
Abstract
Severe fever with thrombocytopenia syndrome (SFTS) is an emerging tick-borne infectious disease that is endemic to China, Japan, and the Republic of Korea (ROK). In this study, 8313 ticks collected from SFTS outbreak areas in the ROK in 2013 were used to detect the SFTS virus (SFTSV). A single SFTSV was isolated in cell culture from one pool of Haemaphysalis longicornis ticks collected from Samcheok-si, Gangwon Province, in the ROK. Phylogenetic analysis showed that the SFTSV isolate was clustered with the SFTSV strain from Japan, which was isolated from humans. To the best of our knowledge, this is the first isolation in the world of SFTSV in ticks collected from vegetation.
Introduction
S
Although human-to-human transmission of SFTSV through contact with an infected patient's blood or mucus has been reported (Liu et al. 2012, Gai et al. 2012, Kim et al. 2015), the virus is transmitted to humans predominantly by tick bites. The SFTSV has been previously detected in tick species, including Haemaphysalis longicornis and Rhipicephalus microplus (Yu et al. 2011, Zhang et al. 2012). In our study on SFTSV in ticks collected from humans in the ROK, Amblyomma testudinarium and Ixodes nipponensis ticks were also implicated as potential SFTSV vectors (Yun et al. 2014). Previous studies regarding SFTSV show that domestic animals, such as goats, cattle, dogs, and chickens, and small mammals, such as rodents and shrews, can act as hosts of SFTSV (Jiao et al. 2012, Zhao et al. 2012, Niu et al. 2013, Cui et al. 2013, Liu et al. 2014).
In the ROK, an SFTS case had not been reported prior to a fatal case in 2012 that was identified in Gangwon Province (Kim et al. 2013). Since then, a total of 36 cases, with a mortality rate of 47.2%, has been reported in several regions in the ROK, and 26 virus isolates were obtained from the serum of SFTS patients in 2013 (Park et al. 2014a). Our previous studies reported the existence and prevalence of SFTSV in ticks from the ROK (Yun et al. 2014, Park et al. 2014b). These results showed evidence that SFTS is endemic to several regions of the ROK. However, no studies have focused on the detection of SFTSV in ticks collected from areas within the ROK in which human cases have occurred. To verify the correlation between SFTSV prevalence in ticks and SFTS outbreaks and to determine the possible vector of SFTSV, we aimed to investigate the prevalence of SFTSV in ticks collected from SFTS outbreak areas in the ROK in 2013 and attempted to isolate SFTSV from positive tick pools.
