
Abstract
Western Burrowing Owls (Athene cunicularia hypugaea) are small, ground-dwelling owls of western North America that frequent prairie dog (Cynomys spp.) towns and other grasslands. Because they rely on rodent prey and occupy burrows once or concurrently inhabited by fossorial mammals, the owls often harbor fleas. We examined the potential role of fleas found on burrowing owls in plague dynamics by evaluating prevalence of Yersinia pestis in fleas collected from burrowing owls and in owl blood. During 2012–2013, fleas and blood were collected from burrowing owls in portions of five states with endemic plague—Idaho, Oregon, Washington, Colorado, and South Dakota. Fleas were enumerated, taxonomically identified, pooled by nest, and assayed for Y. pestis using culturing and molecular (PCR) approaches. Owl blood underwent serological analysis for plague antibodies and nested PCR for detection of Y. pestis. Of more than 4750 fleas collected from owls, Pulex irritans, a known plague vector in portions of its range, comprised more than 99.4%. However, diagnostic tests for Y. pestis of flea pools (culturing and PCR) and owl blood (PCR and serology) were negative. Thus, even though fleas were prevalent on burrowing owls and the potential for a relationship with burrowing owls as a phoretic host of infected fleas exists, we found no evidence of Y. pestis in sampled fleas or in owls that harbored them. We suggest that studies similar to those reported here during plague epizootics will be especially useful for confirming these results.
Introduction
P
Avian species that inhabit prairie dog (Cynomys spp.) colonies and other mammal burrows, and that have potential to harbor fleas, are important candidates for investigation. One such species is the Western Burrowing Owl (Athene cunicularia hypugaea; hereafter burrowing owls), which are small, ground-dwelling owls of western North America. Burrowing owls have the potential to act as vectors or reservoirs of Y. pestis for a number of reasons, including that their geographic range (see Poulin et al. 2011) overlaps areas with endemic plague, they nest widely in prairie dog colonies and burrows (Desmond et al. 2000, Restani et al. 2001, Lantz et al. 2007, Alverson and Dinsmore 2014), outside the range of prairie dogs the owls use fossorial mammal burrows including mammals with frequent exposure to Y. pestis (e.g., American badgers, Taxidea taxus; Messick et al. 1983), owls hunt rodent prey and harbor fleas (Smith and Belthoff 2001), and last, the owls make widespread migratory (e.g., Holroyd et al. 2010) and other movements that have the potential to move fleas among plague regions.
Previously, Jellison (1939) recovered 109 live rodent fleas of six species from a burrowing owl nest in a plague area near Dillon, Montana. He quoted Rucker (1909) who stated, “There is reason to believe that the booby owl, which is a constant companion of the ground squirrel, occupying the same burrows with him, may play an important role in the dissemination of the epizootic. It is thought that this bird, flying from burrow to burrow, may carry infected fleas for long distances.” Wheeler et al. (1941) subsequently collected a burrowing owl following a plague epizootic in California from which 70 sticktight fleas (Echidnophaga gallinacea) were retrieved. Upon mass inoculation into a test guinea pig, the fleas were said to be infected with plague organisms. Wheeler et al. (1941) concluded that this was the first record of a bird host as a carrier of plague-infected ectoparasites and the first demonstration of natural plague infection in this species of flea. Finally, Brown (1944) reported that burrowing owls were unusually abundant in a plague epizootic area (Hanna-Youngstown) in Alberta, Canada. In 1940 and 1941, a small number of fleas (n=47 and 37, respectively; species not identified) were collected from burrowing owl burrows, but they tested negative for Pasteurella (Yersinia) pestis. In 1942, Brown recovered a single flea from each of two live burrowing owls and identified the fleas as Oropsylla idahoensis and (tentatively) Rectofrontia fraterna. Brown (1944) noted that O. idahoensis was a known plague vector.
These studies point to a potential role of fleas found on burrowing owls in plague dynamics, although numerous aspects remain poorly understood. We were interested in determining: (1) if, and how commonly, fleas on burrowing owls harbor Y. pestis, (2) the geographic variation in flea infestations and Y. pestis detection in owl fleas, and (3) if burrowing owls might be exposed to Y. pestis via flea bites, even though owls presumably are not susceptible to plague.
