
Abstract
Norway rats are an abundant synanthropic species in urban settings and serve as reservoirs for many pathogens. Attempts to control their populations have met with little success. Recent genetic studies suggest that local populations are structured and few individuals move significant distances, but there is substantial gene flow. To understand these observations and their implications on control strategies, we genotyped 722 rats from 20 alleys in Baltimore to establish paternity for 180 embryos. Up to 88 males may have contributed to the litters. All litters were sired by ≥2 males, with an average of 4.9 (range 2–7) males. For dams and sires with known locations, most matings (71.7%; n = 46) occurred among animals from different alleys. The average distance between sires and dams was 114 meters (range 8–352 meters). In 10/17 (58.8%) litters, the majority of the identified sires were captured in different alleys than the females. Sires were significantly less related to females than were the males captured in the females' alleys. Although rats may generally restrict their movements, either receptive females and/or breeding males engage in mate-seeking behaviors that extend beyond movement patterns at other times. This geographically extends the sizes of local populations and buffers them from the impacts of control strategies that focus on local infestations.
Introduction
N
Why control fails is not entirely clear. However, studies of urban populations describe genetic structures indicative of spatial isolation, but with sizes of local populations and gene flow, larger than expected (e.g., Gardner-Santana et al. 2009; Pearse and Anderson 2009). In Baltimore, related individuals extended nearly 2 km—far beyond the block level (ca 75–80 meters). A proposed reason for these apparently disparate observations was that R. norvegicus engaged in promiscuity. It has been documented in laboratory conditions (Macdonald et al. 1999; Hinson et al. 2006), confined seminatural areas (Calhoun 1962; Berdoy et al. 1995), and behavioral observations of mating chases by several male wild rats of individual females (Davis 1953; Glass et al. 1989; Macdonald et al. 1999). Promiscuity is also possible because of the large litter sizes of R. norvegicus that could allow multiple males to contribute to females' litters (Dean et al. 2006). However, females are sexually receptive only during a short (6–12 h) time (Connor and Davis 1980), which could limit the behavior.
We characterized the spatial patterns of the mating system of free-ranging R. norvegicus, in urban populations in Baltimore, Maryland, using microsatellite markers from laboratory rat strains. This study describes mate selection by female rats and characterizes reproductively successful male rats.
Materials and Methods
Rats were collected using Tomahawk live traps (Tomahawk, WI) in 20 central alleys of residential blocks in Baltimore, Maryland (Gardner-Santana et al. 2009; Glass et al. 1988, 1989, 2009). Five regions of residential neighborhoods within the city were selected and four adjacent blocks were sampled in each region (Glass et al. 2009). The average distance among block centroids within a region was 142.4 meters (range = 54.1–299.4 meters). This clustered sampling was based on previous studies showing that dispersal of adult rats was rarely beyond 1–2 blocks and that genotyping accurately assigned the majority (>90%) of rats to their alleys of capture (Gardner-Santana et al. 2009). The five regions were from 3.34 to 11.46 km (mean = 6.62 km) apart. Movement of rats among regions was expected to be minimal.
Within each alley, 15 traps were set at dusk, baited with peanut butter, and retrieved the following morning. Areas were trapped several nights until ∼50 rats were caught per region. Trapping was repeated ∼1 year later to obtain 100 total individuals per region. Rat population sizes within alleys were estimated by catch per unit effort methods (Caughley 1977; Glass et al. 2009). The proportion of removed rats was estimated using the population size estimate as the denominator and the numbers of captured rats as the numerator (Glass et al. 2009).
Animals were returned to the laboratory, euthanized by means of CO2 inhalation, standard necropsy data were gathered, and embryos in the second or third trimester of pregnancy were collected. Protocols were approved by the Johns Hopkins University Institutional Review Board (RA06H302). DNA was extracted from adult kidneys or embryonic tissue, respectively.
