
Abstract
Dengue is one of the most important diseases caused by arboviruses in the world. Yellow fever is another arthropod-borne disease of great importance to public health that is endemic to tropical regions of Africa and the Americas. Both yellow fever and dengue viruses are flaviviruses transmitted by Aedes aegypti mosquitoes, and then, it is reasonable to consider that in a given moment, mosquito cells could be coinfected by both viruses. Therefore, we decided to evaluate if sequential infections of dengue and yellow fever viruses (and vice-versa) in mosquito cells could affect the virus replication patterns. Using immunofluorescence and real-time PCR-based replication assays in Aedes albopictus C6/36 cells with single or sequential infections with both viruses, we demonstrated the occurrence of viral interference, also called superinfection exclusion, between these two viruses. Our results show that this interference pattern is particularly evident when cells were first infected with dengue virus and subsequently with yellow fever virus (YFV). Reduction in dengue virus replication, although to a lower extent, was also observed when C6/36 cells were initially infected with YFV followed by dengue virus infection. Although the importance that these findings have on nature is unknown, this study provides evidence, at the cellular level, of the occurrence of replication interference between dengue and yellow fever viruses and raises the question if superinfection exclusion could be a possible explanation, at least partially, for the reported lack of urban yellow fever occurrence in regions where a high level of dengue transmission occurs.
Introduction
Y
In Brazil, the intense vaccination against the YFV in endemic regions and especially the A. aegypti eradication program significantly contributed to an effective prevention of urban YFV epidemics and control of dengue epidemics until the 80s (Figueiredo 2000, 2007, Vasconcelos 2003). However, in the last 50 years, there has been a dramatic growth of urban centers in the tropical Americas, mostly in a chaotic way, paving the grounds for the reinfestation of the main urban vector for DENV and YFV, the A. aegypti mosquito (Gubler 2002). This epidemiologic situation has resulted in an increased number of reported dengue cases in Brazil but not in the reemergence of urban yellow fever (Codeço 2004) despite the fact that since 2001, an increased number of the sylvatic yellow fever has been reported in Brazil (Vasconcelos 2001, 2003, Figueiredo 2007).
The high yellow fever vaccine coverage in Brazil and the fact that different vectors are responsible for urban and sylvatic transmission could be responsible for these epidemiological observations. However, other factors may be involved in the lack of simultaneous transmission of dengue and yellow fever viruses in Brazil. To contribute to the further understanding of these epidemiological findings, we hypothesized that a considerable percentage of A. aegypti in the cities undergoing dengue outbreaks could be infected with DENV and they would not support subsequent infection with YFV. Thus, we decided to investigate if such subsequent infections could interfere with the replication of these viruses. This phenomenon, known as viral interference or superinfection exclusion (homologous interference), refers to the inability of a virus to replicate in an organism or a cell that has been previously infected by another closely related virus (Johnston 1974, Blitvich and Firth 2015). This phenomenon is different from dual infections, a situation where either cell lines or even humans can be infected simultaneously by two viruses (Leroy et al. 2009). To address superinfection exclusion between DENV-2 and YFV, we have performed molecular and morphological assays in A. albopictus cells, which were exposed either to a single infection by each one these viruses or to a sequential infection with both of them. Our data show that YFV replication is severely impaired in cells that had been previously infected with DENV-2, and that DENV-2 replication is also impaired in C6/36 cells previously infected with YFV, although to a lower extent.
Materials and Methods
Virus strains
Viral stocks of DENV-2 (New Guinea C strain), YFV-17D, and YFV-BeH111 strains were prepared in C6/36 cells and suckling mouse brains and used throughout the study. C6/36 cells were kept at 28°C and maintained in the Leibovitz L-15 medium containing 10% heat-inactivated fetal bovine serum (L15-10% FBS), 10% tryptose phosphate broth, 150 U/mL penicillin, and 100 mg/mL streptomycin. C6/36 cells were infected with each virus during 1 h at room temperature. Then, a fresh maintenance medium (2% FBS) was added to the cell cultures and incubated at 28°C. Specific infection by each virus was detected by immunofluorescence assays (IFA) after 7 days postinfection using hyperimmune mouse ascitic fluids prepared against DENV-2 (Tesh 1979) and rabbit immune serum against YFV-17D. Supernatants from virus-infected C6/36 cells were harvested, aliquoted in the presence of L15-2% FBS, and stored at −80°C for later use.