Materials and Methods
During the 2012 and 2013 breeding seasons, burrowing owls (Fig. 1) were captured to collect fleas from their plumage and blood samples for diagnostic testing. Owls were captured in 10 study sites (Fig. 2) in five states: (1) Morley Nelson Snake River Birds of Prey National Conservation Area, Ada and Elmore Co., Idaho (NCA); (2) Umatilla Chemical Depot, Morrow and Umatilla Co., Oregon (DEPOT); (3) East of Baker City, Baker Co., Oregon (BAKER); (4) Yakima Training Center, Yakima Co., Washington (YTC); (5) near Pasco, Franklin Co., Washington (PASCO); (6) Umatilla National Wildlife Refuge, Benton Co., Washington (UNWR); (7) private lands northeast of Naches, Yakima Co., Washington (NACHES); (8) Fitzer/Eberhardt Arid Lands Ecology Reserve, Benton Co., Washington (ALE); (9) Rocky Mountain Arsenal National Wildlife Refuge, Adams Co., Colorado (RMA); and (10) Conata Basin, Buffalo Gap National Grassland, Pennington Co., South Dakota (BGNG). Each study site had the potential for enzootic plague, but to our knowledge there were no observable or widespread epizootic outbreaks, although there were two confirmed human cases of plague in 2012 in nearby portions of Oregon during the time of our study (Oregon Public Health Division CD Summary, Vol. 62, No. 11, May 2013; see also Kugeler et al. 2015). Prior to our study, the BGNG study site experienced an explosive plague epizootic, prompting the remaining prairie dog habitat to be treated annually with insecticide to reduce fleas and plague transmission in prarie dogs. Such treatments could have reduced the occurrence of fleas on burrowing owls and potential exposure of owls to Y. pestis in this area, but not our other study areas.

Trail camera image taken May 31, 2012, of a burrowing owl (Athene cunicularia) nest in southwestern Idaho showing adults (flying) and nestlings near a nest burrow. Adult and juvenile burrowing owls were captured at nest burrows, fleas collected from their plumage, and owl blood obtained for analysis of potential exposure to Y. pestis, which is the organism that causes plague.
