
Abstract
We examined fleas collected from black-tailed prairie dog (Cynomys ludovicianus) burrows from 2009 through 2011 in five national park units east of the known distribution of active plague across the northern Great Plains for the presence of Yersinia pestis. Across all national park units, Oropsylla tuberculata and Oropsylla hirsuta were the most common fleas collected from prairie dog burrows, 42.4% and 56.9%, respectively, of the 3964 fleas collected from burrow swabbing. Using a nested PCR assay, we detected 200 Y. pestis–positive fleas from 3117 assays. In total, 6.4% of assayed fleas were Y. pestis positive and 13.9% of prairie dog burrows swabbed contained Y. pestis–positive fleas. Evidence of the presence of Y. pestis was observed at all national park units except Devils Tower National Monument in Wyoming. We detected the presence of Y. pestis without large die-offs, i.e., enzootic sylvatic plague, east of the known distribution of active plague and near the eastern edge of the present distribution of black-tailed prairie dogs. This study, in combination with previous work suggests that sylvatic plague likely occurs across the range of black-tailed prairie dogs and should now be treated as endemic across this range.
Introduction
S
Prairie dogs (Cynomys spp.) are an ecologically important species of short- and mixed-grass prairie ecosystems that are particularly susceptible to epizootic plague (Gage and Kosoy 2005, 2006). Their extirpation can have wide-ranging ecological effects (Eads and Biggins 2015). Prairie dog colonies east of the 103rd meridian were thought to be safeguarded from plague (Antolin et al. 2002). In 2005, black-tailed prairie dog (Cynomys ludovicianus) colonies on the Pine Ridge Indian Reservation in South Dakota experienced the first documented epizootic east of the naturalized plague zone (Rocke et al. 2008). This was followed by epizootics in the Conata Basin, South Dakota, in 2008 (Griebel 2009, Abbott et al. 2012). Outbreaks east of the 103rd meridian demonstrated formation of a new emergence zone, or area where sylvatic plague outbreaks had been historically rare and limited in distribution (Antolin et al. 2002, Adjemian et al. 2007). Spread of sylvatic plague out of its naturalized zone provided an opportunity to study how Y. pestis spreads on a landscape scale.
Until recently, the dominant paradigm for prairie dog–Y. pestis interactions focused on prairie dogs serving as highly susceptible epizootic hosts that experience nearly 100% mortality and exhibit large-scale outbreaks (Gage and Kosoy 2005, 2006). However, evidence for presence of enzootic plague (presence of Y. pestis without massive die-offs over large areas in short time spans) in black-tailed prairie dogs has recently accumulated from sites in Montana (Hanson et al 2007, Biggins et al. 2010, Jones and Britten 2010, Matchett et al. 2010, Jones et al. 2012). There is also evidence that individual prairie dogs survive exposure to Y. pestis (Cully et al. 1997, Pauli et al. 2006, Jones et al. 2012).
Limited research has been conducted in the recent sylvatic plague emergence zone (Rocke et al. 2008, Griebel 2009, and Lower Brule Sioux Tribe, unpublished data), but understanding the spread of Y. pestis beyond its previous distribution is very important to wildlife conservation. Studies documenting Y. pestis where plague is considered endemic may not provide an accurate picture of its dynamics in areas along the invasion front. Invasive species, such as Y. pestis, often exhibit fluctuating distributions across space and time (Lomolino et al. 2005, Lowell et al. 2015). This suggests plague transmission dynamics may differ between the naturalized plague zone and locations in the current emergence zone.
While prairie dogs have been well studied in this system, fleas are an understudied part of the sylvatic plague system despite being the primary mode of transmission of Y. pestis between individual hosts. By seeking blood meals from different hosts, fleas play a key role in the sylvatic plague cycle as transmitters of Y. pestis, yet they lack the capacity to disperse farther than a few meters on their own (Marshall 1982). In our study area, prairie dogs and their burrows are primarily infested with two species of “prairie dog” fleas: Oropsylla hirsuta and Oropsylla tuberculata (Brinkerhoff 2008, 2011, Jones 2010). Both Oropsylla species are capable of early-phase transmission of Y. pestis (Wilder et al. 2008a, b). Plague is commonly documented by prairie dog epizootics; however, detection of Y. pestis in fleas offers a more sensitive detection method (Hanson et al. 2007). An advantage of studying fleas directly is that there exists an effective way of reducing flea populations. Deltamethrin kills fleas, resulting in decreased transmission of plague and increased survival, even when plague-induced prairie dog die-offs are not apparent (Biggins et al. 2010, Matchett et al. 2010). Jones et al. (2012) demonstrated the efficacy of deltamethrin in preserving black-tailed prairie dog genetic diversity and increasing survivorship and densities of prairie dogs on dusted colonies during an epizootic in Montana.