Plaque assay
Virus titers were determined by plaque assays in Vero cell monolayers grown in 24-well plates. DENV-2, YFV-BeH111, and YFV-17D viral stocks were serially 10-fold diluted in sterile 1× PBS buffer and inoculated into duplicate wells. After adsorption, monolayers were washed with 1× PBS buffer, overlaid with 1.0 mL of 3% carboxymethylcellulose in L15-2% FBS, and incubated at 37°C/5% CO2 for 7 days. After this period, cells were stained with 2% neutral red, incubated for 5 h at 37°C, and the number of plaques was converted to number of plaque-forming units per milliliter (PFU/mL).
Extraction of viral RNAs
Viral RNAs were extracted from infected cell supernatants and viral stocks using the QIAamp Viral RNA Kit (QIAGEN) according to the manufacturer's instructions. The RNAs were eluted in sterile water and stored at −20°C for later use.
Interference assays
In vitro interference experiments were carried out in C6/36 cells, a cell line derived from A. albopictus to investigate, in vitro, what we hypothesized to occur in vivo. In the infected control groups, C6/36 monolayers grown in 25 cm2 flasks were infected either with DENV-2 or YFV-17D viruses with an MOI of 0.1, and 7 days later, the infection was confirmed by IFA. In the superinfection exclusion groups, named sequential infection groups, 7 days after infection with one of the viruses, cells were infected with the second virus at the same MOI (i.e., cells that were initially infected with DENV-2 were infected with YFV-17D [DENV-2/YFV-17D] 7 days later; and conversely, those initially infected with YFV-17D were then infected with DENV-2 [YFV-17D/DENV-2]) after the same period of time. Twenty-four hours after the coinfection, culture medium aliquots from the coinfected cells were collected and stored at −20°C for the specific infection experiments. The same protocol was used to investigate the interference phenomenon between DENV-2 and the wild strain YFV-BeH111 after sequential infections. A mock-infected group, consisting of cells and 1× PBS, was used as reference for the infections.
Cell morphology analyses
Cell morphology was analyzed daily by optical microscopy (Nikon Eclipse 50i microscope) during the 7 days anteceding the sequential infection assay and during 4 days after the second infection. Cell morphologies from control and sequential infection groups were used to detect changes that could indicate the occurrence of cell injury.
Immunofluorescence assays
C6/36 cells were cultivated in a chamber-slide system (Lab-Tek System; SIGMA) and the IFAs performed on the third day after infection with either YFV or DENV-2, depending on the sequential infection. Infected cells were initially incubated with primary antibodies against DENV-2 and YFV (1:100 and 1:50 dilution, respectively), generated in mice and rabbits, respectively, and then with goat secondary antibodies against mouse and rabbit IgGs labeled with FITC and TRITC (Sigma-Aldrich®) at 1:300 and 1:600 dilutions, respectively. Images were registered in a BX40 fluorescence microscope (Olympus) and captured by a Q-Color 5 camera (Olympus) with the Q-Capture 2.68.6 software, always under the same exposition parameters.
