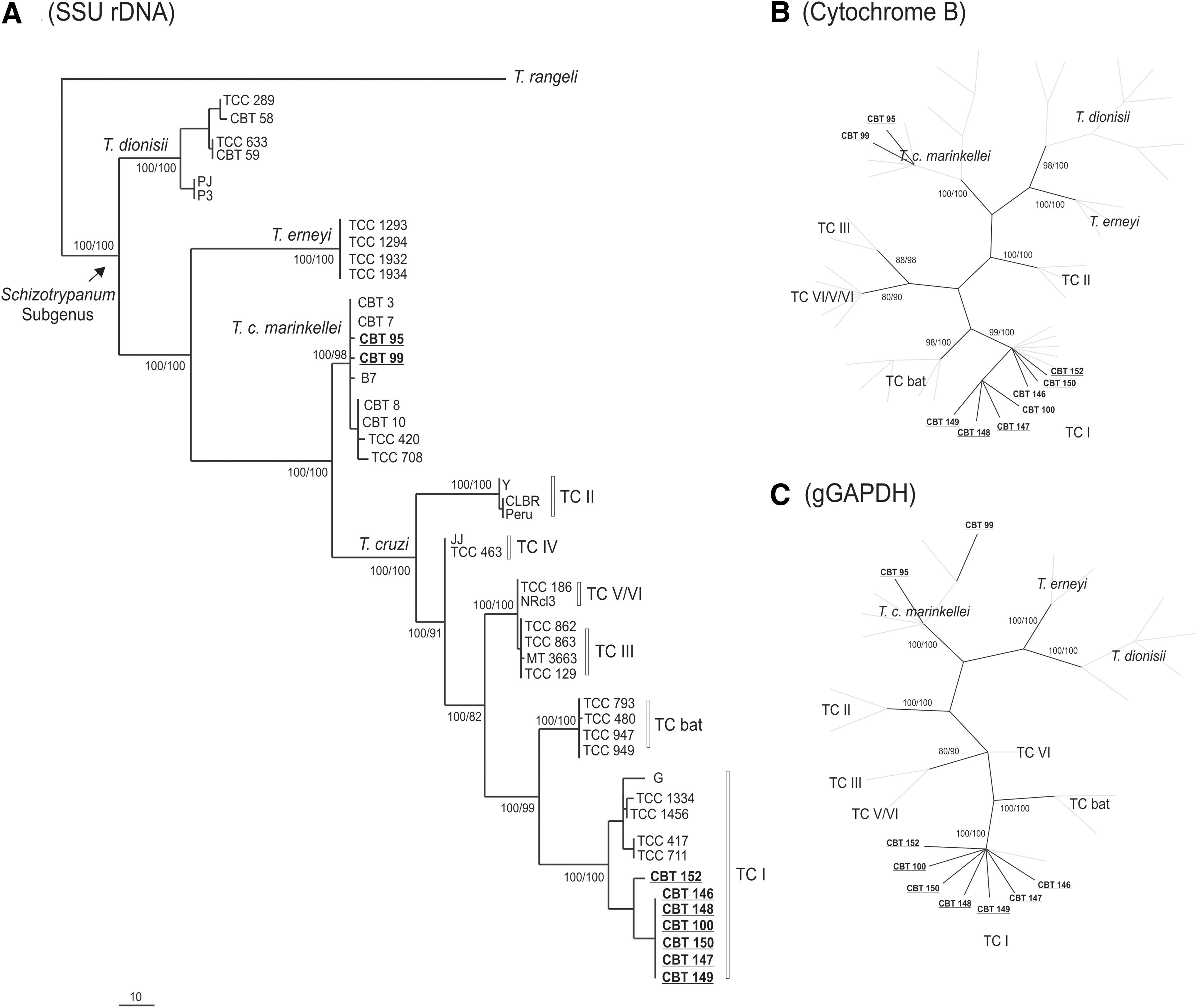
Phylogenetic trees inferred by maximum parsimony and Bayesian analysis. Numbers at nodes are the support values for the major branches (bootstrap or posterior probability) derived from 500 replicates respectively for MP and B analyses. (

Dendogram based on concatenated small subunit ribosomal DNA (SSU rDNA) and glycosomal glyceraldehyde 3-phosphate dehydrogenase (gGAPDH) sequences of six leishmania species employed for maximum parsimony and Bayesian inferences with 1276 characters. Numbers at nodes are the support values for the major branches (posterior probability/bootstrap; 500 replicates).
