
Abstract
Trypanosoma rangeli is a nonpathogenic parasite for humans; however, its medical importance relies in its similarity and overlapping distribution with Trypanosoma cruzi, causal agent of Chagas disease in the Americas. The genetic diversity of T. rangeli and its association with host species (triatomines and mammals) has been identified along Central and the South America; however, it has not included data of isolates from Ecuador. This study reports infection with T. rangeli in 18 genera of mammal hosts and five species of triatomines in three environments (domestic, peridomestic, and sylvatic). Higher infection rates were found in the sylvatic environment, in close association with Rhodnius ecuadoriensis. The results of this study extend the range of hosts infected with this parasite and the geographic range of the T. rangeli genotype KP1(−)/lineage C in South America. It was not possible to detect variation on T. rangeli from the central coastal region and southern Ecuador with the analysis of the small subunit ribosomal RNA (SSU-rRNA) gene, even though these areas are ecologically different and a phenotypic subdivision of R. ecuadoriensis has been found. R. ecuadoriensis is considered one of the most important vectors for Chagas disease transmission in Ecuador due to its wide distribution and adaptability to diverse environments. An extensive knowledge of the trypanosomes circulating in this species of triatomine, and associated mammal hosts, is important for delineating transmission dynamics and preventive measures in the endemic areas of Ecuador and Northern Peru.
Introduction
T
T. rangeli is mainly transmitted by salivary inoculation, although oral transmission by ingestion of triatomines has also been proposed as an important epidemiological route of infection with trypanosomes (T. rangeli and T. cruzi) of mammalian hosts such as dogs (Montenegro et al. 2002, Pineda et al. 2011) and other mammals with grooming behavior. Distribution of T. rangeli is related to the distribution of the triatomines; however, it depends on the capacity of triatomines to harbor the infective stage of the parasite in the salivary glands (Guhl and Vallejo 2003). Most species of triatomines can transmit T. cruzi, but the transmission of T. rangeli has been more restricted to the triatomines of the genus Rhodnius (D'Alessandro and de Hincapie 1986, Vallejo et al. 2009). Experimental infection has been conducted in Triatoma, Panstrongylus, and Rhodnius, but only Rhodnius has demonstrated the presence of the infective forms of T. rangeli in the salivary glands (De Stefani Marquez et al. 2006). Natural or experimental infection has been demonstrated in 12 out of the 15 species of Rhodnius (Guhl and Vallejo 2003), although natural infection of the salivary glands has also been reported in the genus Triatoma (Marinkelle 1968).
Biochemical, immunological, and molecular characteristics have identified polymorphisms in T. rangeli strains isolated from different hosts (Grisard et al. 1999). However, the close association with the genus Rhodnius has defined the distribution of the different genotypes (Machado et al. 2001, Urrea et al. 2011). Two main groups (KP1[+] and KP1[−]) have been defined based on the presence/absence of the KP1 minicircle in the kinetoplast DNA (kDNA) (Vallejo et al. 2002). Additionally, analysis of the small subunit of ribosomal RNA (SSU-rRNA) has also identified the presence of five lineages (A, B, C, D, and E) in Central and South America (Maia Da Silva et al. 2007, 2009). Further analysis of the splice leader intergenic region (SL) confirmed that the different genotypes of T. rangeli circulate in association with vectors of the same evolutionary line, independently of the geographic distance (Urrea et al. 2011). Most recently, subdivision in KP1(−) has been detected by microsatellite typing (Sincero et al. 2015). Moreover, with the publication of the genome of T. rangeli (Stoco et al. 2014), the complexity of this parasite could be further addressed.
Multiple studies have evaluated the genetic diversity of T. rangeli along Central and South America. However, none of those have included samples from Ecuador. Even though the presence of T. rangeli has been reported previously in triatomines and mammals from central coastal and southern Ecuador (Pinto et al. 2006, Grijalva et al. 2011, 2012, Villacis et al. 2015), a detailed picture of its distribution and prevalence in these areas has not been assessed. The aim of this study was to contribute information about the diversity of the T. rangeli populations that are circulating in mammals and triatomines in the southern province of Loja and the central coastal province of Manabí, both considered as endemic areas for Chagas disease (Black et al. 2007, 2009).
