
Abstract
This study aimed to investigate the prevalence and genetic diversity of Bartonella organisms in small mammals in Cotonou, Benin. We captured 163 rodents and 12 insectivores and successfully detected Bartonella DNA from 13 of the 175 small mammal individuals. Bartonella spp., identical or closely related to Bartonella elizabethae, Bartonella tribocorum, and Bartonella rochalimae, was detected. A potential new Bartonella species, proposed as Candidatus Bartonella mastomydis, was found in three Mastomys individuals and genetically characterized by targeting two housekeeping genes (rpoB and gltA) and the intergenic species region. However, 20.8% of gray rats were found to be infected with Bartonella spp., and none of the black rats analyzed was positive. This work may be important from a public health point of view due to the zoonotic nature of the Bartonella species detected and warrants further investigation on the unknown zoonotic potential of this newly proposed Bartonella species.
Introduction
T
Although a recent study showed that the overall prevalence of Bartonella sp. in Xenopsylla cheopis fleas in Benin was 34.67% (Leulmi et al. 2014), no study was conducted in small mammals to identify Bartonella reservoirs. Consequently, our aim was to analyze the prevalence of Bartonella infections in small mammal populations of three wide areas in Cotonou, where half of the nation's population (over 3 million people) resides (Dossou and Glehouenou-Dossou 2007).
To estimate the zoonotic risk and evaluate the genetic diversity associated with these Bartonella organisms, genotypes of Bartonella spp. were determined using the citrate synthase gene (gltA), which is especially popular and a widely used target for distinguishing closely related Bartonella species and genotypes (Kosoy et al. 2012), RNA polymerase beta-subunit (rpoB), and 16S-23S rRNA intergenic species region (ITS). Evaluating the genetic heterogeneity of the Bartonella strains in small mammals in Benin will also serve to increase the knowledge about Bartonella host specificity.
Materials and Methods
A total of 163 rodents and 12 insectivores were captured in three zones of Cotonou, Republic of Benin (Fig. 1). Animals were trapped inside human residences and peridomestic areas between November 2009 and July 2010, as Houemenou et al. (2013) described previously. Rodent individuals belonged to the species Rattus rattus (black rats), Rattus norvegicus (gray rats), Mastomys sp., and Mus sp., whereas Crocidura sp. and Crocidura olivieri were the insectivore species included in this study (Table 1). The Xenopsylla cheopis fleas harbored by some of these small mammal individuals that had been already screened for the presence of Bartonella sp. (Leulmi et al. 2014).