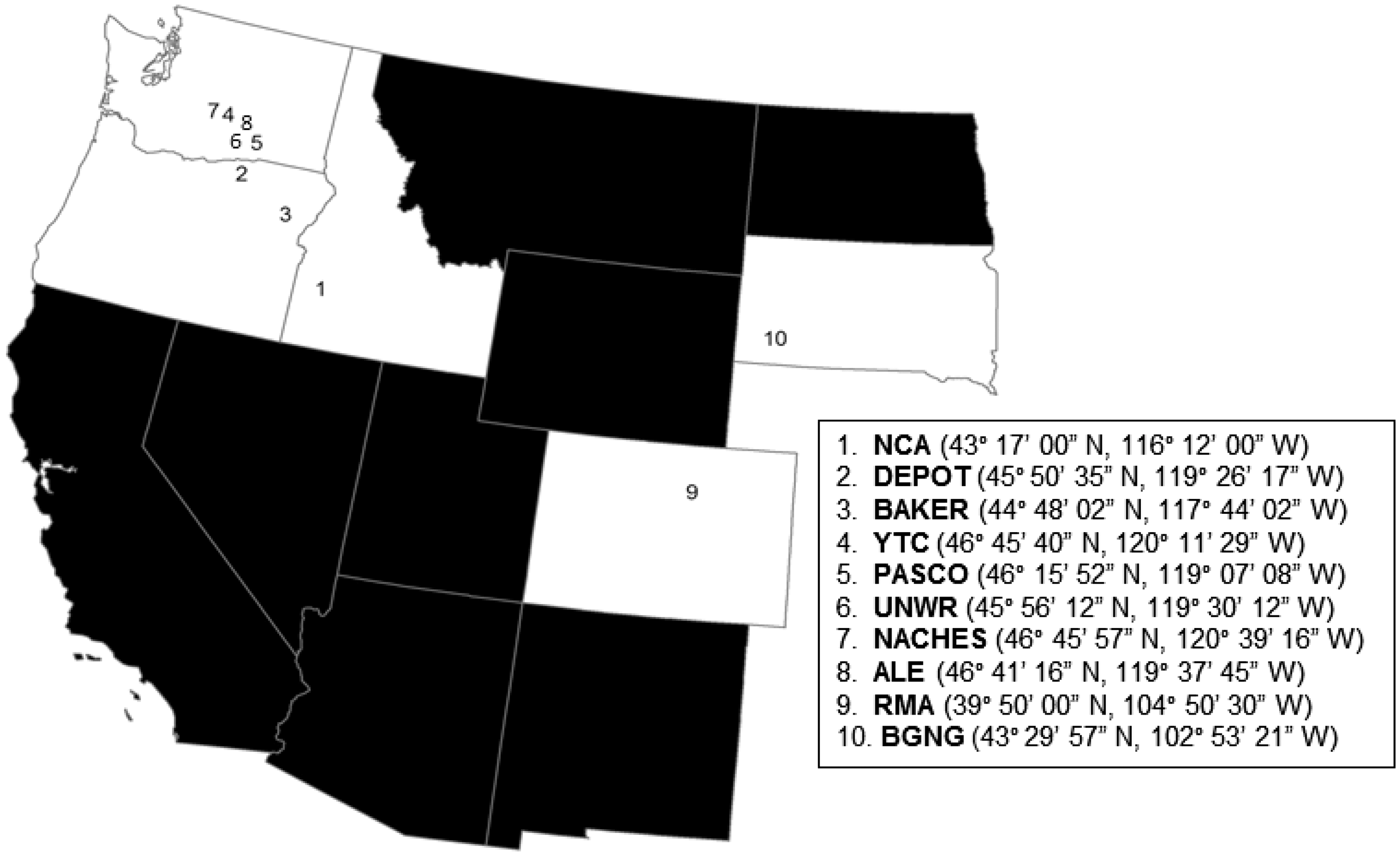
Geographic coordinates and locations of 10 study sites in five western states (white) in which potential relationships between burrowing owls, fleas, and Y. pestis were examined. See text for explanations of study site abbreviations.
Capture of owls
Burrowing owls were located by scanning appropriate habitat with binoculars from roads and while traversing fields on foot. The owls we studied in Idaho, Oregon, and Washington nested in artificial burrows (Belthoff and Smith 2003) or in natural burrows often dug by American badgers, whereas owls in Colorado and South Dakota inhabited black-tailed prairie dog (Cynomys ludovicianus) towns and nested in natural burrows. Adult owls tending nests were captured using traps placed at or near the mouth of the nest burrow or with bow nets baited with a pet-shop mouse. Owls nesting in artificial burrows were captured by hand after opening the artificial burrows or in traps as they exited nests. All captured owls were fitted with US Geological Survey aluminum leg bands for identification.
Flea collection and identification
Fleas were collected with tweezers or an aspirator directly from nestlings and adult owls and stored by nest site in Ziploc bags or scintillation vials on ice in a portable cooler. Samples were frozen at −20°C upon return from the field. Fleas were subsequently thawed, enumerated, sexed, and identified taxonomically under a dissecting microscope. Taxonomic identification included a standard clearing procedure to lighten the flea for more efficient morphological evaluation (Hastriter and Whiting 2003, Whiting et al. 2008). For clearing, fleas were soaked for several hours in a dilute concentration of potassium hydroxide and then underwent a series of ethanol washes. Once fleas were sufficiently lightened to visualize internal organ structures, they were mounted in Canada balsam on a microscope slide for final identification. This preparation allowed us to taxonomically identify reference individuals but prevented including the reference individuals in PCR tests for Y. pestis.