Sequences determined in this study and deposited in GenBank are underlined and bold.
Brazilian states: Espirito Santo (ES); São Paulo (SP); Rondônia (RO); Goiás (GO); Mato Grosso (MT); Bahia (BA); Mato Grosso do Sul (MS); Amazonas (AM); Rio Grande do Norte (RN); Tocantins (TO); Paraná (PR); Pará (PA); Acre (AC); Rio de Janeiro (RJ); Distrito Federal (DF); Piauí (PI); Rio Grande do Sul (RS); Maranhão (MA).
SSU rDNA, small subunit ribosomal DNA; gGAPDH, glycosomal glyceraldehyde 3-phosphate dehydrogenase.
Discussion
Chagas disease is one of the most important parasitoses in Latin America and a major cause of morbidity and death in people from affected countries (World Health Organization 2014). The epidemiological importance of searching for trypanosomes in wild animals is based on the need to understand all of the links in the transmission of trypanosomatids, and, thus, contribute to the establishment of appropriate standards for epidemiological surveillance and/or control in northeast Brazil.
In this study, the presence the T. cruzi I in marsupials and bats from Maranhão State were reported. This lineage/group is considered predominant in human population and causes Chagas disease in Venezuela, Colombia, and Brazil (outbreaks in Amazonia and Caatinga Region), where different clinical forms of the disease have been reported, ranging from asymptomatic to fatal both in the acute and chronic phases (Coura et al. 2002, Montilla et al. 2002, Añez et al. 2004, Teixeira et al. 2006). The T. cruzi I (TcI) lineage is initially more associated with the sylvatic cycle of this parasite, circulating among triatomines and small wild mammals, such as opossums, rodents, and primates (Marcili et al. 2009, Maia da Silva et al. 2008, Zingales et al. 2012), corroborating the findings in Maranhão State.
Marsupials are an ancient mammalian group that appeared in the Late Cretaceous Period (80 million years ago) and are probably the most ancient reservoirs of T. cruzi (Schofield 2000). Thus, animal encounters of marsupials that are positive are very important because it was observed that synanthropic animals frequently visit homes and adjacent buildings, acting as a link between the transmission of T. cruzi in sylvatic and peridomestic areas (Alvarado-Otegui et al. 2012, Coura 2013). Studies conducted in different Brazilian states, showed that the marsupials have two main roles—maintenance and transmission of T. cruzi when suffering predation and amplifying the agent and transmission for triatomine vectors (Roque et al. 2008).
The species of the Schizotrypanum subgenus were validated by molecular phylogenies (Stevens et al. 2001, Hamilton et al. 2007). Recently, a new species in the Schizotrypanum subgenus (Trypanosoma erneyi) was described in bats in Africa (Lima et al. 2012). In bats from Brazil, T. cruzi (TcI, TcII, Z3, and TcBat), T. cruzi marinkellei, and T. dionisii (Lisboa et al. 2008, Marcili et al. 2009, Cavazzana et al. 2010, Marcili et al. 2013, Acosta et al. 2014) were found and corroborated as a most ancient reservoir of T. cruzi (Hamilton et al. 2012).
In bats caught in this study, T. cruzi I and T. cruzi marinkellei were identified. The presence of TcI suggests the circulation of this lineage in this particular endemic area for Chagas disease. This fact corroborates previous observations that TcI is widespread in mammalian hosts within the sylvatic cycle. It also strengthens previous studies showing the potential epidemiological significance of bats as possible reservoir hosts for T. cruzi, due to the frequency of infection in nature, and indicating that bats may be important reservoirs and potential source of T. cruzi infection to humans (Marinkelle 1976, Marcili et al. 2009).