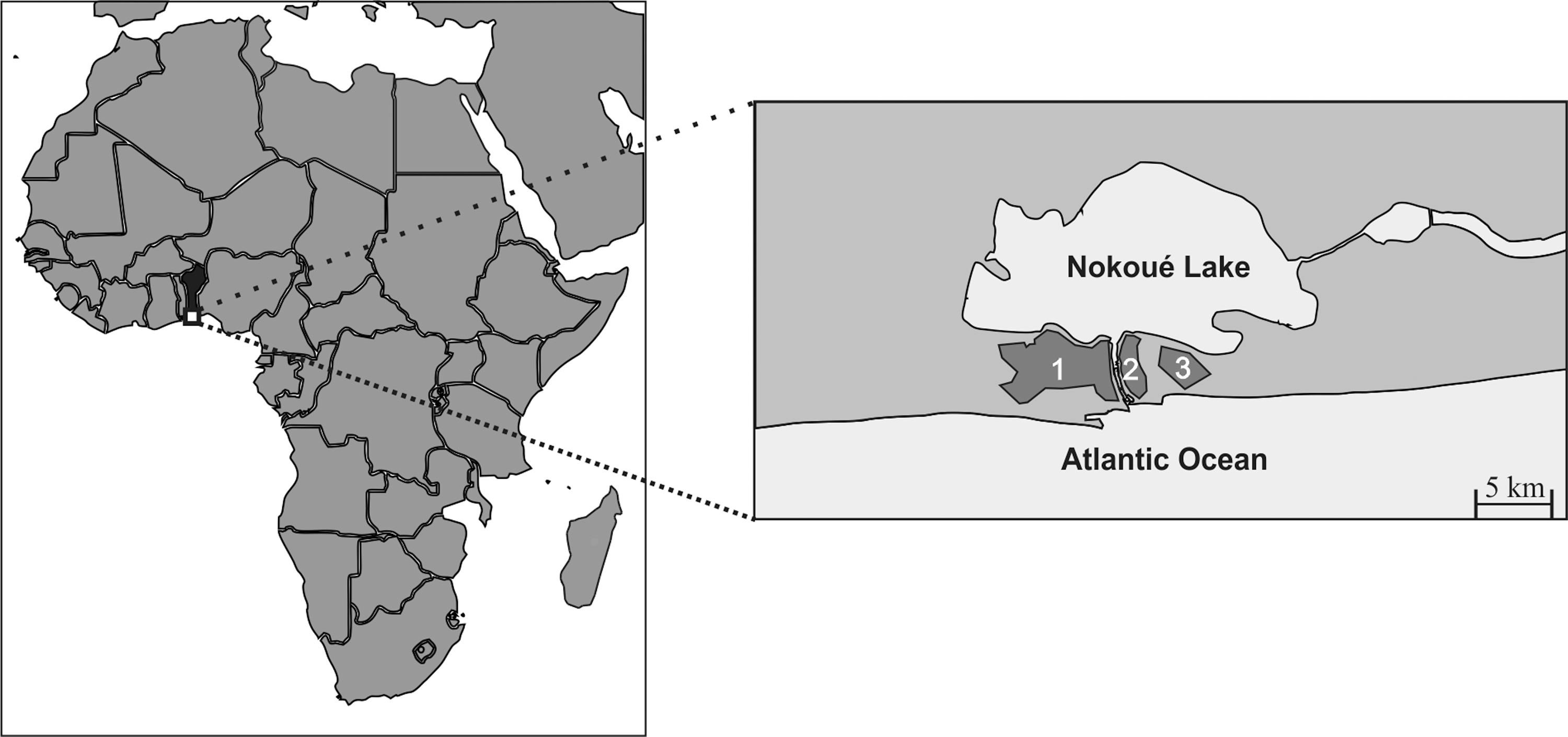
Sampling areas in Cotonou, Benin.
Spleen samples were collected from the small mammals Rattus rattus, Rattus norvegicus, Mastomys sp., Crocidura olivieri, and Crocidura sp. A total of 175 small mammals were examined for the presence of Bartonella species in three zones in Cotonou, Benin.
N, number of small mammals analyzed; +, number of positive samples; P, prevalence.
Spleens of animals were obtained, and DNA was isolated by using the Illustra Tissue and Cells Genomic Prep Mini Spin Kit (GE Healthcare, Little Chalfont, UK). The presence of Bartonella was examined by PCR targeting the gltA gene as Billeter et al. (2011) described previously. This amplicon encompasses a 327-bp-specific zone that has been described to be useful for distinguishing Bartonella species (La Scola et al. 2003). In case of a positive PCR result, we performed a second PCR targeting the rpoB gene, as Renesto et al. (2001) described previously. The ITS was also sequenced if the homology of both gltA and rpoB genes with the rest of described Bartonella species suggested the existence of an undescribed Bartonella species. The ITS was amplified according to Roux and Raoult (1995).
Those positive rodent individuals identified morphologically only at the genus level were genotyped by amplification and sequencing the nuclear interphotoreceptor binding protein gene (IRBP), as Weksler (2003) described previously. This nuclear gene has been used extensively to address questions of mammalian interordinal phylogeny (Stanhope et al. 1996, Springer et al. 1999) and to discern relationships at lower taxonomic levels as well (Yoder and Irwin 1999, Jansa and Voss 2000).
Sequencing reactions were performed for both strands at Macrogen, Inc. In the case of suspecting mixed infections due to the presence of clear double peaks in the chromatograms, the amplified products were cloned into the pGEM-T Easy cloning vector (Promega) for further sequencing using plasmid primers SP6 and T7. New and previously published Bartonella sequences, which were retrieved from GenBank, were aligned with the multiple alignment program ClustalW in MEGA 6.06 (Tamura et al. 2013). Sequences were compared by using the nucleotide–nucleotide BLAST (blastn) program.
Phylogenetic relationships of the Bartonella spp. genotypes based on partial sequences of the gltA and rpoB genes were assessed by the neighbor-joining method with Kimura's two-parameter distance model (Kimura 1980). Bootstrap analysis was performed with 1000 trials. In the case of suspecting the presence of an undescribed Bartonella species, a phylogenetic tree was constructed from concatenated sequences of gltA and rpoB genes and the ITS.