Materials and Methods
Ticks were collected by the flagging and dragging method or by using dry ice–baited tick traps at 14 sites from vegetation in six provinces and two metropolitan cities in the ROK in 2013 (Fig. 1). These sites were selected for the survey based on the number of SFTS cases identified in humans in these regions. Ticks were collected at or near the residences of human cases of SFTS. After collection, the ticks were placed in plastic tubes and transported to the laboratory to identify the species and developmental stage under a dissecting microscope according to a reported classification method (Yamaguti et al. 1971).
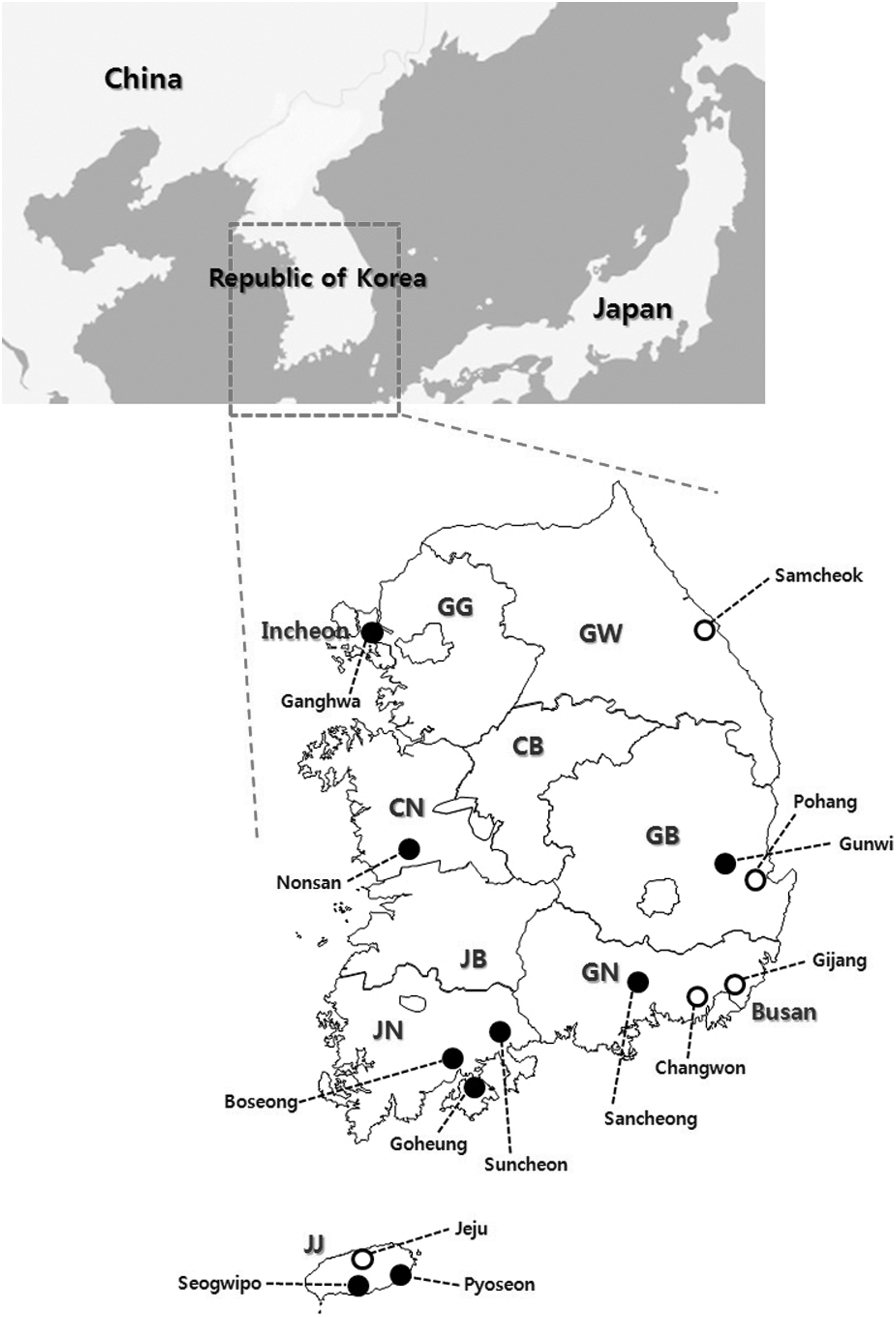
Geographical location of collection sites in this study. Open circles show the sites in which positive tick pools for SFTSV were detected. GG, Gyeonggi Province; GW, Gangwon Province; CB, Chungcheongbuk Province; CN, Chungcheongnam Province; JB, Jeonllabuk Province; JN, Jeonllanam Province; GB, Gyeongsangbuk Province; GN, Gyeongsangnam Province; JJ, Jeju Special Autonomous Province.
Tick samples were pooled according to species, developmental stage, and site (1–50 larvae per pool, 1–30 nymphs per pool, and 1–5 adults per pool). The pooled ticks were homogenized in 600 μL of phosphate-buffered saline (PBS; pH 7.0) containing 10% fetal bovine serum (FBS; Gibco BRL, Grand Island, NY), penicillin (500 IU/mL, Gibco BRL), and streptomycin (500 μg/mL, Gibco BRL) using a Precellys® 24 homogenizer (Bertin Technologies, Bretonneux, France) and 2.8-mm stainless steel beads. The tick homogenates were centrifuged at 8600 × g for 5 min, and RNA was extracted from the supernatants of the tick homogenates according to the manufacturer's instructions using a Viral Gene-spin™ RNA Extraction Kit (iNtRON Biotechnology, Seongnam, ROK). To detect the SFTSV M segment gene in tick homogenates, one-step reverse transcription polymerase chain reaction (RT-PCR) was conducted using a DiaStar 2X OneStep RT-PCR Premix Kit (SolGent, Daejeon, ROK) via a previously described method (Yun et al. 2014). The RT-PCR amplicon was purified using a QIAquick® Gel Extraction Kit (QIAGEN, Valencia, CA) according to the manufacturer's instructions and sequenced after cloning into a pCR®2.1-TOPO® plasmid (Invitrogen, Carlsbad, CA) using an ABI Prism BigDye Terminator Cycle Sequencing Kit and an ABI 3730xl Sequencer (Applied Biosystems, Foster City, CA) at Macrogen Inc. (Daejeon, ROK).