Nearly all fleas collected (see Results section) from burrowing owls were Pulex spp. (Siphonaptera: Pulicidae). Morphological features allow researchers to distinguish male Pulex fleas as P. irritans or P. simulans. There are currently no known morphological features in female Pulex fleas to distinguish species. Thus, we used DNA extracted from fleas, as described later, for secondary molecular confirmation to distinguish Pulex fleas. Previously published primers by Gamerschlag et al. (2008) were used to initially amplify and sequence the internal transcribed spacer (ITS) region of morphologically identified male P. irritans and comparative P. simulans specimens. Single-nucleotide polymorphism (SNP) loci were identified in the ITS region of the male P. irritans and P. simulans. These SNPs were used to develop an allele-specific PCR. Allele-specific primers (5′-GTCGAATCGCATTTTCCA-3′, 5′-ACGCTTTCGGTATTTATC-3′) were identified to discriminate between SNP alleles and P. irritans and P. simulans based on size or melting temperature (Okimoto and Dodgson 1996). Melting curve PCR (Urdaneta-Marquez et al. 2008) was performed to confirm species in male and female Pulex fleas.
Flea cultures and DNA extraction
Fleas were pooled in 1.5-mL screw-cap vials by species according to nest site. For homogenization of fleas, two glass beads were added to each vial. For vials containing fewer than 10 fleas, we added 100 μL of sterile heart infusion broth (HIB) with 20% glycerol. Flea pools containing 10–200 fleas received 10 μL of HIB per flea (up to 500 μL). Pools containing more than 200 fleas were divided into smaller aliquots and processed accordingly. Using a Mini-BeadBeater, we then homogenized pooled fleas for 1–3 min. A 20-μL aliquot from each freshly homogenized flea pool was then inoculated onto sheep blood agar (SBA). Cultures were incubated at 28°C for 48–72 h and visually examined for presence of gray/white and nonhemolytic colonies morphologically suggestive of Y. pestis.
DNA was extracted from an aliquot of the remaining volume of flea homogenates using the MagNA Pure Compact Nucleic Acid Isolation Kit (Roche Life Sciences, Branford, CT). Sample volumes of 20–200 μL were extracted using the manufacturer-supplied DNA Blood External Lysis protocol. Sample volumes of less than 200 μL were adjusted to that volume with phosphate-buffered saline (PBS) prior to extraction. DNA was eluted in volumes of 100 or 200 μL.
Burrowing owl blood
To assess potential exposure of burrowing owls to Y. pestis, ∼200–300 μL of whole blood was obtained from either or both the adult male and the adult female tending the nests. A 26-guage needle was used to puncture a wing vein to collect blood using microhematocrit tubes. Blood was immediately transferred to 1.5-mL polypropylene centrifuge tubes, which were stored on ice until returned from the field and placed in long-term storage at −20°C. When the birds were about 2–4 weeks of age, 70 μL of whole blood was collected from nestling burrowing owls. Blood from nestlings at a nest was pooled prior to DNA extraction/analysis if there was more than one nestling captured. In 2013, a portion (100 μL) of adult blood samples was added to separate Nobuto filter paper strips for serology, which were allowed to air dry before storing separately in coin envelopes. Occasionally there was sufficient blood from adult owls for serology but not PCR testing, so sample sizes for analyses differed.
For DNA extraction, frozen blood samples were thawed to room temperature and then 10–50 μL of whole blood was added to 300 μL of Queen's Lysis Buffer (0.01 M Tris-Cl, 0.01 M NaCl, 0.01 M EDTA, 1% n-lauroylsarcosine, pH 8.0) and 10 μL Proteinase K. Each sample was allowed to incubate overnight in a water bath at 65°C. After incubation, the mixture was added to 200 μL of lysis binding buffer (EZ BioResearch EZ Blood/Cell DNA Isolation Kit, EZ BioResearch LLC, St. Louis, MO), and genomic DNA was purified according to manufacturer's instructions. Successful DNA extraction and concentration was determined using a Nanodrop ND-2000 UV-VIS spectrophotometer (Wilmington, DE).