Mothers, embryos, and all captured males (as potential fathers) were genotyped at nine (CA)n dinucleotide microsatellite loci from different linkage groups (D1Cebr3, D1Cebr9, D2Cebr1, D3Cebr3, D4Cebr2, D5Cebr1, D6Cebr1, D10Cebr1, and D20Cebr1; Giraudeau et al. 1999). These primers were highly polymorphic in Baltimore rats, providing adequate power to elucidate associations between related individuals within the sample (Gardner-Santana et al. 2009; Sefc and Koblműller 2009). One primer of each pair was fluorescently labeled to detect and determine the size of microsatellite loci using an ABI Prism™ 3100-Avant Automated Genetic Analyzer (Foster City, CA). PCR amplifications involved 25 μL multiplex reactions containing either 3.13 μM total of five primer pairs (0.5 μM D2Cebr1, 1 μM D4Cebr2, 0.38 μM D5Cebr1, 0.75 μM D10Cebr1, and 0.5 μM D1Cebr9) or 2.25 μM total of four primer pairs (0.25 μM D1Cebr3, 0.5 μM D3Cebr3, 0.5 μM D6Cebr1, and 1 μM D20Cebr1), 2.5 mM MgCl2, 3 units of Taq DNA polymerase, 0.05 mM dNTPs, and 4 μL of the DNA extraction. The thermal profile was as follows: 95°C for 90 s; 30 cycles of 95°C for 40 s, 55°C for 40 s, 70°C for 120 s; 70°C for 1 h; and 4°C for 120 s. To 14.5 μL loading buffer (0.5 μL of GS-400HD ROX size standard and 14 μL of formamide), 0.5 μL of the PCR product was added.
Individuals were genotyped at each locus (GENESCAN™ 3.7 software) and alleles were assigned using GENOTYPER™ 3.7 software. We calculated the pairwise relatedness (R) to determine levels of relatedness of individuals (SPAGeDi; Hardy and Vekemans 2002). Movement between alleys was elucidated using an assignment test with the leave-one-out procedure (GeneClass 2.0; Piry et al. 2004). A partial Bayesian approach (Rannala and Mountain 1997) assigned individuals to the 20 alleys.
Paternity assignment simulated paternity with allele frequencies from all genotyped males—as potential fathers, the known mothers, and all their offspring (CERVUS 3.0.3; Kalinowski et al. 2007). We used 10,000 iterations, 34 possible fathers per offspring, 0.5 proportion of fathers sampled, and the standard error rate of 0.01 (Marshall et al. 1998; Taylor et al. 2000; Denny et al. 2002; Holleley et al. 2006). Simulated genotypes were used to calculate likelihood ratios as an LOD score for each candidate father. Assignment of fathers to offspring was made by comparing LOD scores of actual genotypes to those of simulated genotypes. We calculated the pairwise relatedness coefficient (Queller and Goodnight 1989) to determine the relatedness of mother–father pairs as well as the mother to other males in her alley of capture (SPAGeDi; Hardy and Vekemans 2002).
Determination of multiple paternity was initially performed by assigning sires (CERVUS 3.0.3; Kalinowski et al. 2007). More than one assigned sire per litter inferred multiple paternity. Following assignment, the genotypes of the remaining embryos were examined, maternal alleles were eliminated, and paternal alleles identified at the nine loci. We established the minimal number of additional fathers needed for unassigned offspring by counting the number of remaining unique paternal alleles (n) at each locus. For each n alleles (truncate n/2 + n modulo 2 for n ≥ 2; and 1 for n = 1) additional sires were considered to exist. The total number of unidentified sires for a litter was the maximum of (truncate n/2 + n modulo 2) over all nine loci. The total number of sires for a litter was the total number of assigned males plus the number of additional unidentified males. The geographic localities and demographic characteristics of identified sires were compared to remaining sampled adult males. Relatedness of sires to dams was compared with males residing in the alley where the female was captured using a two-way analysis of variance with regions in the city (Gardner-Santana et al. 2009) as one effect and whether males either were sires or were unassigned males from the female alleys as the second effect.
Results
A total of 542 rats were captured and genotyped, and estimated population sizes averaged 49.5 rats/alley, ranging from 2 to 216 individuals. It was estimated that 56% of rats were removed from the alleys during trapping (Glass et al. 2009). Based on necropsy, 53 (17.9% of males) of the trapped male rats were juveniles or subadults and not reproductively active. None was identified as putative sires in the analyses. Among captured adult rats, 245 were females, leaving 244 males as potential sires.
One hundred eighty pups were obtained from 18 females trapped in 12 alleys (Fig. 1; Table 1). Litter sizes ranged from 4 to 21 (mean [ ± SD] = 10.0 ± 4.1). Pregnant females ranged in size from 237 to 585 g (mean [ ± SD] = 433.6 ± 95.8). Sample sizes were too small to detect any relationship between the female body size and litter size (Table 1).