To isolate SFTSV from positive tick pools, tick homogenates were added to an equal volume of Dulbecco's Modified Eagle Medium (DMEM; Gibco BRL) containing penicillin (5000 IU/mL, Gibco BRL), and streptomycin (5 mg/mL, Gibco BRL). The mixture was filtered using a 0.45-μm syringe filter and clarified by centrifugation, and the supernatants were used as the inoculums for virus isolation. Vero E6 cell monolayers formed in 12-well plates were inoculated with the supernatants and incubated at 37°C for 7 days. Then, cell suspensions were passaged to new monolayers of Vero E6 cells, as previously described (Park et al. 2014a), and stored at −70°C until use.
To identify the isolated virus, we examined the presence of the SFTSV M segment gene in passaged cell supernatants using RT-PCR and confirmed the results by indirect immunofluorescent antibody assay (IFA), immunoblotting using a monoclonal SFTSV nucleocapsid (N) protein antibody (manufactured by our laboratory), and transmission electron microscopy (TEM) analysis for morphological identification.
For phylogenetic analysis, sequence alignment and construction of the phylogenetic tree were performed using MEGA software version 6.0 (Tamura et al. 2013).
Results and Discussion
A total of 8313 ticks belonging to three genera and four species (H. longicornis, Haemaphysalis flava, I. nipponensis, and A. testudinarium) were collected. Of the identified ticks, H. longicornis (n = 8230; 99%) was the most frequently collected species in this study, followed by H. flava (n = 73; 0.88%), I. nipponensis (n = 9; 0.11%), and A. testudinarium (n = 1; 0.01%). All ticks collected in this study had not fed. Among them, SFTSV RNA was detected in nine pools (eight pools in H. longicornis and one pool in H. flava) (Table 1). The SFTSV minimum infection rate per 100 ticks (minimum infection rate [MIR] = no. of positive pools/no. of examined ticks in pools ×100) was 0.11% (nine pools/8313 individuals). These results suggest that H. longicornis and H. flava, from which SFTSV genome was detected, serve as potential vectors in the ROK, although H. flava has not been previously considered as a SFTSV vector. Additionally, SFTSV RNA was detected in H. longicornis larva, suggesting that SFTSV may possibly be transmitted by the transovarial route. To support this hypothesis, SFTSV will need to be isolated from infected ticks (potential vectors) and the virus characterized further. Laboratory vector competence studies are needed to confirm the transovarial transmission of SFTSV in ticks.
L, larvae; N, nymphs; M, adult males; F, adult females.
MIR = Minimum infection rate per 100 ticks (no. of positive pools/no. of examined ticks in pools ×100).
−, Negative result in the virus isolation test.
Of the nine SFTSV-positive tick pools, only one pool of H. longicornis nymphs collected from Samcheok-si, Gangwon Province, was positive in the virus isolation test. As shown in Figure 2, the virus isolate designated as KAGWT from H. longicornis was identified as a SFTSV. The sequence of KAGWT determined in this study has been deposited in GenBank (acc. no. KP777541).