We conducted surveys for Y. pestis in fleas on black-tailed prairie dog colonies across several national park units east of the naturalized plague zone. The objectives of this research were to detect enzootic plague and determine the prevalence of Y. pestis within fleas east of the naturalized plague zone, document the flea fauna of black-tailed prairie dog colonies, and suggest management strategies to prevent or mitigate plague epizootics.
Materials and Methods
Five national park units (National Park [NP], and National Monument [NM]) were examined for presence of sylvatic plague: Badlands NP (SD), Devils Tower NM (WY), Scotts Bluff NM (NE), Theodore Roosevelt NP (ND), and Wind Cave NP (SD), as well as colonies surrounding Badlands NP on Buffalo Gap National Grassland (SD) (Fig. 1). Localities were selected because they were east of the known distribution of plague and had prairie dog colonies of variable sizes that were of management interest to the National Park Service. Badlands NP, Buffalo Gap National Grassland (NG), and Wind Cave NP support reintroduced black-footed ferrets (Mustela nigripes). Within these parks, colonies were selected for survey based on presence of ferrets, colony size (1.6–305.3 ha,
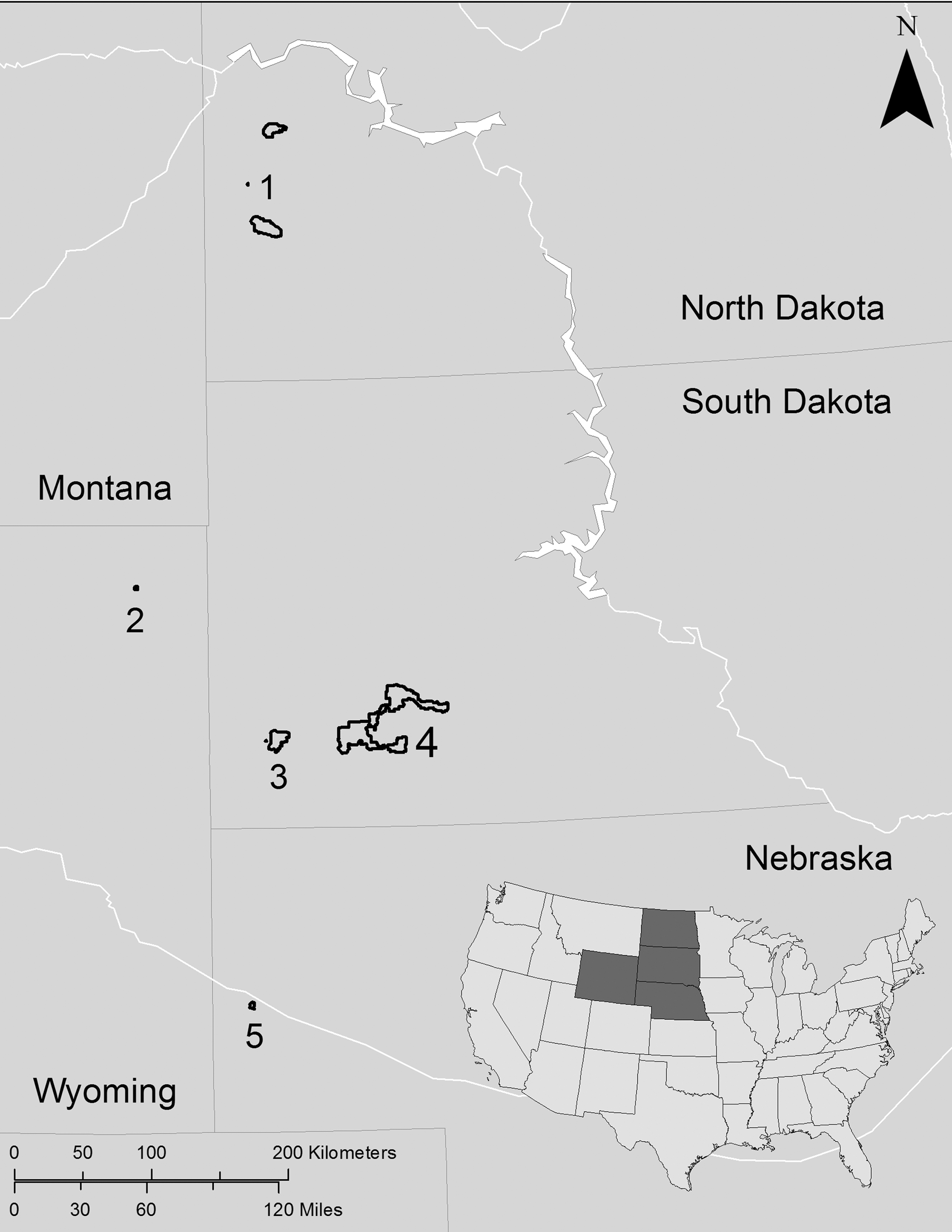
Map of National Park Units where black-tailed prairie dog colonies were sampled for fleas: 1, Theodore Roosevelt National Park (ND); 2, Devils Tower National Monument (WY); 3, Wind Cave National Park (SD); 4, Badlands National Park (SD); and 5, Scotts Bluff National Monument (NE).