Amplification of Y. pestis DNA in flea and owl blood samples
Detection of Y. pestis plasminogen activator (Pla) (Table 1) was performed using nested PCR in 25-μL reaction volumes following an approach similar to Hanson et al. (2007). Negative (no template) and positive (Y. pestis genomic DNA, Idaho Bureau of Laboratories, Boise, ID) controls were amplified and run with experimental samples. Owl1 primers (Table 1) were used to confirm that bird DNA was extracted from the blood. Primers to amplify the Y. pestis low calcium response gene (lcrV) were also constructed (Table 1), but they yielded inconsistent results and, thus, were not used on all samples. Any equivocal results obtained from the pla nested PCR analyses were confirmed with two rounds of PCR using primers specific to the Y. pestis caf and yop genes found on a separate larger plasmid (Table 1). Amplification of all genomic targets was performed under the same conditions, and PCR products were analyzed on 9% or 1.5% agarose gels.
Plague serology on burrowing owl serum
Owl blood samples stored on paper strips (Nobuto) were tested for the presence of antibodies against the Y. pestis–specific F1 capsule antigen using the passive hemagglutination/inhibition assay (PHA/HI) (Chu 2000). A Nobuto titer ≥1:32 is considered positive for evidence of past or current infection. Serological analysis was focused on adult owls to increase the chance of detecting any historical antibody evidence of Y. pestis exposures either on their breeding grounds or from possible recent exposure on wintering grounds/migratory routes; the owls in our study populations were primarily migratory.
Results
Fleas from burrowing owls
From May 29 to June 30, 2012, and April 12 to June 22, 2013, more than 4750 fleas were collected from 86 burrowing owl nest sites located in Idaho, Oregon, Washington, and Colorado (Table 2). The median number of fleas collected per nest site was 36.5 in 2012 (range, 1–244; n=40) and 40.5 in 2013 (range, 1–282; n=46). Although fleas were prevalent on burrowing owls in study sites in Idaho, Oregon, and Washington, just one of 60 adult burrowing owls captured at RMA (Colorado) and none of 55 adult burrowing owls captured at BGNG (South Dakota) harbored fleas (Table 2).
See text for study site locations and abbreviations.
Study site visited only in 2013.
Study site visited only in 2012.
Only owl blood was collected from this study site (See Table 3).
Adult burrowing owls trapped in South Dakota (BGNG, n=55) were devoid of fleas at the time of capture.
More than 99.4% of the fleas we collected from burrowing owls were Pulex spp. (Table 2). Morphological (males) and molecular (males and females) analyses indicated that they were P. irritans, the human flea (Fig. 3). The remaining ∼0.6% of fleas (n=28; Table 2) included individuals that keyed to Meringis, Dactylopsylla, and potentially Oropsylla thrassis. All homogenized flea samples from both 2012 and 2013 yielded bacterial growth on nonselective blood agar media, but none of the cultures contained colonies morphologically indicative of Y. pestis. Therefore, we considered cultures of fleas collected from burrowing owls in both years of study negative for Y. pestis.

Micrographs (10×) of male (
In 2012, there were 46 separate pools of fleas analyzed by PCR for Y. pestis. These included 40 pools of P. irritans from 40 burrowing owls nests located in NCA, DEPOT, YTC, NACHES, and ALE, and six pools of “non-Pulex species” from six nests in NCA, DEPOT, and ALE (Table 2). For 2013, 48 pools of P. irritans from 48 different nests in NCA, DEPOT, BAKER, PASCO, ALE, and RMA were analyzed for Y. pestis, and one pool of “non-Pulex species” from one nest in the NCA was analyzed. Five nest sites in 2013 each contained a single flea that was not P. irritans, and these individuals underwent the clearing procedure for identification that precluded testing for Y. pestis. Thus, 95 flea pools collected at 86 burrowing owls nests in Idaho, Oregon, Washington, and Colorado during 2012–2013 were tested for Y. pestis by PCR. Just as in culturing results, there was no Y. pestis detected in the fleas found on burrowing owls.