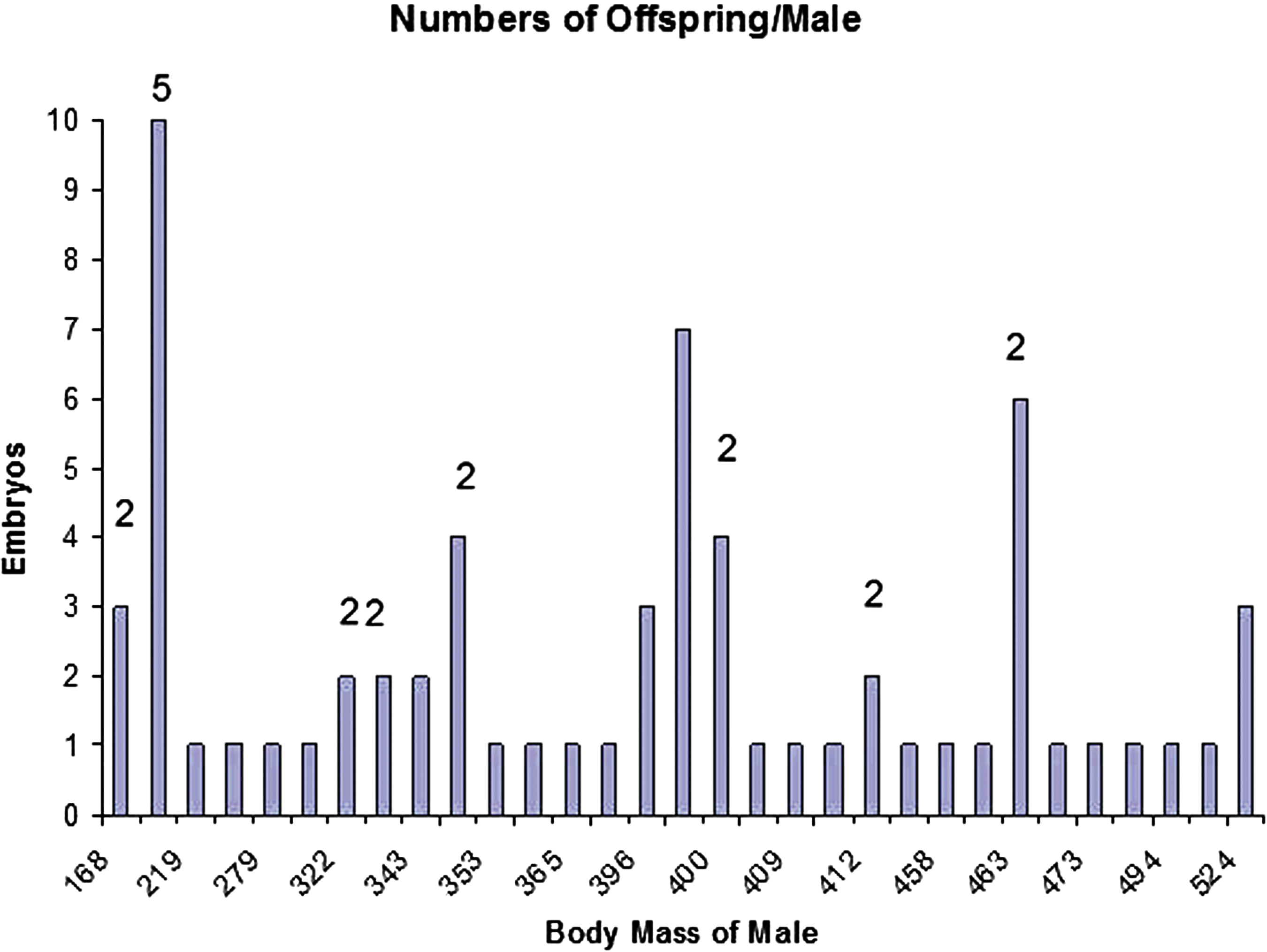
Histogram of male reproductive success in relationship to body mass (x axis). Y axis is number of embryos assigned to each male. The number above bars indicates the number of females associated with that success. No number indicates only a single female contributed to male breeding.