Materials and Methods
This study was carried out with DNA samples previously collected as part of the Chagas disease projects of the Center for Infectious and Chronic Disease Research (CIDCR) from 2004 to 2012. Samples of this study were collected in 50 rural communities (19 counties) from central coastal Ecuador (Manabí Province) and 53 rural communities (11 counties) from southern Ecuador (Loja Province) (Table 1). DNA was obtained from intestinal contents (Grijalva et al. 2012) of triatomines and blood samples from mammals (Pinto et al. 2006). An aliquot of each sample was inoculated in culture for in vitro trypanosomes isolation (Chiari et al. 1989, Yeo et al. 2007), and DNA was isolated from cultures that presented growth of parasites. All samples were obtained in strict accordance with the protocol approved by the Ohio University Institutional Animal Care and Use Committee (IACUC).
Includes species from the genera Artibeus, Desmodus, and Glossophaga.
Includes species from the genera Didelphis, Marmosa, and Micoureus.
Includes species from the genera Aegialomys, Akodon, Cavia, Mus, Rattus, Rhipidomys, and Sciurus.
Includes species from the genera Didelphis, Marmosa, and Philander.
Includes species from the genera Handleyomys, Hoplomys, Mus, Proechimys, Rattus, Rhipidomys, Sciurus, and Transandinomys.
Differential PCR for T. rangeli identification
Initial screening for detection of natural infection with T. rangeli was carried out on 3606 DNA samples obtained from the intestinal contents of triatomines and blood from mammals (Table 1). Samples were amplified for the conserved domain of the minicircle of kDNA, as previously described by Vallejo et al. (1999) for the samples from 2004–2010 and Virreira et al. (2006) for the samples from 2011–2012. Differential detection between T. cruzi and T. rangeli was determined based on the size of the PCR products. A band of 330 bp was expected for T. cruzi, whereas a band of 760 bp together with bands of 300–450 bp defined T. rangeli. Confirmation of T. rangeli infection was carried out with further amplification of the D7a divergent region of the large subunit rRNA gene, as previously described (Souto et al. 1999). A band of 210 bp was expected for T. rangeli, whereas T. cruzi presented a band of 265 bp. A negative control (without DNA) and control DNA for T. cruzi and T. rangeli were used to validate the gels. Bands were detected in a 2% agarose gel.
Selection of T. rangeli samples for genotyping
Genotyping was carried out in a subset of 111 samples. From them, 68 were genotyped from the DNA obtained from intestinal contents/blood samples and 43 from DNA obtained from T. rangeli isolates in culture. This subset is representative of the geographic origin of the samples and includes a diversity of hosts (seven species of triatomines, and four species of mammals) (Table 2). The geographic area included 12 and 27 rural communities in Loja and Manabí Province, respectively.
N.D., not determined.
Identification of T. rangeli groups (KP1[+] and KP1[−]) by amplification of the conserved regions of the minicircles of kDNA
The conserved regions of the minicircle of kDNA were amplified with the primers S35 (5′-AAA TAA TGT ACG GGT GGA GAT GCA TGA-3′), S36 (5′-GGG TTC GAT TGG GGT TGG TGT-3′), and KP1L (5′-ATA CAA CAC TCT CTA TAT CAG G-3′), as previously described (Vallejo et al. 2002). A band at 760 bp and fragments between 300 and 450 bp were expected in all samples. KP1(+) was identified by the presence of an additional band at 165 bp, whereas KP1(−) was identified by the absence of this fragment. Additionally, samples that could not be classified by the previous markers were amplified for the specific variable region KP1 using the primers KP1U (5′-GTA GAA AGA TCC GAA AAA ATG C-3′) and KP1L (5′-ATA CAA CAC TCT CTA TAT CAG G-3′) (Vallejo et al. 1994). A band of 280 bp was visible in KP1(+) group and was absent in KP1(−). A negative control and positive DNA for KP1(+) and KP1(−) were used to validate the gels. Bands for both molecular markers were detected in a 6% polyacrylamide gel.