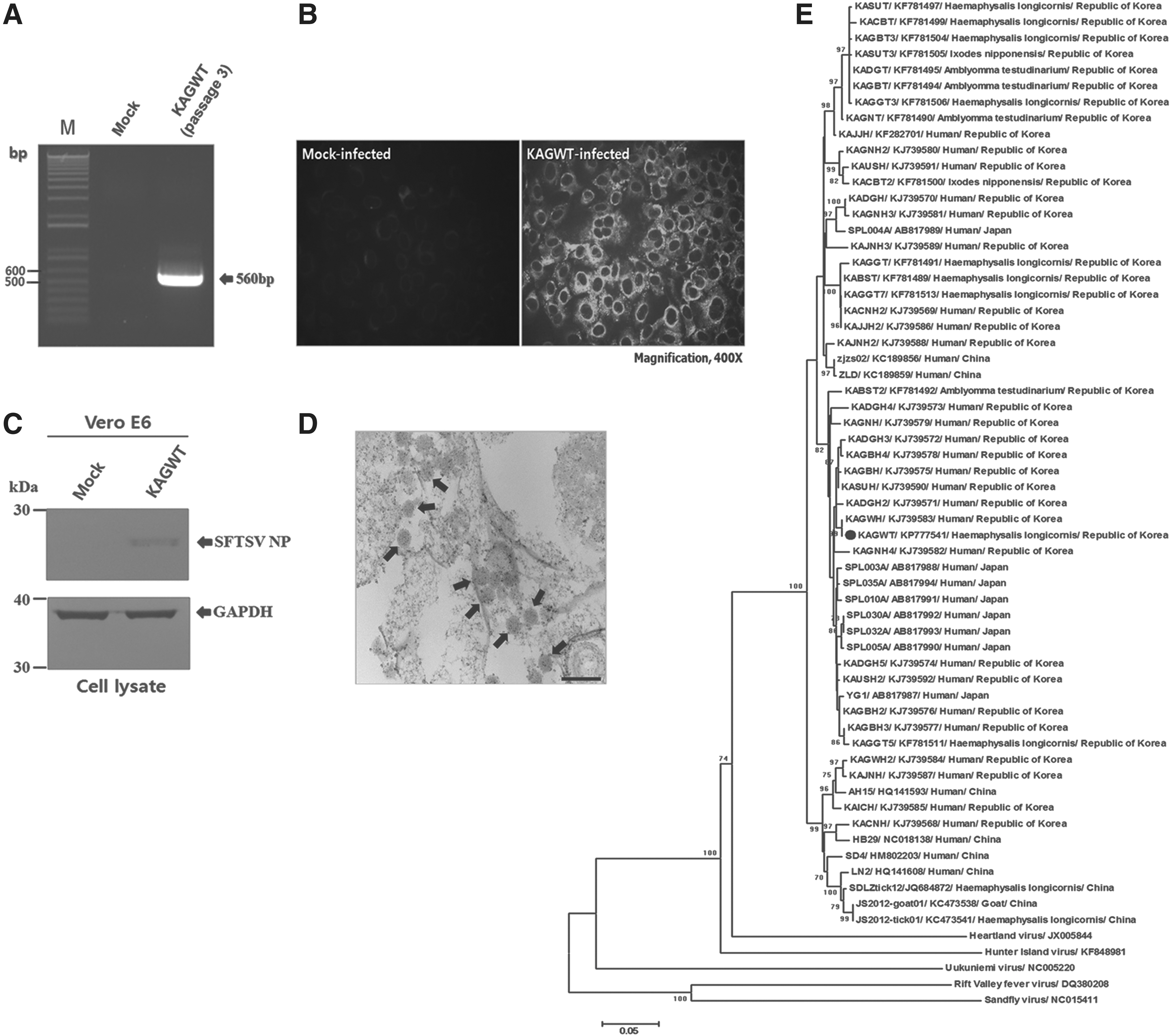
Identification of severe fever with thrombocytopenia syndrome virus (SFTSV) isolated from ticks and phylogenetic analysis of SFTSV strains. (
The partial M segment genome-based phylogenetic analysis revealed that the SFTSV KAGWT was more closely related to the SFTSV strain from Japan, which was isolated from humans. Our result is consistent with a previous report (Park et al. 2014a) indicating that SFTSV Korean isolates from humans are closely related to Japanese isolates. To our knowledge, this is the first survey of SFTSV prevalence in ticks collected from SFTS outbreak areas of the ROK, and this article describes studies that identify the first isolation of SFTSV from ticks collected in the ROK.
In conclusion, it was demonstrated that SFTSV-infected ticks were distributed in several areas of the ROK in which an SFTS outbreaks occurred in 2013. Furthermore, SFTSV was first isolated from H. longicornis ticks collected in the study area in which the SFTS human case occurred. This suggests that H. longicornis is a vector for SFTSV in the ROK. Although virus isolation from H. flava failed, we suspect that this tick may play a role in SFTSV transmission. These findings emphasize the need for continuous tick-based surveillance for SFTSV to monitor information about SFTSV activity and distribution in the ROK. Taken together, further research, such as whole genome analysis of a tick-derived SFTSV Korean isolate, is needed to understand better the genetic diversity and molecular evolution of SFTSV.
Footnotes
Acknowledgments
The authors thank Ki Ju Choi for technical support with transmission electron microscopy analysis and Dr. Sun-Whan Park for providing us with the SFTSV monoclonal antibody used in this study. This research was funded by an intramural grant of the Korea National Institute of Health (grant no. 2014-ND53001-00).
Author Disclosure Statement
No competing financial interests exist.