Each national park unit was surveyed over 3 years (2009–2011); in 2009, each park was surveyed once between June and August, in 2010 colonies were surveyed April–October, and in 2011 colonies were surveyed May–July. During the 2010 field season each park was surveyed twice with the exceptions of Devils Tower NM (once), Scotts Bluff NM (once), and Wind Cave NP (three). In 2011, each park was surveyed once between May and August with the exception of Scotts Bluff NM (twice). In 2010 and 2011, the sampling season began earlier in an attempt to capture as much flea diversity as possible.
Fleas were collected directly from prairie dog burrows along the longest axis of each colony using a swabbing technique developed by Ubico et al. (1988). A 30-cm2-square flannel cloth was inserted into a prairie dog burrow for 2 min. Each burrow was swabbed only once regardless of flea presence/absence. An attempt was made to collect fleas from a minimum of 20 burrows or when 100 prairie dog burrows were swabbed for fleas. GPS coordinates were taken for each burrow where fleas were collected. Fleas were labeled individually, stored in 70% ethanol, and identified using the keys of Lewis (2002), Furman and Catts (1982), and Hubbard (1947). Whole genomic DNA was extracted using PrepGem (Zygem, New Zealand) extraction kits. Fleas were tested for presence of Y. pestis using a nested PCR assay modified from Hinnebusch et al. (1993) by Hanson et al. (2007; see also Jones and Britten 2010, Jones et al. 2011). Due to small sample sizes, fleas from the Johnson's Plateau colony at Theodore Roosevelt NP in 2009, Sanctuary colony at Wind Cave NP in 2009, and EP08 and WR05 colonies at Buffalo Gap NG in 2010 were pooled by burrow and species when analyzed. Due to the large number of fleas collected in 2011, a subset of O. tuberculata and O. hirsuta were randomly selected from each colony for analysis. The total number of fleas collected, prevalence of fleas in burrows (number of burrows with fleas/total number of burrows), mean intensity of flea infestation in burrows (number of fleas/number of burrows with fleas), total number of Y. pestis–positive fleas, total number of burrows with Y. pestis–positive fleas, prevalence of Y. pestis–positive fleas (number of Y. pestis–positive fleas/total number of fleas tested), and prevalence of Y. pestis in burrows (number of burrows with Y. pestis–positive fleas/total number of burrows with fleas tested) were estimated for each colony and national park with 95% confidence intervals using Quantitative Parasitology version 3.0 (Margolis et al. 1982, Rózsa et al. 2000).
Aggregation was estimated for fleas in burrows and Y. pestis–positive fleas in burrows for each colony and national park. Coefficient of aggregation (k) and variance to mean ratio (
Results
We swabbed 5236 prairie dog burrows for fleas from 2009 through 2011. We collected 3964 fleas from 23.5% (1231/5236) of burrows swabbed (Table 1). The majority of fleas collected were O. tuberculata (42.2%; 1673/3964) and O. hirsuta (56.9%; 2257/3964, Table 2). O. tuberculata was proportionally the most abundant flea species collected from prairie dog burrows in March–May, whereas O. hirsuta was proportionally the most abundant from June–October (Fig. 2, Table 2). Less than 1% were other species (33/3964; 12 Oropsylla spp, one Foxella ignota, one Ctenophthalmus pseudayrtes, and 19 unidentifiable fleas). Across the colonies surveyed, the number of fleas collected from individual burrows ranged from 0 to 224 total fleas (0–177 O. tuberculata and 0–161 O. hirsuta; Fig. 2, Table 2; Mize 2015), mean intensity of infestation of burrows ranged from 0.0 to 14.6 total fleas (0.0–9.5 O. tuberculata and 0.0–14.6 O. hirsuta; Mize 2015), and prevalence of fleas in burrows ranged from 0.0 to 73.3% (0.0–70.0% O. tuberculata and 0.0–43.8% O. hirsuta; Table 2; Mize 2015).

