
Abstract
Currently, no vaccine has been developed to protect humans from naturally acquired heterologous Orientia tsutsugamushi infections. To enhance the validity of vaccine candidates, we are developing a murine chigger challenge model with the O. tsutsugamushi Lc-1–infected Leptotrombidium chiangraiensis Line-1. To this end, an intraperitoneal (i.p.) murine challenge model using an O. tsutsugamushi Lc-1 isolate was developed for eventual validation of the chigger challenge model. We have determined that the murine lethal dose that kills 50% of the challenged mice (MuLD50) of a liver/spleen homogenate developed from O. tsutsugamushi Lc-1–infected ICR Swiss mice to be 10−6.9. Employing different inoculum doses of this homogenate, the bacterial load using quantitative real-time PCR (qPCR) was determined to range from 60 to 1.6 × 105 genome equivalent copies (GEC)/μL of liver and 33.4 to 2.2 × 105 GEC/μL of spleen tissue. The clinical outcomes relative to homogenate dose levels followed a dose-dependent pattern. The successful development and characterization of the O. tsutsugamushi Lc-1 i.p. challenge model will assist in the development and validation of a mouse chigger challenge scrub typhus model.
Introduction
S
Vaccination would provide a substantial means to control scrub typhus. However, the lack of a successfully developed long-lasting, broadly proactive vaccine after over seven decades of research highlights the difficulty of this endeavor (Chattopadhyay and Richards 2007). Development of a broadly protective scrub typhus vaccine has been hampered by the inability of the host to produce long-term immune protection against the numerous immunogenically disparate Orientia tsutsugamushi isolates in nature and the laboratory (Chattopadhyay and Richards 2007, Hussain et al. 2010). Thus, scrub typhus vaccines developed against one strain have not been effective against other strains of O. tsutsugamushi (Chattopadhyay and Richards 2007).
Another possible shortcoming of scrub typhus vaccine development has been the use of needle-syringe challenge procedures against vaccinated laboratory animals in preclinical studies. Though vaccines showed efficacy in these studies, whether killed, inactivated, live, attenuated, or subunit vaccines, they met with limited success in protecting people exposed to O. tsutsugamushi in natural settings (Chattopadhyay and Richards 2007, Hussain et al. 2010). To evaluate scrub typhus vaccine candidates better, a more natural challenge laboratory animal model needs to be developed, such as a murine chigger challenge model. This model is currently under development by the Department of Entomology, Armed Forces Research Institute of Medical Sciences (AFRIMS).
AFRIMS maintains 12 lines of Leptotrombidium mites, infected with O. tsutsugamushi originally obtained from wild rodents captured in Thailand (Tanskul et al. 1998). The mite lines are derived from three species: Leptotrombidium chiangraiensis, Leptotrombidium deliensis, and Leptotrombidium imphalum. Five of the 12 lines are from L. chiangraiensis collected from areas around homes of patients infected with O. tsutsugamushi in northern Thailand (Tanskul et al. 1998). A recent study investigating the effects of these L. chiangraiensis lines in causing O. tsutsugamushi infection in ICR (CD-1) outbred Swiss mice demonstrated variation in the virulence of the O. tsutsugamushi strains resident in the L. chiangraiensis mite lines (Lurchachaiwong et al. 2012). The L. chiangraiensis Line-1 containing the O. tsutsugamushi Lc-1 isolate proved to infect mice consistently after feeding by a single chigger and was particularly lethal with an average mouse mortality of 93% (Lurchachaiwong et al. 2012). This line is now being used in the development of a chigger challenge model for scrub typhus and will be compared against the current gold standard for scrub typhus, the murine intraperitoneal (i.p.) challenge model (Chattopadhyay and Richards 2007), using O. tsutsugamushi Lc-1.
In this article, we characterize the O. tsutsugamushi Lc-1 isolate and its use in developing a murine i.p. challenge model. From O. tsutsugamushi Lc-1–infected ICR mice, we produced a liver/spleen tissue homogenate for challenge studies, calculated the 50% murine lethal dose (MuLD50) of that homogenate, and evaluated the challenge homogenate by determining the tissue bacterial load and clinical outcome of ICR outbred mice after i.p. inoculation.