Viral replication assays by real-time PCR
Quantitative real-time RT-PCR used oligonucleotide primers for DENV-2 as described by Houng et al. (2001), and the oligonucleotide primers for YFV-17D were designed according to the general guidelines for primer design. Also, RT-PCRs performed on YFV RNAs using DENV-2 primers under the same protocol as for DENV-2 resulted in no cross-amplification. Primer sequences were as follows: DENV-2 (sense: 5′AAGGTGAGATGAAGCTGTAGTCTC-3′; antisense: 5′-CATTCCATTTTCTGGCGTTCT-3′) [10] and YFV-17D (sense: 5′-TTTGCCACTGCTAAGCTGTGA-3′; antisense: 5′-CGCAAAACCTGGTTTCTGGG-3′). Total RNAs collected on the interference assays as well as the viral RNA for the standard curves were extracted and used in SYBR Green-based RT-PCRs (Applied Biosystems) according to the manufacturer's directions. Real-time RT-PCR protocols consisted of 20 min at 48°C, 10 min at 95°C, followed by 40 cycles of 15 s at 95°C and 1 min at 60°C. The standard curve was constructed with 10-fold dilutions of the viral stock previously diluted to 106 PFU/mL and ranged from 101 to 106 PFU/mL. The specificity of the reaction for the each virus was analyzed through the melting temperature (Tm).
Statistical analysis
Differences among the experimental group averages were evaluated by two-way ANOVA and Bonferroni's post-test, and considered significant when p < 0.05.
Results
Cell morphology analyses
Cells infected either with DENV-2 or YFV-17D showed no cytopathic effects. In the cells that were initially infected with YFV-17D and 7 days later with DENV-2 (YFV-17D/DENV-2), the cell morphology analyses revealed a normal aspect (Fig. 1A), suggesting that, in this group, the viral replication of both viruses did not induce cell injury. In contrast, cells that were initially infected with DENV-2 and 7 days later with YFV-17D (DENV-2/YFV-17D) showed significant changes in morphology (Fig. 1B), characterized by the presence of oversized and clustered cells. These cell morphologies were not observed either in the control cells or in the YFV-17D/DENV-2 experiments, showing that this sequence of infection accelerated the fusion-from-within mechanism, a finding that may occur to C6/36 cells infected with DENV for long periods of time. A higher level of cell death after 4 days of infection was observed with YFV-BeH111 (DENV-2/YFV-Be111) when submitted to sequential infection assays (Fig. 1C). Both groups of sequentially infected cells (interference assays) were compared to noninfected cells (Fig. 1D).