Chi-square contingency tables were used to compare the proportion of Bartonella-infected small mammals between the different host species and areas included in the study. Fisher's exact tests were used if expected cell counts were less than five. A probability value less than <0.05 was considered as statistically significant.
This study was approved by the Ethical Committee of l'Université d'Abomey-Calavi of Benin, reference no. 208/MCOT/SG/DRH/DDCPRS/SFERM.
Results
Thirteen out of the 175 small mammals analyzed were positive for Bartonella species. Therefore, the overall prevalence was 7.4%. Bartonella DNA was found in R. norvegicus, Mastomys sp., and C. olivieri. The highest Bartonella prevalences were found in Mastomys sp. (25.9%) and R. norvegicus (20.8%), whereas none of the black rats analyzed (n = 110) were found to be harboring Bartonella organisms. As a result, Bartonella prevalence was significantly higher in both Mastomys sp. and R. norvegicus than in R. rattus (p < 0.001 Fisher's exact test for both host comparisons). Bartonella-positive mammals belonged to the three areas studied; although the highest prevalence was observed in area 2 (9.1%), no significant differences of Bartonella prevalence were found among the three areas included in the study.
Seven Mastomys sp. individuals were positive for Bartonella spp., and five of them were identified as Mastomys natalensis on the basis of their IRBP gene sequences, which were submitted to the GenBank under the following Accession numbers: KP862858–KP862860 and 1863188.
Five gltA genotypes were detected and designated as genotypes g-A to g-E. The phylogenetic relationships between the isolates obtained in this study and previously described Bartonella species are shown in Figure 2, and additional information about these gltA genotypes is supplied in Table 2. The genotypes g-A and g-B showed 99% and 100% identity, respectively, with GenBank deposited B. elizabethae sequence (GenBank Accession number JX158351). The genotype g-C had 100% similarity to the GenBank deposited B. tribocorum sequence (GenBank Accession number JX158361), and the genotype g-D showed 100% identity with B. rochalimae (GenBank Accession number FN645459). Finally, the genotype g-E showed 94% BLAST similarity with the closest gltA sequence deposited in the GenBank database (GenBank Accession number AY584853), which corresponded to Bartonella taylorii isolated from Apodemus agrarius (Mediannikov et al. 2005).

Phylogenetic classification of Bartonella genotypes based on gltA gene sequences. Only the bootstrap values above 70% obtained are given. The GenBank Accession numbers for reference sequences are given between brackets. Isolates sequenced in this study are indicated in bold. Sequences obtained from coinfecting Bartonella organisms isolated from the same host individual were designated with a number after a dash. Scale bar indicates 20 substitutions per nucleotide position. The gltA sequence of Bartonella tamiae was included as the out-group.
A total of 175 small mammals were screened for the presence of Bartonella spp. by using citrate synthase gene (gltA) and RNA polymerase beta-subunit (rpoB) primers; 13 individuals harbored Bartonella organisms. Five gltA genotypes and five rpoB genotypes were observed. Similarities below 95% for both genes suggested the existence of a new Bartonella species. Dashes indicate that the corresponding isolate could not be sequenced. Mastomys individuals were genetically characterized by targeting the interphotoreceptor binding protein gene (IRBP) gene.
During genetic analyses, clear double peaks (distinct from the baseline noise) were visible at species-specific positions on the sequencing chromatograms of the gltA products corresponding to three isolates, Ben165, Ben178, and Ben205, suggesting mixed infections. Among them, only the isolate Ben205 was cloned successfully, allowing us to distinguish two different gltA genotypes, g-A and g-E. Selected Bartonella gltA sequences were deposited in GenBank under the following accession numbers: KP857582–KP857587, KR107234, KT881099, and KT881100.
With regard to the rpoB gene, five partial rpoB genotypes were also obtained and designated as genotypes r-A to r-E. The phylogenetic relationships between the isolates obtained in this study and previously described Bartonella species according to rpoB sequences analysis are shown in Figure 3, and additional information is also depicted in Table 2. The genotypes r-A to r-C of rpoB gene were highly similar to B. elizabethae (98–99% BLAST similarity). The isolates Ben9 and Ben28, which had shown 100% similarity to B. tribocorum and had been consequently classified as genotype g-C when analyzing the gltA gene, were grouped together with B. elizabethae (98–99% BLAST similarity) on the basis of the rpoB genotype. On the contrary, the genotype r-D was concordant with the genotype g-D of the gltA gene and showed 97% similarity with B. rochalimae.