Materials and Methods
Scrub typhus i.p. challenge mouse model
The L. chiangraiensis mites were established and maintained at the AFRIMS in the animal biosafety level 3 (ABSL3) laboratories, as previously described (Lurchachaiwong et al. 2012). Female 8-week-old ICR (CD-1) outbred mice, originally derived from Charles River Laboratories (BioLASCO, Taiwan) but maintained at the Department of Veterinary Medicine (DVM), AFRIMS, were used for all experiments. All procedures involving animals were approved by the AFRIMS Institutional Animal Care and Use Committee (IACUC). To determine the MuLD50, we estimated the range of doses that would cover the lowest probability to the highest probability of responses. To ensure negligible bias, the sample size was calculated by using the standard deviation (SD) of the Spearman–Karber estimator. The doses were not spaced more than 2 SD apart. With the use of five animals per dose, we obtained a reliable MuLD50, which had high probability of 95%.
The infected inoculum was prepared as outlined previously (Lurchachaiwong et al. 2014). When the mice showed severe clinical signs (i.e., fur appearance overtly ruffled and activity level reduced to no activity), they were euthanized and their livers and spleens were harvested for inoculum preparation. The pooled liver and spleen tissues were homogenized in Snyder's I buffer (0.2 gram/mL) (Chan et al. 2003). Subsequently, 0.2 mL of a 1:1000 dilution inoculum was used to inoculate by i.p. injection three additional ICR mice as a further adaptation step. A stock inoculum of 0.2 gram/mL tissue homogenate in Snyder's I buffer from these three mice was used in the i.p. challenge experiments.
We used a series of serial dilutions (10−1–10−9) to determine the MuLD50 of our stock inoculum. For each dilution, five mice received a 0.2-mL suspension of inoculum by i.p. injection, whereas the control group of five mice received a 0.2-mL suspension of 10−1 tissue homogenate prepared from uninfected mice. We observed all mice for up to 21 days postinoculation. Clinical illness was assessed using a modification of the procedure of Özkaya et al. (2012). Mice were scored on fur appearance (normal = 2, slightly ruffled = 1, and overtly ruffled = 0) and activity level (normal = 2, reduced activity = 1, no activity = 0), where reduced activity was defined as slow movement after moderate stimulation. On the basis of Özkaya et al. (2012), each mouse received a total score due to its fur appearance and activity level, which indicated the progression of illness from normal “4” to moribund “0”. The total score of “1” was considered a humane end point for severe disease and those animals were euthanized. Mice that died overnight were given a total score of “0”. The rest of the mice that survived to day 21 (end of study) were euthanized.
Detection and quantitation of O. tsutsugamushi
We assessed the bacterial load of O. tsutsugamushi in the liver and spleen tissues of our ICR mice using the Otsu47 quantitative real-time PCR (qPCR) assay (Jiang et al. 2004). Using the QIAamp Mini kit (Qiagen, USA), we extracted O. tsutsugamushi DNA separately from 25 μL of homogenized whole liver and 25 μL of homogenized whole spleen, which were eluted each in 200 μL of kit AE buffer (Qiagen). One-microliter samples of the homogenate DNA extraction preparations were used to determine the genomic equivalent copies (GEC) of infected tissues by the Otsu47 qPCR assay. A plasmid containing the target sequence and the samples were amplified by following a previously reported protocol (Jiang et al. 2004). The GEC number was calculated from the produced standard curve using the Chromo 4 qPCR machine software package per manufacturer's instructions (Bio-Rad, Hercules, CA).
Statistical analysis
The MuLD50 was calculated using a Bayesian model of the mouse dose–response with diffuse, normal, priors for the linear model coefficients (Gelman et al. 2004). A three-chain simulation using the Hit-And-Run Metropolis algorithm (Turchin 1971) was run for an initial burn-in of 106 iterations followed by 15 × 106 iterations with a thinning rate of 5000. Visual inspection of the three chains showed complete mixing and the Gelman–Rubin method (Gelman and Rubin 1992) indicated acceptable convergence for all chains. The parameter values were <1.007. The results from all three simulation chains were used to estimate the posterior LD50 distribution. The Markov chain Monte Carlo (MCMC) results are reported using the median, 2.5%, and 97.5% quantiles (95% confidence interval [CI]). Confidence intervals for clinical illness were calculated assuming a multinomial distribution of probabilities among the five score categories using the method proposed by Sison and Glaz (1995). We fit local regression models to highlight the pattern between inoculum dilution and genomic equivalent copies of O. tsutsugamushi in mouse liver and spleen tissues. These models fit quadratic polynomials in a region defined by the span or amount of the total data set used in the local smoother and assume a normal distribution of the model residual error.
Results
The MuLD50 of the liver/spleen homogenate preparation derived from three ICR mice infected with O. tsutsugamushi Lc-1 by i.p. injection was determined to be 10−6.92 with 95% credible intervals ranging from 10−7.32 to 10−6.48 (Table 1).