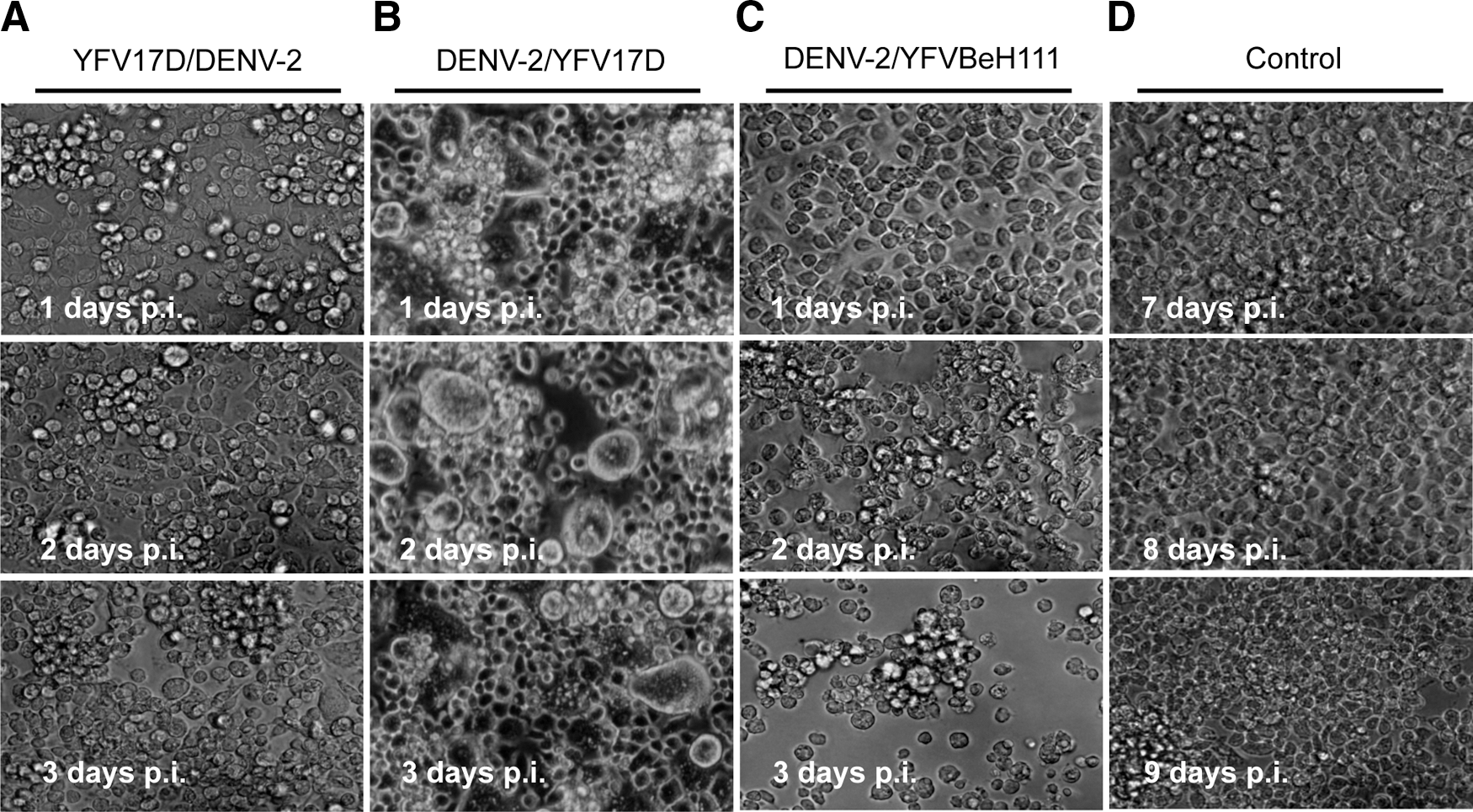
C6/36 cell morphology analyses after coinfections.
Replication profiles after DENV-2, YFV-17D, or YFV-BeH111 virus single infection
Viral replication on C6/36 cells was assessed by measuring the number of RNA copies/mL on the supernatants of infected cells. After 24 h of infection, replication profiles for all viruses in the control group presented a considerable decrease in the viral load compared to the initial inoculum, corresponding to 1.0 × 105 PFU/mL for DENV-2 strain, 1.0 × 106 PFU/mL for YFV-17D strain, and 1.0 × 106 PFU/mL for YFV-BeH111 strain; a phenomenon known as the eclipse period (Fig. 2). In the next 2 days, viral replication increased, and by the third day, viral load levels reached 7.0 × 105, 1.0 × 105, and 8.5 × 103 PFU/mL, respectively, for DENV-2, YFV-17D, and YFV-BeH111 strains (Fig. 2A–C, respectively). High viral loads were detected up to the eighth day for all viruses, evidencing a persistent infection in C6/36 cells.

Replication profiles of DENV-2, YFV-17D, or YFV-BeH111 viruses in C6/36 cells. C6/36 cells were single infected with DENV-2
Replication profiles of DENV-2 and YFV-17D after sequential infection assays
Quantification of viral replication in C6/36 cells subjected to interference assays shows that in the DENV-2/YFV-17D group, the YFV-17D replication was significantly reduced, whereas DENV-2 replication was kept at a high level (Fig. 3A). The extent of impairment of YFV-17D replication, caused by the established DENV-2 infection, can be better represented compared to the YFV-17D regular replication profile when single infection was performed (Fig. 3B). Similarly, in the YFV-17D/DENV-2 group, the DENV-2 replication was significantly reduced, whereas YFV-17D replication was kept at high levels (Fig. 3C), as also shown in panel D, which compares with the regular replication profile of DENV-2 when single infection was performed. These results show that, when the cells are already chronically infected with the other virus, although to a different extent, both YFV-17D and DENV-2 replication profiles in C6/36 cells are significantly impaired.

Interference assays: replication profiles obtained for DENV-2 and YFV-17D when sequentially coinfection was performed.