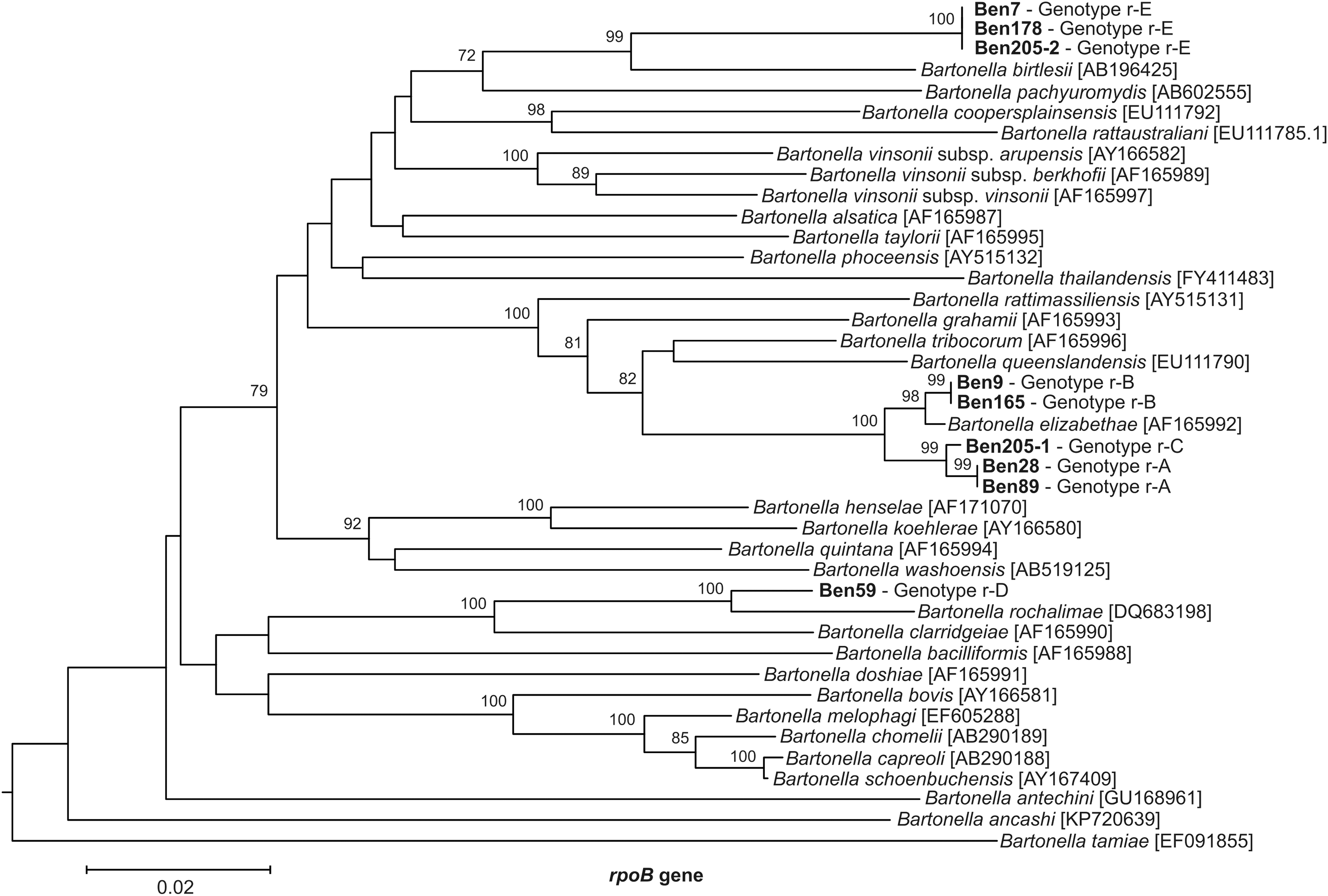
Phylogenetic classification of Bartonella genotypes based on rpoB gene sequences. Only the bootstrap values above 70% obtained are given. The GenBank Accession numbers for reference sequences are given between brackets. Isolates sequenced in this study are indicated in bold. Sequences obtained from coinfecting Bartonella organisms isolated from the same host individual were designated with a number after a dash. Scale bar indicates 20 substitutions per nucleotide position. The rpoB sequence of Bartonella tamiae was included as the out-group.