Bolded relatedness values are for sires that differ in their relatedness with the dams when compared with males from her assigned alley.
Based on the nine loci, these populations were structured comparably to those previously reported from 11 different blocks in the city (Gardner-Santana et al. 2009). Estimates of neighborhood size (n = 48.7 vs. n = 44.5), distance between related individuals (1.1 vs. 1.7 km), and axial dispersal distance (81.5 vs. 62 meters) did not differ from the earlier study. In addition, nearly all animals were correctly assigned to their alley of capture. Among sampled rats, 97% (526/542) was assigned, based on their genotype, to the alley in which they were captured. Of the remaining individuals, 12/16 were assigned to one of the three remaining sampled alleys in each of the five regions. None was assigned to sampled alleys in the other regions of the city. Thus, individuals residing within blocks appeared relatively sedentary showing limited evidence of between-alley movement. The distance between related individuals, however, suggested more movement associated with breeding than was evident from trapping.
Paternity analyses indicated that all 18 litters had more than one sire. A total of 88 males were inferred to be sires for at least one of the pups in the 18 litters and 34 (38.6%) were trapped and genotyped. On average, nearly five males contributed to each litter (mean [ ± SD] = 4.9 ± 1.5), with a range of 2–7 sires/litter. In 14/18 (77.8%) litters, more than one of the trapped males was assigned as a father. Among the remaining litters, either none of the sires were trapped (one litter) or only one trapped male was identified as the sire and the remaining sires were inferred from the genotypes of the unassigned offspring (three litters). Among the dam–sire dyads, the numbers of trapped fathers ranged from 0 to 4 (mean [ ± SD] = 2.5 ± 1.25; n = 45), and these males accounted for 70/180 (38.9%) of the pups.
Identified sires tended to be smaller than dams (399.2 ± 105.7 g; range 168–611 g; Fig. 1), but males from among the smallest reproductively active individuals to the largest were sires. There was no obvious relationship between the male body size and reproductive success or the number of females with which they bred (Fig. 1). Eight of the 34 (23.5%) identified males successfully mated with multiple females. Seven of the males who successfully mated with >1 female fathered pups with two females, while one male fathered pups with five females (Fig. 1). On average, males who mated with more than one female produced no more offspring per female (1.7 ± 1.1; x ± SD) than did males who mated with only a single female (1.5 ± 1.3; x ± SD).
The locations where females and identified male consorts were trapped show that, for four of the five regions, females and their mates primarily came from different alleys (Figs. 2 and 3). Seven of the eight (87.5%) males that bred with more than a single female resided in alleys different from at least one of the mothers, and, for 3/8 (37.5%) of these males, none of their mates resided in the same alley as they did. Among the 26 males that bred with only a single female, 18 (69.2%) were from different alleys than the dams. For 10/17 (58.8%) females (one female rat from the fifth region had no identified consorts and was not mapped; Fig. 3), the majority or all of the consorts came from a different alley than where she resided. For half (n = 5) of these females, all of their mates resided in the same alley but it was an alley different from hers (Fig. 3). The average distance between sites of capture for dams and sires was 114.3 ± 75.3 meters (mean ± SD) ranging from 8.0 to 352.5 meters. The distribution of distances between sites of capture of dams and sires tended to peak at intermediate distances (100–200 meters), which characterized distances between where the largest numbers of traps were set (Fig. 2).