MuLD50, 50% murine lethal dose; SD, standard deviation.
Assessment of the bacterial load of both liver and spleen tissues of mice with the 10-fold dilutions of the liver/spleen homogenate used in the MuLD50 determination is shown in Figure 1. The GEC displayed a threshold pattern with a plateau of approximately 10−5 GEC/1.25-μL tissue homogenate at 10-fold dilutions ranging from 10−1 to 10−9 of the homogenate preparations for both tissues and were first detected within the range of dilutions covered by the estimated MuLD50 95% credible intervals (Fig. 1). Median GEC at a dose of 10−7 were 60.5 (range, 0–5.7 × 104) and 33.4 (range, 0–2.8 × 105) per microliter of tissue homogenate, respectively, in liver and spleen tissues. Unfortunately, all the mice at the 10−4 dose and three of the five mice at the 10−5 dose died overnight, resulting in no tissue being harvested for GEC determination.
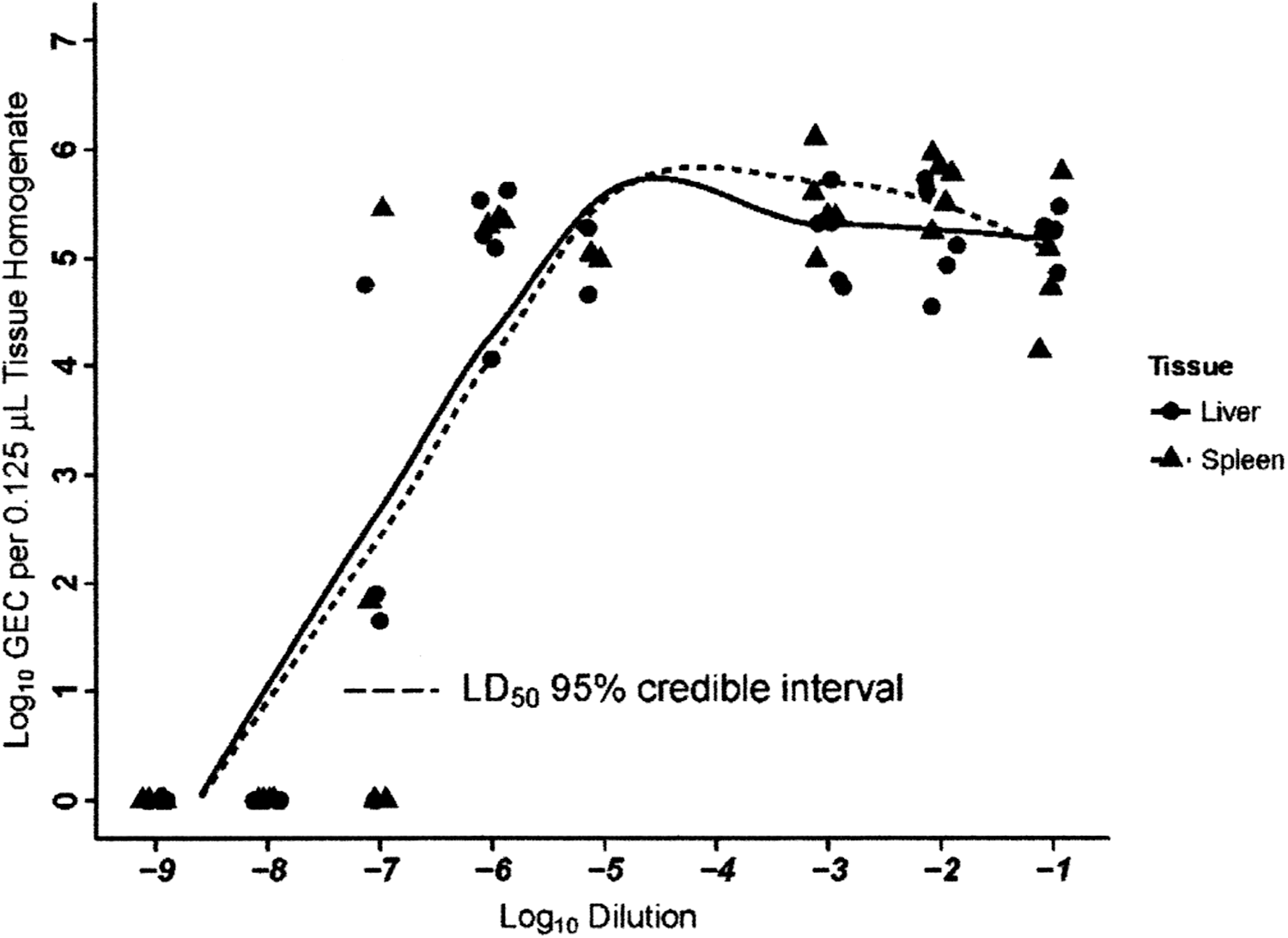
Measurement of genomic equivalent copies of Orientia tsutsugamushi in mouse liver and spleen tissues following increasing doses of O. tsutsugamushi challenge inoculum. The dotted line represents the 50% murine lethal dose (MuLD50) 95% credible interval. Some titer points were moved horizontally to help differentiate overlapping data points. GEC, genome equivalent copies.