Finally, the genotype r-E of rpoB, which corresponds to the genotype g-E of the gltA gene, showed the highest similarity (99%) with an undescribed Bartonella organism that had been previously found in Arvicanthis dembeensis and Mastomys awashensis in Ethiopia (GenBank Accession number JQ425643 and JQ425641, respectively) (Meheretu et al. 2013). This genotype r-E showed 94% BLAST similarity with the closest, related described Bartonella species, B. birtlesii. Consequently, since partial sequences of both gltA and rpoB genes of isolates Ben7 and Ben205-2 (corresponding to genotypes g-E and r-E) were less than 95% similar to the rest of described Bartonella species, fulfilling the species description cutoff designated by La Scola et al. (2003), it is likely that this strain belongs to a new Bartonella species.
According to the isolate Ben205, double peaks were also found at species-specific positions, suggesting again the presence of two Bartonella strains. Cloning these rpoB PCR products allowed us to confirm the existence of two coinfecting Bartonella organisms, whose genotypes were classified as r-C and r-E. Bartonella rpoB sequences were deposited in GenBank under the following accession numbers: KP997017–KP997022 and KT881101–KT881103 (Table 2).
Those Bartonella organisms whose gltA and rpoB genotypes were less than 95% similar to the rest of described Bartonella species were further analyzed for the ITS, and the homology was 91% or less between our isolates and known Bartonella species (GenBank Accession number KT895985). The highest BLAST similarities for the ITS, along with gltA and rpoB genes, are given in Table 3. The phylogenetic tree of the concatenated sequences of gltA and rpoB genes and ITS showed that this undescribed Bartonella species aligned most closely to B. birtlesii, and both Bartonella organisms were grouped separately in a well-supported clade (99% bootstrap support; Fig. 4), as was observed in the rpoB phylogenetic tree (Fig. 3).
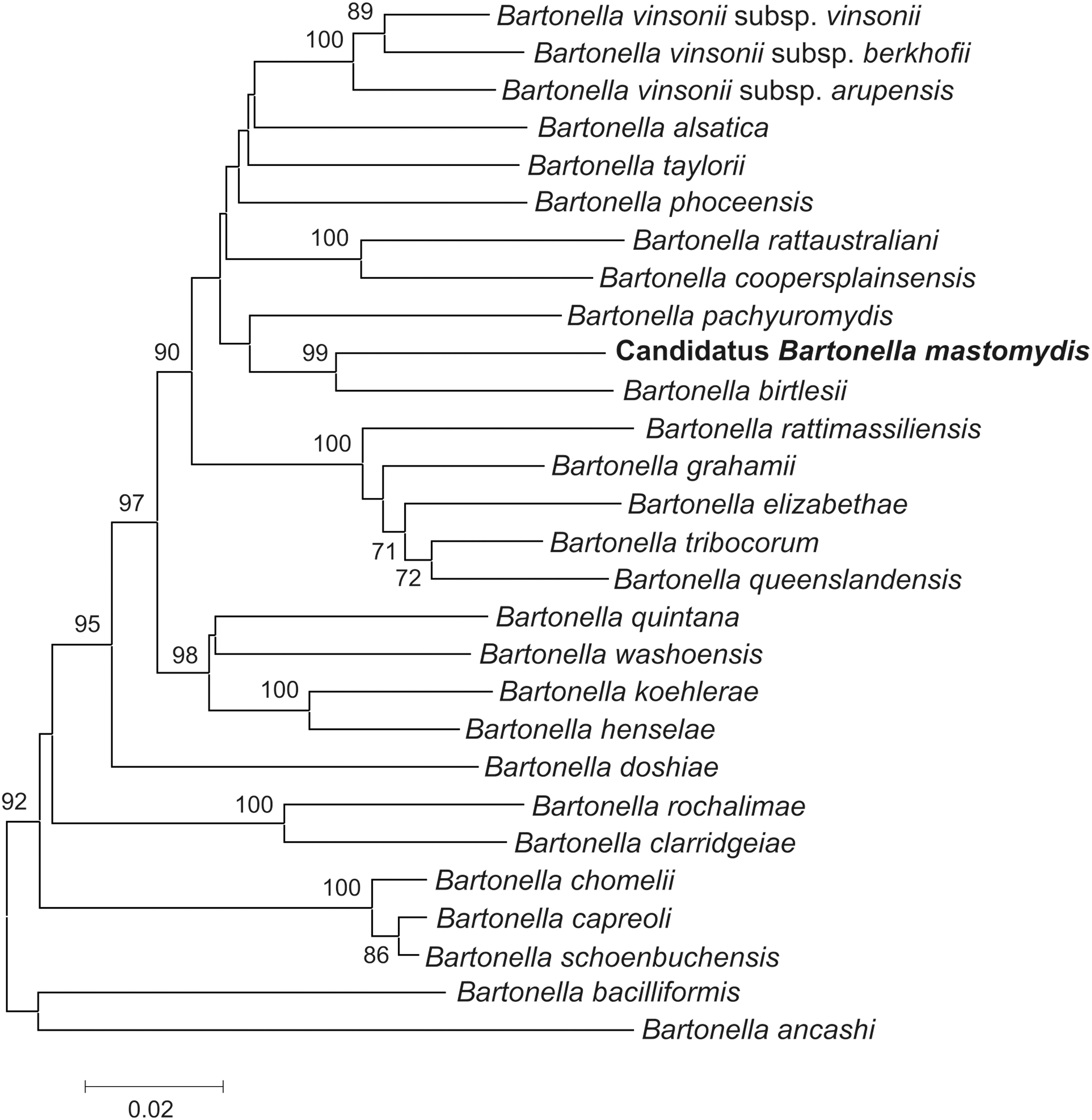
Phylogenetic tree of Candidatus Bartonella mastomydis and other Bartonella species. The phylogenetic tree was based on concatenated sequences of gltA and rpoB genes and the 16S-23S rRNA intergenic spacer region and was constructed from 1378 base pair sequences, inferred using the Neighbor-Joining method, and 1000 replicates in the bootstrap test. Scale bar indicates 20 substitutions per nucleotide position. The newly proposed Bartonella species is indicated in bold letters. Only those Bartonella species for which gltA, rpoB, and intergenic species region sequences of the desired length were available at the GenBank database were included in this phylogeny.