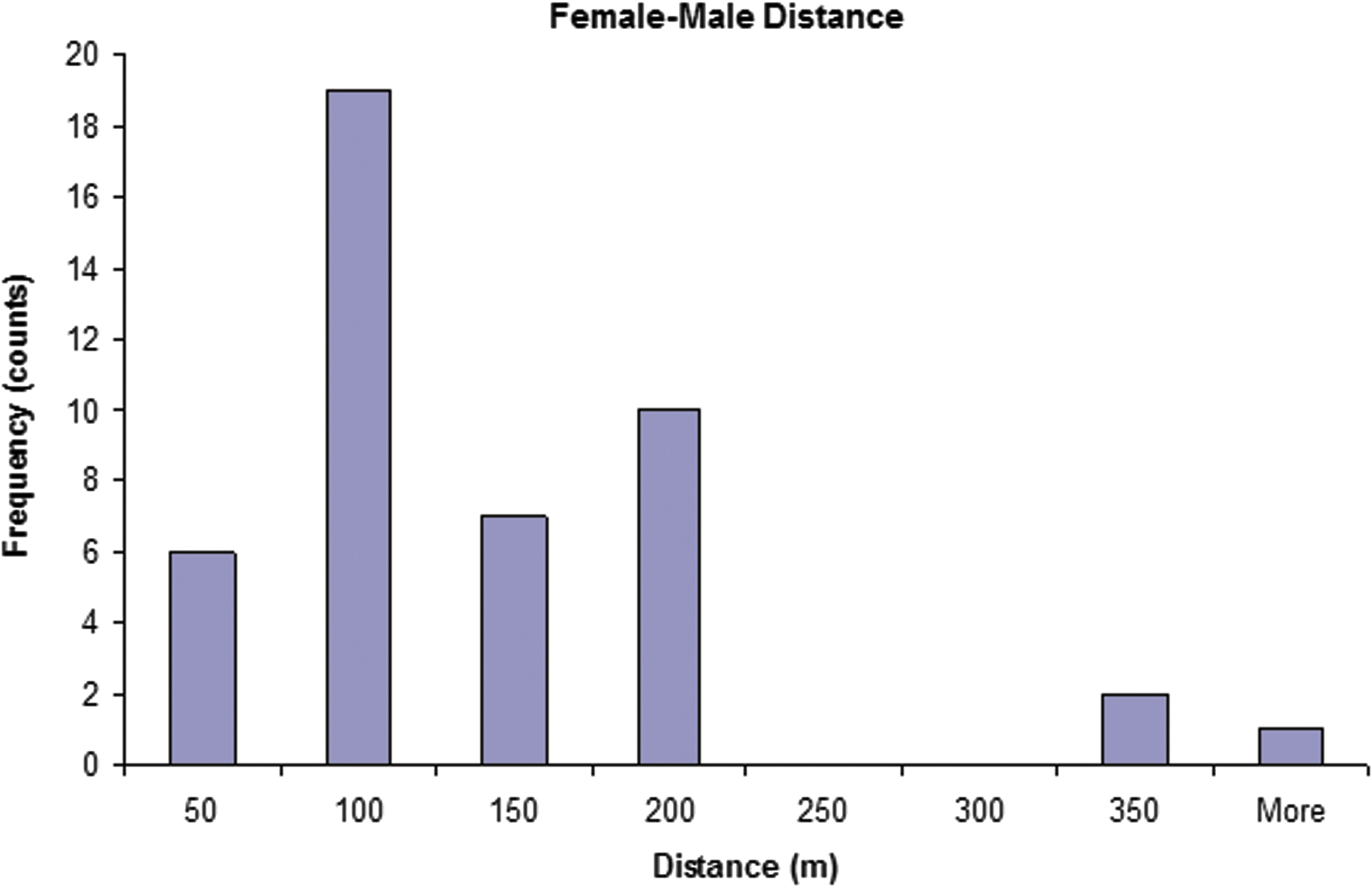
Histogram of female–male partner distances between captures (x axis). Within-alley distances were typically less than 80 m—indicating that most matings took place among individuals from different alleys.