The positive animals (bat and marsupials) were collected in an area close to human habitation, whose shelter was the babaçu palm tree. Several studies have highlighted the importance of palm trees as natural ecotopes of triatomine vectors, especially the species of the genus Rhodnius, an important link in the chain of transmission of Chagas disease. This increases the risk of transmission by infected insects that can invade and eventually colonize houses because homes in the study region were built or refurbished mostly from the timber extracted from the forest that surrounds the community; thus, the same wood can serve as a vehicle for triatomines (Valente et al. 2000, Teixeira et al. 2001, Massaro et al. 2008).
Bats need special attention, because many species use homes as shelter areas, representing a potential trophic effect for maintaining populations of triatomines. One should take into account also that colonies of bats eventually swap shelter, thus constituting the probable elements dispersion of trypanosomiasis (Fabián 1991).
Besides Trypanosoma parasites, in bats primary samples L. (L.) infantum chagasi were also detected in P. parnellii. The first reports of the relationship between bats and Leishmania are dated 1975 and 1987, from serological studies conducted in Kenya and Egypt, respectively (Mutinga 1975, Morsy 1987). In fact, the first confirmations of this relationship occurred recently with the isolation of a strain of L. (L.) infantum chagasi from a frugivorous specimen of bat (Carollia perspicillata) in Venezuela (De Lima et al. 2008).
Bats can be an alternative source of blood to L. longipalpis, the major vector of American VL (Lampo et al. 2000). Considering the fact that the species P. parnellii is insectivorous, this species probably became infected by L. (L.) infantum chagasi following the normal pattern by the bite of an infected female phlebotomine. In experimental conditions, P. parnellii served as an alternative source of feeding to the female of L. longipalpis (Lampo et al. 2000).
Many studies have been conducted in Brazil for detection of Leishmania parasites. Serological and molecular data based on kinetoplast DNA target found L. (L.) infantum chagasi and L. amazonensis in a nonendemic area in São Paulo, Brazil (Savani et al. 2010). In Brazil, using PCR-RFLP and PCR, L. braziliensis was detected in blood samples from two bats (Molossus molossus and Glossophaga soricina) in Mato Grosso do Sul, Brazil, an endemic area (Shapiro et al. 2013). In our study, using trypanosomatid barcode (SSU rDNA) genes and gGAPDH (Marcili et al. 2014), one animal was detected as positive in a sample of liver and spleen. The use of DNA barcoding for identification and phylogeny of trypanosomes has been used in several studies (Marcili et al. 2009, Viola et al. 2009, Teixeira et al. 2011, Marcili et al. 2014). This is the first study with sequences of L. (L.) infantum chagasi from Brazilian bats positioned in phylogenetic studies. The ability of P. parnellii to fly long distances and its distribution in all Brazilian biomes, contact with humans and domestic animals, and the preference of the L. longipalpis female to feeding in this bat species became important to the public health and epidemiology of this parasite.
The importance of the use of wild species as biomarkers lies in the fact that they may reflect the impact of changes in the environment, acting as sentinels of environmental health (Dobson et al. 2006). The wild animals of this study shown to be exposed to infection reflect the presence of parasite transmission in peridomiciliary areas, making it necessary to discuss the importance that each mammalian species plays in maintaining the parasite and the use of wild mammals as a sentinel surveillance measure. Therefore, epidemiological and entomological surveillance and implementation of control programs are necessary to prevent transmission of Chagas disease and VL.
Knowledge about the involvement of mammalian and insect vectors in transmission cycles is crucial for providing measures for the control and prevention of leishmaniasis. In some places, the surveillance measures adopted have not been sufficient to decrease the number of human cases (World Health Organization 2010), thus it is necessary to investigate whether other species are supporting endemism in these localities.
Footnotes
Acknowledgments
The authors would like to thank São Paulo State Research Foundation—FAPESP for scholarships to A.P.C. and A.M. and for financial support of the project and Agência de Defesa Agropecuaria of Maranhão State for cooperation in the animal catches. S.M.G. is a recipient of a fellowship from CNPq, Brazil.
Author Disclosure Statement
No competing financial interests exist.