Percentage and GenBank Accession numbers.
ITS, intergenic species region.
Discussion
The results of this study indicate that Bartonella organisms are widely distributed in Cotonou, as they were found in all the areas included in this study, which cover ∼14 km in length.
To our knowledge, this work constitutes the first finding of Bartonella spp. in rodents in Benin. The overall prevalence observed in this study (7.4%) was clearly influenced by the absence of Bartonella spp. in black rats (n = 110).
The remarkable prevalence found in R. norvegicus and Mastomys sp. could indicate that Bartonella infections are common in these rodent species.
The discovery of five gltA and five rpoB genotypes in this restricted sampling area indicates a high level of Bartonella diversity among small mammals in Cotonou. The finding of B. elizabethae in small mammals in Cotonou increases the already widespread distribution of this zoonotic species throughout Africa, as it has been already found in countries as geographically distant as the Democratic Republic of Congo (DRC), Tanzania (Gundi et al. 2012), Uganda (Billeter et al. 2014), South Africa (Brettschneider et al. 2012b), or Ethiopia (Meheretu et al. 2013). Almost the same could be said for B. tribocorum, as it has also been already isolated in many African countries, such as Nigeria (Kamani et al. 2013), South Africa (Pretorius et al. 2004), DRC, or Tanzania (Gundi et al. 2012).
La Scola et al. (2003) proposed that newly encountered Bartonella isolates should be considered a new species if the 327-bp gltA fragment shares <96.0% sequence similarity with those of the validated species and if an 825-bp rpoB fragment shares <95.4% sequence similarity with those of the validated species. Two isolates analyzed in this study satisfied this species description cutoff for both genes. Therefore, it is likely that this strain belongs to a new Bartonella species. This conclusion was supported by the fact that the analyzed portion of the ITS of this organism showed 91% similarity with the closest Bartonella species, whereas La Scola et al. (2003) demonstrated that the median value of interspecies similarity for the ITS, with regard to those Bartonella species already described at that time, was 93.9%.