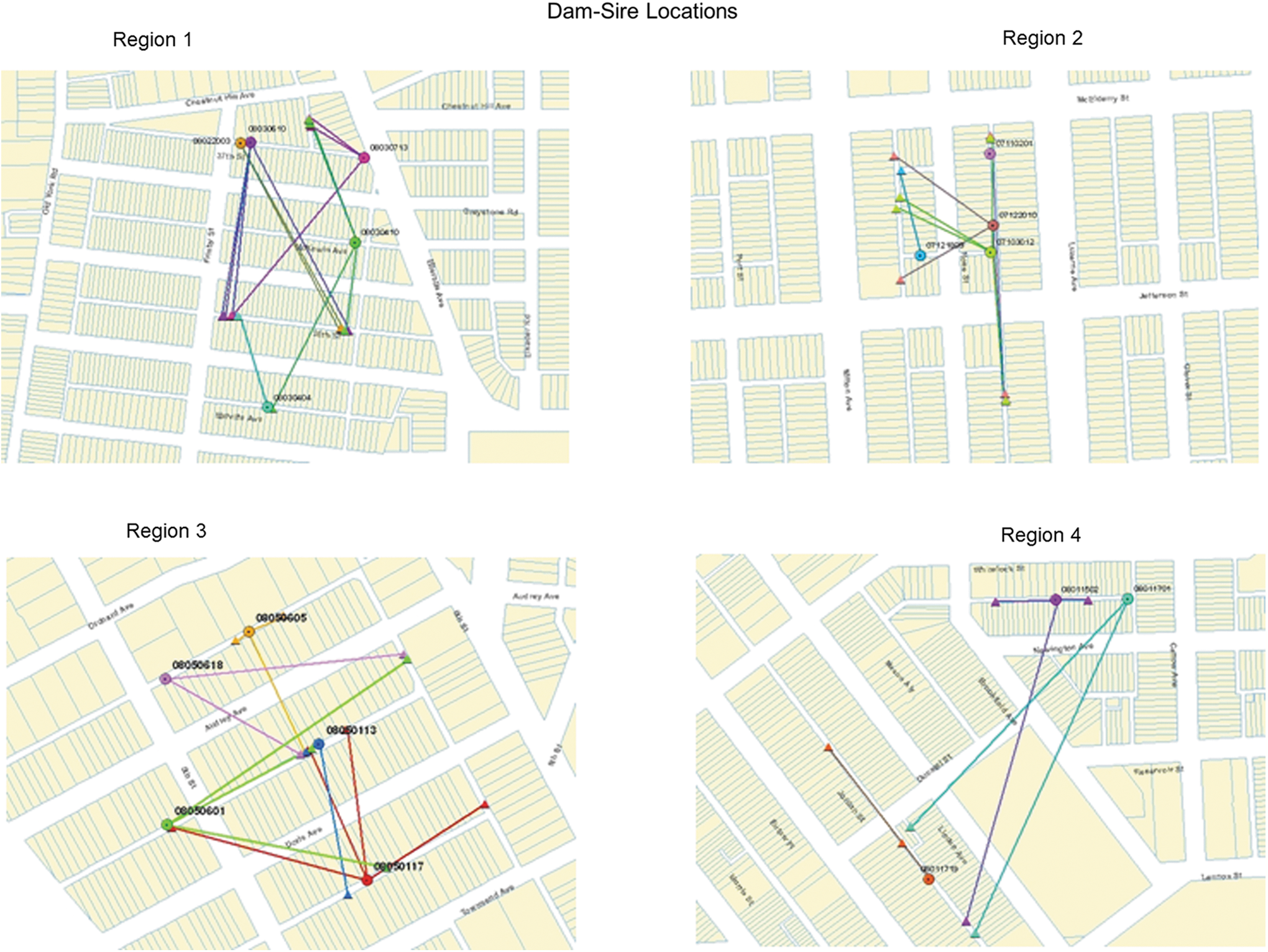
Mapping of females and their consorts in four regions of Baltimore. Females are shown as circles and males as triangles. Females and their consorts are joined by connecting lines. Broad white areas are roads, while narrow white areas are central alleys with the blocks. Properties are shaded.
Similar to previous reports, there were differences in the relatedness among males and females across regions of the city (F = 2.78; 3, 235 df; p = 0.04), with a north–south gradient from region 2 (R = 0.052) through central Baltimore (R = 0.088 in region 4 and R = 0.136 in region 1) to southern Baltimore (R = 0.179) (Gardner-Santana et al. 2009). Regardless of region, females tended to breed with males that were significantly less related than were the males in her alley (F = 7.53; 1, 235 df; p = 0.006). This effect held across all the regions of the city (interaction of region and mate was not significant F = 0.75; df = 3, 235; p = 0.523).
Discussion
These results indicate that, contrary to current perspectives on urban rats, movements in mate acquisition differ from other activities of rats' daily lives. This likely has remained unrecognized because mate seeking during the short postpartum estrus represents a tiny fraction of the lifetime activity budget of a wild rat (Calhoun 1962). The practical effect is to geographically extend the size of the potential breeding pool and buffer local (block level) subpopulations from perturbations associated with targeted control efforts such as food reductions, baiting, and environmental modifications.