Replication profiles of DENV-2 and YFV wild-type strain (YFV-BeH111) after sequential infection assays
To investigate if the low replication of the YFV observed in the interference assays could be due to a particular behavior of the vaccine strain, YFV-17D, we repeated the sequential infection assay now using the YFV-BeH111 strain, a wild-type YFV isolated from a patient. Quantification of YFV-BeH111 replication in C6/36 cells subjected to sequential infection assays (DENV-2/YFV-BeH111) also showed that YFV-BeH111 replication was significantly impaired, whereas DENV-2 replication was kept at a high level (Fig. 4). It is important to say that sequential infection in the DENV-2/YFV-BeH111 group was highly cytotoxic to these cells, and the experiment could be only assessed up to 4 days, suggesting that this sequential infection accelerated the process of cell death. Due to the YFV-BeH111 cytotoxicity to C6/36 cells, the YFV-BeH111/DENV-2 assay could not be performed because after a single infection with YFV-BeH111, cells barely survived until the eighth day.

Interference assays: replication profiles obtained for DENV-2 and YFV wild strain (YFV-BeH111) when sequentially coinfection was performed. C6/36 cells were first infected with DENV-2 virus, and after 7 days were coinfected with YFV-BeH111. Quantification of viral replication in C6/36 reveals that a significantly lower replication of YFV-BeH111 is observed when compared with DENV-2 replication profile, and also to YFV-BeH111 replication profile when single infection was performed (inner panel). As described in the Results section, due to the high cytotoxicity of the YFV-BeH111 strain, this assay could be followed only up to 4 days. Data shown represent the average ± SEM of three independent experiments. **p < 0.01 versus control, inner panel, two-way ANOVA.
Coinfection assays evaluated by IFAs
To further validate our data, we performed IFAs in C6/36 cells subjected to DENV-2/YFV-17D, as described earlier. A strong fluorescence for DENV-2 is observed (Fig. 5A), but not for YFV-17D (Fig. 5B), showing that the YFV-17D infection was minimal. Figure 5A and B corresponds to the same field and captured with different wavelengths specific for FITC and TRITC, respectively, with the same time of exposition. As positive controls, single infections either with DENV-2 or YFV-17D are shown in Figure 5C and D, respectively. Noninfected cells (Fig. 5E, F) were used as negative controls for both virus infections.

Interference assays: evaluation by IFAs. IFAs were performed in C6/36 cells subjected to sequential coinfections (a first infection with DENV-2 and 7 days later with YFV-17D) and single infections using antibodies against either DENV-2 (left panels) or YFV-17D (right panels). Results from the sequential coinfection group show that DENV-2 replication is kept at high levels after YFV-17D infection
Discussion
DENV cause the most important arboviral disease in the world, accounting for ∼390 million infected humans every year (Guzman and Kouri 2003, Halstead 2007, Bhatt et al. 2013). Although yellow fever is restricted endemically to the forested regions of Central and South Americas, and to Africa, the YFV is periodically responsible for isolated cases and outbreaks of different extents in the Americas (Vasconcelos 2003). Nevertheless, since the beginning of the 80s, the sylvatic form of yellow fever has resurged in South American countries, including Brazil (Vasconcelos et al. 2001, Vasconcelos 2003), raising the concern for the possible reurbanization of the disease. Several preexisting epidemiological conditions favor this occurrence, such as the circulation of YFV in the region, a large population of susceptible individuals, and the presence of the mosquito vector A. aegypti (Prata 2000).