Burrowing owl blood
During 2012–2013, blood was obtained from adult burrowing owls (Table 3) in Idaho (NCA), Oregon (DEPOT, BAKER), Washington (PASCO, UNWR, NACHES, ALE), Colorado (RMA), and South Dakota (BGNG). Blood from nestlings (Table 3) was obtained in Idaho (NCA), Oregon (DEPOT), and Washington (NACHES, ALE). While numerous adult burrowing owls were captured in Colorado (RMA) and South Dakota (BGNG) during 2013, there were no nestlings observed at these study sites during trapping. Thus, blood from 170 adult (n=99 females, 71 males) and >400 nestling burrowing owls of unknown sex was obtained. Blood samples/pools (n=239) collected at 143 nests were subjected to PCR testing for Y. pestis (2012: n=5 adult male, 29 adult female, and 48 nestling pools; 2013: n=61 adult male, 69 adult female, and 26 nestling pools). None of the owl blood samples tested positive for Y. pestis.
See text for study site locations and abbreviations.
Blood contributing to pooled sample was from 1 to 11 burrowing owl nestlings per site [mean=6.2±2.7 standard deviations (SD) nestlings per pool].
Study site visited only in 2013.
Study site visited only in 2012.
Serum samples from 128 adult burrowing owls (Table 3) were also collected and analyzed for Y. pestis antibodies. These samples came from owls in Idaho (NCA, n=25 females, 19 males), Oregon (DEPOT, n=11 females, 13 males; BAKER, n=4 females, 6 males), Washington (ALE, n=2 females, 1 male; PASCO, n=5 females, 2 males), Colorado (RMA, n=9 females, 11 males), and South Dakota (BGNG, n=10 females, 8 males). All burrowing owl serum samples tested negative with titers of <1:32.
Discussion
We found that fleas were prevalent on burrowing owls in Oregon, Idaho, and Washington, where we collected more than 4750 fleas from owls at 86 nests during two owl breeding seasons. Examination by traditional culturing techniques and molecular methods (PCR testing) provided no evidence of Y. pestis in these fleas. We also obtained blood from more than 500 burrowing owls at more than 140 nest sites in five states. Molecular testing of owl blood for Y. pestis and serologic examination for Y. pestis antibodies were also consistent with the absence of Y. pestis. Thus, whereas fleas were common and owls inhabited areas with endemic plague, unlike Wheeler et al. (1941), we detected no direct relationship between owl fleas and plague in the areas we studied. It is possible that the low numbers of bacteria present during enzootic cycles are undetectable using these methods and that any such relationships in these areas are more detectable during plague epizootics (Biggins et al. 2010, Matchett et al. 2010), but if and how infection rates would vary during a plague epizootic remains unknown.
The most prevalent species of flea on burrowing owls in Oregon, Washington, and Idaho was P. irritans. Moreover, the only flea collected from a burrowing owl in Colorado was this same species. Smith and Belthoff (2001) found that P. irritans was the most common ectoparasite among lice and fleas previously enumerated from a small number of burrowing owls in Idaho. Although geographic variation in flea abundance and the flea species that burrowing owls harbor throughout their range remain poorly understood, our study extends results about the prevalence of P. irritans on burrowing owls to other states in the region beyond Idaho. In North America, P. irritans is mainly a parasite of large mammals showing a particular preference for carnivores (Lewis et al. 1988). For instance, Harrison et al. (2003) reported that P. irritans was among the most common fleas collected from foxes in New Mexico, where 58.3% of kit foxes (Vulpes macrotis), 73.3% of swift foxes (V. velox), and 11.1% of gray foxes (Orocyon cinereoargenteus) harbored P. irritans. McGee et al. (2006) found that P. irritans was the most prevalent and abundant flea on swift foxes in Rita Blanca National Grassland, Texas, during a plague epizootic. Harris et al. (2014) found P. irritans on black-footed ferrets (Mustela nigripes) in Conata Basin, South Dakota, but in fewer numbers than the most abundant species Oropsylla hirsuta. Finally, Lewis et al. (1988) indicated that the majority of their records of P. irritans from the Pacific Northwest were from mule deer (Odocoileus hemionus columgianus), but indicated occurrence was likely accidental rather than reflecting a true host preference.