Identification of T. rangeli lineages by the intergenic-spacer of the SL gene
T. rangeli lineages were identified by differential amplification of the SL intergenic spacer using the primers TraSL1 (5′-GAA CGG TCG TGT TCT G-3′) and TraSL2 (5′-GAC GGG ATG TGG TGC-3′), as previously described (Maia Da Silva et al. 2007). The expected band sizes for the different lineages are: lineage A (417 bp), lineage B (380 bp), lineage C (480 bp), lineage D (500 bp), and lineage E (987 bp) (Maia Da Silva et al. 2007, 2009).
Sequencing of the SSU-rRNA (18S rRNA) gene
From the samples that were able to be characterized by both the conserved region of the minicircle of kDNA and the intergenic-spacer of the SL gene, a representative subset of samples (n = 17), which represents the geographic and host diversity, were selected for sequencing of a fragment of the SSU-rRNA gene (Table 3). PCR amplifications were conducted with the newly designed primers SSU4_F (5′-GTG CCA GCA CCC GCG GTA AT-3′) and 18Sq1R (5′-CCA CCG ACC AAA AGC GGC CA-3′), using a touchdown PCR profile (Murphy and O'Brien 2007). The PCR products were cleaned with ExoSAP-IT (Affymetrix, Santa Clara, CA), and sequencing reactions for both ends were performed with the ABI BigDye chemistry (Applied Biosystems, Inc., Foster City, CA) for the external primers mentioned above and the internal primers SSU561F (5′-TGG GAT AAC AAA GGA GCA-3′) and SSU561R (5′-CTG AGA CTG TAA CCT CAA AGC-3′) (Noyes et al. 1996, 1999).
Samples in bold indicate the sequences newly generated for this study.
Sequencing of DNA fragments was conducted on an ABI 3730xl DNA Analyzer automatic sequencer (Applied Biosystems, Inc., Foster City, CA). Contigs for each trypanosome sample were assembled using Geneious version 8.0.3 (Biomatters 2014) with untrimmed sequences with a percentage of high quality bases (HQ%) higher than 50%, resulting in trimmed sequences with a HQ% higher than 90%; consensus sequences were extracted from each contig. The sequences obtained were aligned with T. rangeli representatives of lineages A to E (Maia Da Silva et al. 2007, 2009) and selected species of the T. cruzi clade as outgroups (Table 3) using the MUSCLE (Edgar 2004) plugin in Geneious. The alignment was checked manually for misplacements and trimmed to 816 bp to ensure completeness of 100% of the sequence length of representatives of all T. rangeli lineages. A phylogenetic analysis using maximum likelihood was performed in RAxML v. 8 (Stamatakis 2014) using the GTRGAMMA model and 1000 bootstrap replicates.
Results
Diversity and distribution of hosts infected by T. rangeli
A total of 3606 samples from triatomines and mammals were screened for the presence of trypanosomatids. Of those, 351 (10%) presented infection for T. rangeli, and from them, 1.25% corresponded to mixed infections with T. cruzi. A higher infection rate was found in Manabí Province than in Loja Province, 14% and 7%, respectively (Table 1) (Fig. 1A, B). T. rangeli had a wide distribution in both regions and was present in more than 60% of the counties included in this study. Along this geographic distribution, mixed infections with T. cruzi were also reported (Fig. 2).