The range of clinical outcomes generally followed a dose-dependent pattern (Table 2). Although two mice received a clinical illness score of zero at an inoculum dilution of 10−5, their time to clinical symptoms (TTC) was consistent with the other mice in that group. However, time to mortality (TTM) was considerably shorter at 15 days versus 21 days for the remaining three mice in this group. A plurality (49%) of mice were categorized with a clinical illness score of one, with 68.2% of them receiving inoculum diluted between 10−1 and 10−3. These mice all showed clinical symptoms after 6 days with the median TTM of 8 days (range, 7–9). The remaining mice in this classification included all five mice inoculated at the 10−6 dose and two of the mice dosed at 10−7. However, whereas the latter mice displayed activity and appearance consistent with inoculation at a higher concentration, they took longer to manifest clinical signs of illness (9 days and 12 days, respectively) and consistently lived longer (13 days and 15 days, respectively). All mice dosed at 10−4 and two mice from the 10−5 group were classified with a clinical illness score of 2. The one mouse with a clinical score of 3 occurred at a dilution of 10−7 due to a mildly rough coat appearance. Unless otherwise noted, the remaining mice survived the full 21-day experimental period, with those at the highest dilution never developing signs of clinical illness.
Discussion
Characterizing new challenge models is a critical step in developing and validating their suitability for use in vaccine evaluation. While in the process of developing and characterizing an O. tsutsugamushi Lc-1 chigger challenge model, we have developed and characterized the O. tsutsugamushi Lc-1 i.p. challenge model. The i.p. challenge mouse model is the current standard for evaluating scrub typhus vaccine candidates (Chattopadhyay and Richards 2007, Hussain et al. 2010) and has successfully predicted the success of a vaccine candidate, pKarp47, in providing homologous protection against an intradermal challenge in a nonhuman primate model of scrub typhus (Paris et al. 2015). The O. tsutsugamushi Lc-1 i.p. challenge model was developed using a liver/spleen homogenate from ICR mice fed upon by infected L. chiangraiensis, a mite line derived from a naturally infected mite strain obtained from a wild-caught rodent (Lurchachaiwong et al. 2012, 2014). With both the i.p. challenge and chigger challenge models infected with the same O. tsutsugamushi Lc-1 strain, we will be able to compare data between chigger challenge and i.p. challenge models in assessing vaccine candidate efficacy, which will provide us the bridging information needed to validate the natural chigger challenge model. A validated chigger challenge model will allow us the ability to assess more appropriately the efficacy of prophylactic and therapeutic treatments.
The reason for selecting L. chiangraiensis mites infected with O. tsutsugamushi Lc-1 for the development of a natural challenge model is that these mites, originally collected from the northern part of Thailand (Tanskul et al. 1998), have been consistently shown to have vertical and horizontal transmission rates of O. tsutsugamushi highest among the three species of Leptotrombidium assessed (Lerdthusnee et al. 2002). Moreover, the L. chiangraiensis Line-1 demonstrated the most consistent infection of mice with O. tsutsugamushi following the feeding of single infected chigger, and the O. tsutsugamushi isolate Lc-1 showed a very high virulence for the ICR mice, with an average mortality rate of 93% (Lurchachaiwong et al. 2012). Thus, it was determined that L. chiangraiensis Line-1 would be used to develop a chigger challenge model for evaluation of the efficacy of scrub typhus vaccine candidates and/or therapeutic agents.