Historical studies of Norway rat population studies have contributed to and relied on mate selection theory (Trivers 1972; Jennions and Petrie 2000; Dean et al. 2006; Berdoy and Drickamer 2007). Descriptions of territoriality and defense of nest and food sites led to the interpretation of the social system as one where dominant females and males were the primary reproductive contributors. Males fought for access to receptive females and dominant, pregnant, or nursing females inhibited the successful reproduction of subordinate females (Calhoun 1962). The occurrence of a 6–12 h postpartum estrus in females appeared to make it feasible for alpha males and females to dominate reproductive effort. Coupled with reported nest cannibalism, there seemed a strong pressure for females to mate with dominant males in defensible nest sites (Calhoun 1962) and restrict their movements. These theories, coupled with field experiments manipulating local populations (Davis 1953), led to the intervention strategy of targeting infestations at the neighborhood/block level and these approaches remain in effect (Keiner 2005).
The frequency of multiple male matings observed in this study is high even when compared with other rodent species (Waterman 2007). The patterns observed here are probably most similar to results reported by King et al. (2014) examining the conger Rattus rattus. They similarly reported a high proportion of multiple paternity, a large fraction of pups with unassigned sires and evidence of substantial movement in and around local habitat (remnant forest). Although their study design did not allow them to identify actual movement distances, it appeared that more breeding males moved further than females.
Perhaps the most striking divergence of our observations from theory is the lack of any relationship between the male body size and reproductive success (Fig. 1), when it is presumed to play such a key role in establishing a territory, access to food, and nest sites (Davis 1953; Berdoy et al. 1995). Direct aggression results in physical injury, especially among male rats (Glass et al. 1988, 2009) and has been assumed related to establishing dominance. However, there is no relationship between the male body size and their ability to successfully mate with females, and little evidence that females pair with dominant males near their burrows (Figs. 1 and 3). This suggests that other drivers influence either male investment in aggressive interactions or females' choice in mates.
The relative rarity of females mating with males near their burrows is indicated by the distances between females and sires (Table 1; Fig. 3), which exceeds the length of an alley and approaches the average distances among alleys within the city. The only identifiable aspect of mate selection by females was choosing mates that were substantially less related to them than were males in their alleys. Females also often mated with multiple males from the same adjacent alley. Inbreeding of rat populations in Baltimore varied geographically, ranging from nearly outbred populations in the north to moderately inbred populations in the south of the city. However, females from all areas selected males from nearby alleys rather than the one in which they resided. The choice of unrelated individuals for mates has often been postulated to increase disease resistance (Klein 2000) and decrease the likelihood of accumulating lethal genetic variants (Stockley 2003). However, using this strategy regardless of the level of local inbreeding suggests that it does not accurately evaluate “true” inbreeding potential. Rather, the cues appear to involve “familiarity versus unfamiliarity” (Roberts and Gosling 2003). Regardless, a consequence of this strategy is the potential of homogenizing transmissible pathogen prevalence within urban areas. Whether it is primarily females or groups of males that travel away from their home alleys seeking mates is not answered by this study. While it might seem most parsimonious to presume that a single female moves away from her home alley (as opposed to a cohort of males), what observations that have been reported (Davis 1953; Calhoun 1962; Glass et al. 1989) indicate groups of males engaged in “mating chases.” The geographic origin of these groups is unclear. The question, likely, could be resolved by coupling direct observation and radiotelemetry studies.
These breeding patterns provide a rationale why control of synanthropic R. norvegicus populations remains challenging. Promiscuity and the selection of mates from other alleys buffer local populations from impacts of sporadic population reductions caused by control efforts (or other perturbations) (Pearse and Anderson 2009). The true extent of directed travel in this study is likely underestimated as the surveyed regions did not extend beyond adjacent blocks (∼300 meters; Fig. 3). Males and females from these farthest distances were dyads and suggest that the true extent of mate seeking behavior is likely further. The large proportion of unassigned, but inferred, sires indicates that mate seeking animals may venture much further than the immediate area around the home block to reproduce, further shielding populations from interventions. Consequently, control efforts need to be broadened, both in methods and geography, to substantially impact infestations.
Footnotes
Acknowledgments
This work was supported by the U.S. NSF Ecology of Infectious Diseases research program # EF0525751 and Emerging Pathogens Institute at the University of Florida and the College of Liberal Arts and Sciences, as part of the University of Florida Preeminence Initiative. The authors thank Melissa Nastacio for assistance in the laboratory.
Author Disclosure Statement
No competing financial interests exist.