Seasonal changes in abundance between O. tuberculata (black) and O. hirsuta (grey) in black-tailed prairie dog burrows at five national park units. Seasonal changes in prevalence of Y. pestis in O. tuberculata (circles) and O. hirsuta (squares). Numbers above bars are the total number of O. tuberculata and O. hirsuta observed during each month. Numbers below bars represent the total number of burrows swabbed for fleas during each month. Data were pooled from 2009 through 2011; there were no surveys in September. (*) One O. tuberculata flea was examined for Y. pestis and tested positive.
NP, National Park; NM, National Monument; NG, National Grassland.
BADL, Badlands National Park; BUGA, Buffalo Gap National Grassland; DETO, Devils Tower National Monument; SCBL, Scotts Bluff National Monument; THRO, Theodore Roosevelt National Park; WICA, Wind Cave National Park.
A total of 3117 fleas collected from 1053 burrows from 2009 through 2011 were assayed for plague. Of those fleas, 200 tested positive for Y. pestis with a minimum overall prevalence of Y. pestis–positive fleas of 6.4% (Mize 2015). Across colonies, the number of Y. pestis–positive fleas ranged from 0 to 21 total fleas (0–5 O. tuberculata and 0–21 O. hirsuta; Fig. 2, Table 3; Mize 2015) and prevalence of Y. pestis–positive fleas ranged from 0.0 to 100% (0.0–100% O. tuberculata and 0.0–53.8% O. hirsuta; Fig. 2, Table 3; Mize 2015). Overall prevalence of burrows with Y. pestis–positive fleas was 13.9%. The number of burrows on a colony with Y. pestis–positive fleas ranged from 0 to 12 burrows (0–6 O. tuberculata and 0–12 O. hirsuta; Mize 2015) and prevalence of burrows on colonies with Y. pestis–positive fleas ranged from 0.0 to 100% (0–100% O. tuberculata and 0–100% O. hirsuta; Table 3; Mize 2015).
BADL, Badlands National Park; BUGA, Buffalo Gap National Grassland; DETO, Devils Tower National Monument; SCBL, Scotts Bluff National Monument; THRO, Theodore Roosevelt National Park; WICA, Wind Cave National Park.
The coefficient of aggregation for fleas in burrows was variable across colonies, years, and species, with k ranging from 0.04 to 0.96 and
Discussion
Our observed Y. pestis–positive flea prevalence was similar to previous estimates for O. tuberculata (4.37%); however, O. hirsuta prevalence was two times higher than estimates (4.9%) for fleas collected from black-tailed prairie dog colonies in Colorado (Wilder et al. 2008a, b). Several other studies detected Y. pestis in fleas from burrows on black-tailed prairie dog colonies, with burrow prevalence ranging up to 44.4% in Montana (Hanson et al. 2007, Jones et al. 2011) and 19% in Colorado (St. Romain et al. 2013). These studies examined colonies within the same complex or nearby complexes. Our study is the first to examine fleas from colonies across a large geographic range. This may be in part why our range of prevalence of Y. pestis in burrows is larger than previous studies. In addition, our sampling protocol may not have been timed to catch the peak of O. tuberculata abundance in prairie dog burrows. Accordingly, our observed prevalence of Y. pestis–positive O. tuberculata should be considered a conservative estimate of Y. pestis prevalence at our study locations.
We provide evidence of enzootic plague across a large geographic area east of the naturalized plague zone. Considering that sylvatic plague has been documented as far north as Canada (Antonation et al. 2014) and as far south as Texas (Cully et al. 2010, Johnson et al. 2011, Rocke et al. 2012), the results from this study suggest Y. pestis is endemic across the range of black-tailed prairie dogs.
During this study, we observed either enzootic plague, population declines, or epizootic plague on prairie dogs colonies at all of the park units except for Devils Tower NM (Mize 2015), where we never detected Y. pestis. There are several possible reasons for a lack of detection of Y. pestis at Devils Tower NM: (1) Low sample size, (2) the sole prairie dog colony at Devils Tower NM is one of the most isolated in the study and this relative isolation may provide some protection against the arrival of Y. pestis, or (3) Y. pestis may have been suppressed by climatic conditions (Adjemian et al. 2007, Stenseth et al. 2006, Snäll et al. 2008).