The same epidemiological profile is also observed in some Asian countries where, despite the existence of the aforementioned conditions, the occurrence of urban yellow fever has never been reported. Several hypotheses have tried to explain this epidemiological silence in Asia, such as the immunological cross-protection by heterologous flaviviruses and reduced vector competence for YFV from the Asian strain of A. aegypti (Monath 1997). Cross-immunity between flavivirus infections has indeed been related to a reduced severity of the disease, but not to a complete protection against YFV (Xiao et al. 2003, Izurieta et al. 2009). Also, concerning vector competence, many studies have demonstrated the ability of Asian strain of A. aegypti to transmit YFV (Lourenço-de-Oliveira 2002), hence ruling out such possible explanation. Amaku et al. (2011) have constructed a mathematical model that may explain the lack of YFV circulation in Asia. They concluded that the association of cross-protection between flavivirus infections, especially dengue, would protect against YFV infections, and that the low prevalence of A. albopictus in Africa explains the coexistence of both infections in Africa. These hypotheses have been disproved before (Xiao et al. 2003, Izurieta et al. 2009), and our data show that there could be a competition between viruses inside these vectors, as they were evident in the cell cultures, a hypothesis declined by their mathematical model. The hypotheses presented here and discussed ahead can serve as a starting point for testing these findings in mosquitoes to evaluate if this phenomenon is observed in vivo.
In the present study, we used an in vitro approach to demonstrate the interference of one virus to the replication of the other, a phenomenon known as superinfection exclusion. The A. albopictus C6/36 cells, a continuous cell line from A. albopictus, were subjected to single or sequential coinfections by the two viruses, followed by evaluation of the replication profile for each virus. Our data show that infection of C6/36 cells either with DENV-2 or YFV led to a significant impairment of the subsequent infection by the other virus. The DENV-2/YFV-17D interference assay showed that DENV-2 kept a high replication profile, whereas YFV-17D replication was significantly impaired (Fig. 3A), in opposition to the high replication profile in cells infected only with YFV-17D (Figs. 2B and 3A inner panel). We also performed an assay inverting the order of infections (YFV-17D/DENV-2) and observed interference on DENV-2 replication (Fig. 3B), but to a lesser extent, compared to DENV-2 followed by YFV-17D infections. Similarly, it has been recently described that C6/36 cells infected with a high MOI of DENV led to a significant replication inhibition of a subsequent infection with CHIKV (Potiwat et al. 2011).
Furthermore, the daily morphological analyses revealed changes in morphology following YFV-17D infection in cells initially infected by DENV-2 (Fig. 1B), a phenomenon that was not reported by Karpf et al. (1997). Interestingly, our data show that cells first infected with YFV-17D and then infected with DENV-2 presented with no changes in morphology (Fig. 1A).
To rule out that those results could be due to the attenuated condition of the vaccine YFV strain used in our study (YFV-17D), we also investigated the presence of interference using the YFV wild-type strain, YFV-BeH111. The results obtained with YFV-BeH111 consistently reproduced the data previously obtained with the YFV-17D strain (Fig. 4) and showed YFV-BeH111 infection to be highly cytotoxic to C6/36 cells (Fig. 1C).
Some previously reported superinfection exclusion studies corroborate our findings, where the preinfection of A. albopictus cells with the alphavirus, Sindbis virus, protected from a second homologous infection by alphaviruses such as Semliki Forest virus, Una, or CHIKV; however, allowed the infection by a Bunyavirus (Snowshoe Hare virus) (Eaton 1979). Similarly, cells infected with the Sindbis virus (an alphavirus) interfered with the infection by other alphavirus, but not by viruses from other family, such as YFV (Eaton 1979).
The present work brings the first evidence of the occurrence of superinfection exclusion between the dengue and the yellow fever viruses and sheds light as a possible explanation to the observed phenomenon of lack of urban yellow fever epidemics in areas where dengue is highly prevalent. The importance of cross-immunity and vector capacity of A. aegypti cannot be completely ruled out, but our findings raise the questions if this phenomenon might not happen in nature.
Footnotes
Acknowledgments
The authors thank Dr. Pedro Vasconcellos (Evandro Chagas Institute) and Dr. Robert E. Shope for kindly donating the YFV-BeH111 wild strain and YFV-17D, respectively. This study was supported by the São Paulo State Research Foundation (FAPESP; grant no. 2003/07959-0), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). E.P.A. held a fellowship from CAPES and BALF and a Research Fellowship from CNPq.
Author Disclosure Statement
No competing financial interests exist.