Differential detection of T. cruzi and T. rangeli infection of triatomine's intestinal contents by (
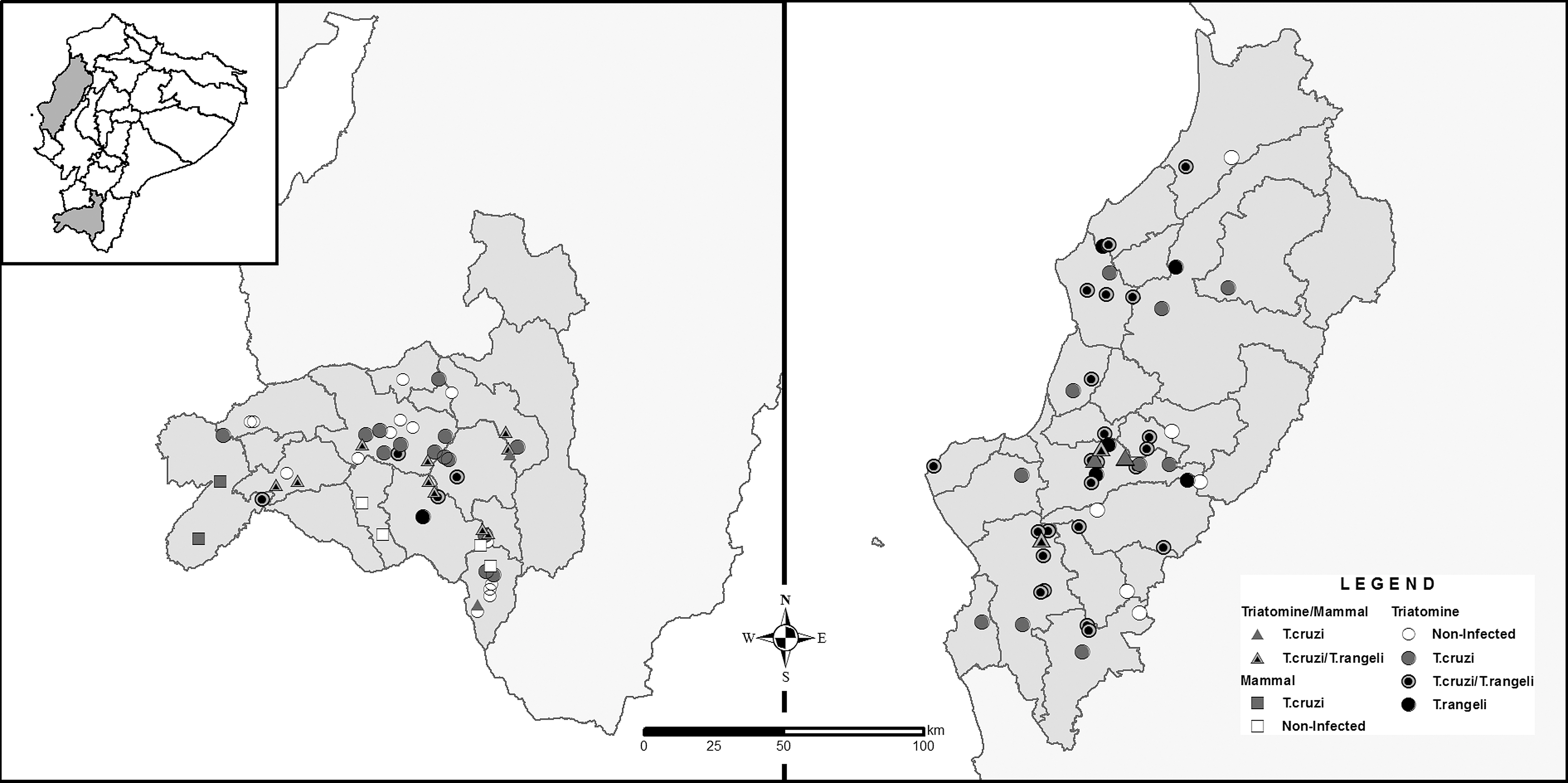
Geographic distribution of T. rangeli and T. cruzi infection in invertebrate (triatomines) and vertebrate (mammal) hosts in southern Ecuador, Loja Province (left), and central coastal Ecuador, Manabí Province (right). Lines indicate county division.