Findings from the Wind Cave NP Bison Flats colony are of interest because fleas were sampled within a refugium for western tiger salamanders (Ambystoma mavortium) established prior to 2009. While the surrounding colony was treated with deltamethrin to control fleas, the burrows within the enclosure were not dusted, in part to assess the effects of the insecticide on the salamander. Y. pestis burrow prevalence declined steadily within the enclosure from 2009 through 2011 until dusting was curtailed outside the enclosure in 2012, after which, the Bison Flats colony boundary decreased in spatial extent and the remaining colony appeared less continuous. This is probably due in part to the movement of Y. pestis–positive fleas into the area after dusting.
The untreated enclosure is similar to “refugia” recommended in integrated pest management strategies to maintain gene flow between susceptible naïve populations and those exposed to insecticides, potentially leading to genetically acquired resistance (Coles and Dryden 2014, Halos et al. 2014). Further investigation is warranted to determine if such a dusting regime is an effective plague mitigation strategy.
We observed O. tuberculata and O. hirsuta, both competent vectors of Y. pestis (Wilder et al. 2008a, b), from burrow swabs at each sampled national park unit, as expected from previous studies (Lewis 2002, Brinkerhoff et al. 2006, Salkeld and Stapp 2008, Jones and Britten 2010, Jones et al. 2011). We observed a shift in the flea community on black-tailed prairie dog colonies from O. tuberculata to O. hirsuta in late May to early June. This is consistent with previous work describing the peak abundance of O. tuberculata in late winter to spring and of O. hirsuta in late spring through fall (Salkeld and Stapp 2008, Wilder et al. 2008b, Friggens et al. 2010). This transition in abundance between species may correspond to peaks in plague activity during spring and again in late summer continuing into fall, which we and Wilder et al. (2008a, b) observed. Given this seasonality, current deltamethrin application procedures mostly affect O. hirsuta and may not provide effective coverage of O. tuberculata at its peak abundance in May. We recommend shifting dusting activities, weather permitting, to May or even as early as April to control both flea species. Such action may aid in reducing early transmission of Y. pestis between prairie dogs as they are emerging from burrows in the spring and prevent epizootics from occurring before park personnel can observe population declines.
The preservation of the black-tailed prairie dog, and the prairie community it helps maintain, depends on our ability to detect this exotic pathogen early and to understand how it persists and spreads in the environment. We recommend routine surveillance of prairie dog colonies (e.g., densities, molecular assays, etc.) as a means of early detection. Vigilant monitoring of prairie dog colonies at regular intervals throughout the year is probably the most cost-effective way to identify colonies in the early stages of plague epizootics because it can be used by managers to target plague mitigation strategies more efficiently (Sidle et al. 2001, Cully et al. 2010, McDonald et al. 2011). Plague activity should trigger management activities in colonies near those colonies experiencing epizootics because these are more likely to die off (Stapp et al. 2004, Collinge et al. 2005, Cully et al. 2010). We note that Wind Cave NP and Badlands NP effectively used our yearly Y. pestis prevalence data to help inform their colony dusting efforts over the 3 years of our study. A fairly aggressive dusting program took place during our study, eventually including six of our nine study colonies at Wind Cave NP and four of 10 study colonies at Badlands NP. We attribute the continued existence of many of the treated colonies on these two national park units to early detection of Y. pestis and prompt management action.
With the spread of Y. pestis into the emergence zone in an enzootic state, a proactive approach is recommended (Griebel 2009, Cully et al. 2010, McDonald et al. 2011). History has shown that Y. pestis is almost certainly capable of dispersing east of the distribution of black-tailed prairie dogs. Monitoring should be continued or implemented to track expansion of plague eastward, both on prairie dog colonies where plague has not historically occurred and beyond the distribution of prairie dogs into new species.
Footnotes
Acknowledgments
We wish to thank the National Park Service Natural Resource Preservation Project for providing funding for this study. Additional resources were provided by The University of South Dakota. We would also like to thank National Park Service personnel from each of the study locations, in particular: J. Delger from Badlands National Park; M. Biel, R. Ohms, and A. Wetz from Devils Tower National Monument; R. Manasek from Scotts Bluff National Monument; M. Oehler and C. Sexton from Theodore Roosevelt National Park; B. Muenchau, D. Roddy, and G. Schroeder from Wind Cave National Park; D. Licht from Midwest Regional Office; and R. Griebel from Buffalo Gap National Grassland. We are especially grateful to the many field and laboratory technicians who helped collect and process thousands of fleas. This manuscript has been greatly improved thanks to comments from D. Biggins, M. Dixon, and two anonymous reviewers.
Author Disclosure Statement
No competing financial interests exist.