In some cases, fleas on burrowing owls may be accidental associates from either the burrow environment or from their prey. For instance, Smith and Belthoff (2001) speculated that the small number of Aetheca wagneri and Meringis hubbardi on burrowing owls in Idaho were accidental associates because these species most commonly parasitize small rodents. Indeed, we also found a small number (<0.6%) of non-Pulex fleas characteristic of small mammals on owls in Idaho, Oregon, and Washington.
That P. irritans occurs on burrowing owls so commonly, in widespread regions, and at times in large numbers, suggests this flea species may have developed a closer relationship with burrowing owls that could include feeding. In fact, recent evidence from molecular blood meal assays has revealed burrowing owl DNA in P. irritans collected from burrowing owls (Graham et al., in prep.). Thus, rather than interacting with owls exclusively as a phoretic host that may move P. irritans fleas potentially infected with Y. pestis from place to place, P. irritans may have potential to bite and infect burrowing owls with Y. pestis. We know that P. irritans can frequently be infected with Y. pestis in portions of its range (Dennis et al. 1999, Ratovonjato et al. 2014), but it may not always be infective because it is not an easily blocked species (Burroughs 1947). However, Eisen et al. (2009) indicate that the importance of common human biting fleas (including P. irritans) as vectors of Y. pestis in places such as Africa may have been underestimated because early-phase transmission by unblocked fleas (cf. Eisen et al. 2006) was not considered. Indeed, Eisen et al. (2009) stress that P. irritans tops the list of additional flea species needing to be evaluated for early-phase transmission of Y. pestis.
In contrast to its habit of infesting carnivores in North America, the cosmopolitan P. irritans commonly infests human dwellings elsewhere. It is considered a potential plague vector in regions of Asia and Africa and was likely a primary vector in Europe during the Black Death (Drancourt et al. 2006). Laudisoit et al. (2007) found that P. irritans is the predominant species among domestic fleas in studies of plague in Tanzania, and it was the only species that inhabited every village and the only species to be significantly higher in high plague frequency villages. In Madagascar, P. irritans was also recently implicated in human-to-human transmission of plague, where it was the most commonly collected flea species, and both engorged and unfed male and female P. irritans carried Y. pestis (Ratovonjato et al. 2014). Because we found no evidence of Y. pestis in the fleas we collected from burrowing owls, if and how effectively P. irritans can carry or transmit Y. pestis to burrowing owls and the response of burrowing owls to such infection remain to be determined. We suggest that future studies similar to ours conducted during plague epizootics in the western United States would clarify the potential relationships we describe or help confirm our negative results.
Footnotes
Acknowledgments
Field and laboratory procedures were approved by Boise State University's Institutional Animal Care and Use Committee (IACUC) Approval #006-AC12-010 and Institutional Biosafety Committee (IBC) Approval #006-IBC12-004. We thank the US Fish and Wildlife Service and the Raptor Research Center at Boise State University for financial and logistical support and M. Schriefer and the Centers for Disease Control and Prevention (CDC), Division of Vector-Borne Disease, Fort Collins, CO, for Nobuto serology. We thank the following for contributions to field and/or laboratory work or help with manuscript figures: C. Alexander, L. Anderson, L. Bristow, A. Bruesch, C. Conway, D. Denlinger, T. Dixon, G. Frye, D. Gillis, J. Giordano, S. Gregory, R. Griebel, M. Hetrick, J. Holderman, J. Hoskins, M. Laskowski, C. Lundblad, J. Mach, H. McLean, K. Meyer, D. Munzing, H. Newsome, E. Pikcilingis, J. Rebholz, M. Stevenson, J. St. Hillaire, J. Wade, and S. Walsh. We thank R. Eisen and C. Graham for input on our study and helpful discussions about plague, and two anonymous referees for numerous comments that helped improve our manuscript. Finally, we thank US Fish and Wildlife Service, US Bureau of Land Management, US Department of Agriculture (USDA) Forest Service, Oregon and Idaho Army National Guard, Umatilla Army Depot, Washington Department of Fish and Wildlife, US Department of Energy Pacific Northwest National Laboratory, and Yakima Training Center for helping to facilitate our field studies.
Author Disclosure Statement
No competing financial interests exist.