Regarding the mammalian hosts, 18 genera from the order Chiroptera (Artibeus, Desmodus, Glossophaga), Didelphimorphia (Didelphis, Marmosa, Micoureus, Philander), and Rodentia (Aegialomys, Akodon, Cavia, Mus, Rattus, Rhipidomys, Sciurus, Handleyomys, Hoplomys, Proechimys, Transandinomys) were found to be infected with T. rangeli. However, infection rates were <6% (Table 1). With respect to the triatomine hosts, five species were found infected: Panstrongylus chinai, P. rufotuberculatus, P. howardi, Triatoma carrioni, and Rhodnius ecuadoriensis. The species R. ecuadoriensis showed the highest infection rates in both regions (10% and 17%), whereas the other species presented infection rates <5% (Table 1).
T. rangeli was found circulating in all the three habitats (domicile, peridomicile, and sylvatic). Nevertheless, a notably higher infection rate was found in the sylvatic environment (17%) compared with the peridomicile and domestic environments, which presented 7% and 2%, respectively (Table 1).
Genotyping of T. rangeli circulating in vertebrate and invertebrate host
From the 351 samples that were determined to be positive for T. rangeli infection, a representative subset of 111 was characterized by the conserved region of the minicircle of kDNA and the intergenic-spacer of the SL gene. Ninety-nine percent of the samples were classified as KP1(−), independent from the geographic or host origin (Fig. 1C). It was not possible to genotype one sample from R. ecuadoriensis due to the presence of nonspecific bands (Table 2). On the other hand, 80% (n = 89) of the samples (including triatomines and mammals) were determined as lineage C (Fig. 1D), while 20% (n = 22) did not amplify any of the expected bands or amplified bands of not expected size (Table 2).
Sequencing of the SSU-rRNA (18S rRNA) gene
Of the 89 samples that were able to be characterized as KP1(−)/lineage C, a subset of 17 samples were sequenced. Within this subset, we included samples that could not be characterized by PCR of the kDNA (TCQ991) or SL (MBJ1411, TCE694). All samples in this study were identical, and the phylogenetic analysis grouped the sequences with lineage C of T. rangeli (Fig. 3). In the phylogenetic tree, lineage E does not differentiate from lineage A, and there is little differentiation between lineages D and A (Fig. 3).

Maximum likelihood tree of isolates of T. rangeli with representatives of five lineages (
Discussion
Manabí and Loja Provinces are considered as endemic areas for Chagas disease in Ecuador, and active transmission due to the presence of the parasite T. cruzi and its hosts (triatomines vectors and mammal hosts) has been confirmed (Grijalva et al. 2003, 2005, Pinto et al. 2006, Black et al. 2009). Moreover, the presence of the nonpathogenic T. rangeli has also been reported as overlapping T. cruzi distribution (Pinto et al. 2006, Grijalva et al. 2011, 2012, Villacis et al. 2015). This study confirmed the presence of both species of trypanosomes in 1.25% of the samples. However, it is important to consider that the infection rate reported may be underestimated due to the fact that screening of T. rangeli was carried out in intestinal contents and not in salivary glands or hemolymph. Nevertheless, a broad delineation of the distribution and genetic diversity of T. rangeli circulating in these areas has not been reported until now.
Infection of T. rangeli has been detected in mammals of at least five orders (Didelphimorphia, Chiroptera, Rodentia, Pilosa, and Primates) (Ramirez et al. 2002, Maia da Silva et al. 2004a, b, Cabrine-Santos et al. 2009, Maia da Silva et al. 2009, Ramirez et al. 2014). This study confirmed the presence of T. rangeli in three mammalian orders: Didelphimorphia, Chiroptera, and Rodentia, and in 18 genera, thus extending the report of host diversity for this parasite (Maia da Silva et al. 2004a, b, Pinto et al. 2006, Gurgel-Goncalves et al. 2012). However, infection rates reported in this study are low compared with others previously reported in Didelphimorphia and Chiroptera (Ramirez et al. 2002, Ramirez et al. 2014).