Lethality in ICR mice is an important component in determining the suitability of an O. tsutsugamushi isolate as a candidate for vaccine challenge studies (Chan et al. 2003). In this study, we established a MuLD50 for the Lc-1 isolate of O. tsutsugamushi, investigated the bacterial loads in selected tissues of mice, and assessed the affects of relative doses on clinical presentation. The MuLD50 of the O. tsutsugamushi Lc-1 liver/spleen homogenate was determined to be 10−6.91, which falls within the range of those estimated by Chan et al. (2003), for eight strains of virulent O. tsutsugamushi in CD-1 mice. All the ICR mice died when challenged with 1000× MuLD50 (10−3.91), a dose used to evaluate the effectiveness of vaccine candidates in laboratory animals when prevention of death is used as a metric of efficacy (Chattopadhyay and Richards 2007).
Bacterial loads of O. tsutsugamushi in murine tissues are associated with strain lethality (Valbuena and Walker 2013). In a recent clinical study, Sonthayanon et al. (2009) found that O. tsutsugamushi DNA loads were two orders of magnitude higher in fatal cases compared to survivors, but clinical illness occurred at much lower levels of bacterial loads than had been previously reported. GEC in our ICR mice rose quickly as doses increased from 10−7 to 10−6, with the median GEC increasing from 60 copies per microliter of homogenate to 1.6 × 105 copies in liver tissue and 33.4 copies to 2.2 × 105 in spleen tissue. However, the range of GEC found in liver and spleen tissues of mice inoculated with the lower doses of homogenate seeds was quite wide and was associated with clinical outcomes of the individual mice.
The onset of clinical symptoms and survival time in this study at inoculum dilutions of 10−6–10−7 also approximate those reported previously for the O. tsutsugamushi Lc-1 strain administered by needle challenge (Lurchachaiwong et al. 2014). Disease onset in two mice inoculated with a 10−7 dose of O. tsutsugamushi Lc-1 liver/spleen homogenate with clinical outcome scores of one was the same as three other mice similarly inoculated with clinical outcome scores of three and four, but death occurred after 15 days for the two mice with clinical outcome scores of one versus 21 days for the three with clinical outcome scores of three and four. These latter three mice and other healthier mice (inoculated with lower doses) had a maximum GEC, regardless of tissue type, of only 78.2 copies compared to 5.6 × 104 copies in the two sicker mice given a 10−7 dose or the minimum of 5.9 × 103 copies in mice given a 10−6 dose. These results suggest that once bacterial dissemination begins, very little of the pathogen is required to induce clinical signs and symptoms whereas larger doses are required for lethality.
The genetic diversity inherent in outbred ICR mice may increase the breadth of immune responses at the same injection dose (Nikodemova and Watters 2011). Lerdthusnee et al. (2006), measuring immunoglobulin M (IgM) and IgG, showed IgM production in 10-week-old ICR mouse infected with O. tsutsugamushi through chigger feeding beginning 12 days postinfection. All of our mice at the 10−7 dose showed symptoms 12 days postinoculation, yet only two mice died before 21 days, suggesting differences inherent to the outbred ICR mice may impact the outcome at intermediate challenge dose levels. Alternatively, while thorough mixing of the inoculum at each dose should reduce heterogeneity in bacterial loads across mice, the possibility of higher bacterial concentrations in individual injections cannot be ruled out.
Intraperitoneal inoculation is the current method used to challenge mice with O. tsutsugamushi for scrub typhus vaccine candidate evaluations (Chattopadhyay and Richards 2007, Hussain et al. 2010). While recognizing the shortcomings of this unnatural technique, we are moving toward the development of a validated murine chigger challenge model. Our intent is to use a well-characterized i.p. challenge model to validate the chigger challenge model and then adopt the direct chigger feeding challenge model for future scrub typhus vaccine evaluations.
Footnotes
Acknowledgments
We thank Dr. Kriangkrai Lerdthusnee, Dr. Wei-Mei Ching, and all staff at the department of Entomology, AFRIMS, Bangkok, Thailand, for assistance and kind support. All procedures involving animals were approved by the AFRIMS Institutional Animal Care and Use Committee (IACUC) (protocol no. 08-04).
Author Disclosure Statement
The opinions or assertions contained herein are the private views of the authors, and are not to be construed as official, or as reflecting the views of the Department of the Army, Department of the Navy, or the Department of Defense. Research was conducted in compliance with the Animal Welfare Act and other federal statutes and regulations relating to animals and experiments involving animals and adheres to principles stated in the Guide for the Care and Use of Laboratory Animals, NRC Publication, 2011 edition. As employees of the US Government (A.L.S. and A.L.R.), this work was prepared as part of their official duties and therefore under Title 17 USC paragraph 105 copyright protection is not available. No competing financial interests exist.