Extensive research of the vector–parasite relationships between T. rangeli and triatomine insects supports that the distribution of T. rangeli is closely related to the distribution of triatomines of the genus Rhodnius (Gaunt and Miles 2000, Machado et al. 2001, Urrea et al. 2005, Vallejo et al. 2007, Pulido et al. 2008, Salazar-Anton et al. 2009, Vallejo et al. 2009, Garcia et al. 2012). Even though the species of the genus Rhodnius are particularly susceptible to infection of salivary glands by T. rangeli, such infection has been also reported in Triatoma dimidiata (Marinkelle 1968, D'Alessandro-Bacigalupo and Gore Saravia 1991). However, the vector capacity due to the presence of infective stages of T. rangeli has only been proven in Rhodnius (Guhl and Vallejo 2003). While other triatomines species of the genera Triatoma and Panstrongylus have also reported to be naturally infected with T. rangeli, these reports are restricted to infection in the intestines (D'Alessandro-Bacigalupo and Gore Saravia 1991, Vallejo et al. 2002, Villacis et al. 2015). This study extends the species of triatomines naturally infected with T. rangeli to P. chinai, P. rufotuberculatus, and T. carrioni. Although, finding other species than Rhodnius infected with this parasite is feasible, our results do not imply the capacity of transmission of these species because the samples came from intestinal contents isolates and not from salivary glands.
R. ecuadoriensis was the species of triatomine with the highest infection rates. Although this study reports T. rangeli infecting intestinal contents, the natural and experimental infection of salivary glands has been previously reported in this species of triatomine (Cuba 1975, Guhl and Vallejo 2003, Urrea et al. 2005). However, its capacity to transmit T. rangeli has not been evaluated yet. Recent studies have demonstrated that R. ecuadoriensis is an important threat for Chagas disease transmission due to its wide geographical distribution, high infection rates with T. cruzi, good vector efficiency, and capacity of adaptation to different habitats (Villacis et al. 2008, Grijalva and Villacis 2009, Suarez-Davalos et al. 2010, Grijalva et al. 2012, 2014). In this regard, the presence of R. ecuadoriensis, infected with T. rangeli, constitutes an important element to consider when the distribution of trypanosomes is evaluated in Chagas disease endemic areas. Although the infection rates in the domestic and peridomestic environments were low (<7%), the high infection rates of T. rangeli in R. ecuadoriensis from sylvatic environments (17%) and the proven synanthropic tendency of the sylvatic populations of this species (Grijalva and Villacis 2009, Suarez-Davalos et al. 2010) indicate a risk for an increase of T. rangeli in domestic/peridomestic environments with its well-known consequences of being a confounding factor for Chagas disease diagnosis (Saldana and Sousa 1996a, b). The characteristics of R. ecuadoriensis and its important percentage of infection with T. rangeli, together with the morphological and genetic similarities that it shares with the pathogenic T. cruzi, need to be addressed when the biological and epidemiological scenarios are assessed, especially considering that infections of T. rangeli in humans have been previously reported in South America (Guhl and Vallejo 2003, Saldana et al. 2005).
Only T. rangeli KP1(−)/lineage C was found, even though the samples came from a variety of host species (triatomines and mammals) and two geographical regions that are ecologically different. This study complements the geographic range for KP1(−) in the Andean region, which has been previously described in Colombia and Peru (Vallejo et al. 2009, Urrea et al. 2011) and also for lineage C, which has been previously reported in Central America and Colombia (Maia da Silva et al. 2004a, b). It was not possible to identify the lineage of 22 out of 111 samples with the SL gene. However, two of these samples (MBJ1411 and TCE694) were included in the subset for sequencing, and they were characterized as lineage C. We relate this gap of information with the source of DNA. These samples were obtained directly from the hosts (intestinal contents or blood), and the amount of T. rangeli DNA was difficult to assess. Although some previous attempts have been carried out to detect Trypanosoma from field samples, especially T. cruzi (Hamano et al. 2001, Marcet et al. 2006), all of them rely on the highly repetitive regions as the kDNA.
The distribution of the different genotypes of T. rangeli is affected by the trypanolytic capability of the species of Rhodnius (Pulido et al. 2008, Vallejo et al. 2009). Indeed, T. rangeli distribution is not associated with the vertebrate host but with the biogeographical distribution of Rhodnius species, even in areas where sympatric Rhodnius species coexist (Machado et al. 2001, Salazar-Anton et al. 2009, Urrea et al. 2011). The presence of the genotype T. rangeli KP1(−) associated with R. ecuadoriensis is in total accordance with what has been previously described for this species of triatomine (Urrea et al. 2005, 2011) and with what is expected regarding lineage C (Vallejo et al. 2009). Thus, the predominantly presence of R. ecuadoriensis in the study area also explains the presence of T. rangeli KP1(−)/lineage C in the samples from mammal hosts.
A phenotypic analysis of antennas and wings from R. ecuadoriensis demonstrated differences between the triatomine population of Loja and Manabí that suggest a speciation process (Villacis et al. 2010), especially considering that both geographic areas are separated by the Andes mountains, which constitute a natural barrier. An effect of triatomine subdivision was not evident in the T. rangeli samples of this study with the SSU-rRNA gene. However, detection of population structure among the T. rangeli KP1 (−/lineage C isolates with other molecular markers is possible, especially in light of the recent subdivision found by microsatellite typing on T. rangeli KP1(−) that was related to geographical location or co-evolution of parasites and hosts (Sincero et al. 2015). The access to the recently published complete genome of T. rangeli (Stoco et al. 2014) will provide new tools for elucidating the complex biology of this parasite and its interaction with its hosts.
Conclusions
This work reports the natural infection by T. rangeli in species of three mammalian orders (Didelphimorphia, Chiroptera, Rodentia) and four triatomine insect species (P. chinai, P. rufotuberculatus, R. ecuadoriensis, and T. carrioni) in southern (Loja) and central coastal (Manabí) Ecuador. Despite its presence in all the three habitats (domestic, peridomestic, and sylvatic), a higher infection rate was found in the sylvatic habitat and related to the presence of R. ecuadoriensis. A representative subset of T. rangeli that includes samples from a wide geographic area and a variety of host species (triatomines and mammals) was classified as KP1(−) and to the lineage C. No diversity was found among the SSU-rRNA gene, even though the samples came from ecologically different areas where subdivision of R. ecuadoriensis populations has been detected by phenotypic analysis.
Footnotes
Acknowledgments
Financial support was received from the Global Infectious Diseases Training Grant–Fogarty International Center–National Institutes of Health (D43TW008261); the Division of Microbiology and Infectious Diseases, National Institute of Allergy and Infectious Diseases, National Institutes of Health (DMID/NIAID/NIH (1 R15 AI077896-01); the United Nations Children's Fund (UNICEF)–United Nations Development Programme (UNDP)/World Bank/World Health Organization Special Programme for Research and Training in Tropical Diseases Research Capability Strengthening Group (grant ID A20785 and A60655); the European Union Seventh Framework Programme, contract number 223034 (ChagasEpiNet); Ohio University Heritage College of Osteopathic Medicine; PEW Latin American Fellows Program in the Biomedical Sciences (ID:00026195); and Pontifical Catholic University of Ecuador (I13048).
Special thanks are directed to the personnel of the National Chagas Control Program–National Vector Borne–Disease Control Service, Ecuadorian Ministry of Health; to Francisco J. Ayala for the critical review of the manuscript; and to María Augusta Chávez and Freddy Proaño from the University of the Army Force–ESPE for their academic assistance. Technical assistance was provided by Anita Villacís, César Yumiseva, Fernanda Latorre, and Andrea Zurita from the Center of Infectious Disease Research at Pontifical Catholic University and Uriel Alvarado and Daniel Zabala from the Laboratorio de de Investigaciones en Parasitología Tropical at Universidad de Tolima.
Author Disclosure Statement
No competing financial interests exist.