Therefore, our study seems to ratify the finding carried out by Meheretu et al. (2013), who indicated that some of the Bartonella strains circulating among Murid rodents in Africa were likely to correspond to undescribed Bartonella species. In this sense, one of those undescribed Bartonella species, designated as clade 5 in their study, corresponds to our newly proposed species.
This Bartonella organism was found in Mastomys sp. individuals in our study, genetically identified as M. natalensis on the basis of their IRBP gene sequences, which were also found to carry B. tribocorum and B. elizabethae organisms in our study. This finding indicates the wide range of Bartonella species for which this rodent species may be acting as a reservoir in Africa. The zoonotic role of M. natalensis is highlighted by the fact that this rodent species is the only known reservoir for Lassa Fever (Lalis et al. 2012). On the contrary, taking into consideration that *272 species of Muridae exist in Africa (37% of the world total) (Musser and Carleton 2005), it seems obvious that many more Bartonella genotypes/species are going to be discovered in this continent.
Since it was not possible to obtain a pure microbiological culture, we propose this Bartonella organism to be named Candidatus Bartonella mastomydis, “Mastomys” being the name of the rodent genus from which the sequence was first obtained. However, to eventually confirm whether this is a new species, further genetic identification by genome sequencing will be required and is currently ongoing.
Some Bartonella species have been found to infect a limited number of rodent species (i.e., B. vinsonii subsp. arupensis and B. washoensis; Kosoy et al. 2003; Inoue et al. 2011, Bai et al. 2011), whereas others such as B. elizabethae or B. queenslandensis have been found to infect many rodent species (Pangjai et al. 2014, Jiyipong et al. 2015). As has been previously observed (Gundi et al. 2010), we found a lack of host specificity for B. elizabethae and B. tribocorum, since similar or identical genotypes associated with these Bartonella organisms were isolated from different small mammal species in our study. This phenomenon could suggest that the transmission of both Bartonella species may occur by shared or common ectoparasites, such as one or more species of flea or tick, found distributed throughout the African continent. In this sense, and taking into consideration the results obtained by Leulmi et al. (2014), X. cheopis fleas are the most likely candidates playing this role in Cotonou.
Although intracellular bacteria have low rates of recombination and horizontal gene acquisition (Vos and Didelot 2009) probably because of their isolated growth niche, different Bartonella species may coexist during their life cycle, probably within their arthropod vectors, providing opportunities for recombination (Buffet et al. 2013). As a consequence, Bartonella strains circulating within rodents have showed more frequent recombination events compared to human and cat adapted species, which may have resulted in a broader host range for rodent adapted species (Berglund et al. 2010; Paziewska et al. 2011). With regard to B. tribocorum, a recent study also demonstrated that its lack of host specificity may have been the result of the action of host adaptability genes that are packaged into bacteriophage particles, promoting rapid diversification and facilitating host shifting (Berglund et al. 2009).
With respect to two isolates (Ben9 and Ben28), they did not show the highest BLAST similarity to the same Bartonella species when comparing gltA and rpoB results. This finding may be due to the elevated number of recombination events that occurs within the gltA gene (Buffet et al. 2013). In this sense, this gltA sequence seems to suggest recombination between B. tribocorum and B. elizabethae, two Bartonella species that were found in other rodent individuals in this study. This recombination event may have probably occurred within the arthropod vector, as X. cheopis fleas have been found to be infected with both Bartonella species (Billeter et al. 2013). Therefore, only using this gene might lead to biases resulting in the false identification of genotypes (Paziewska et al. 2011).
Strikingly, none of the black rats analyzed in this study was found to harbor Bartonella organisms, whereas 20.8% of R. norvegicus individuals were PCR-positive. This result is similar to those obtained by many authors that have indicated that Bartonella prevalence differs markedly in R. norvegicus and R. rattus, being always higher in the former species (Ellis et al. 1999, Hsieh et al. 2010). These prevalence differences are of interest as one would anticipate that related species with similar biology that occurs in the same area would have comparable susceptibility to infection, suggesting that some factor is likely to be responsible for these differences (Brettschneider et al. 2012a).
In accordance with the study carried out by Leulmi et al. (2014), which analyzed the presence of Bartonella sp. in the ectoparasites carried by the same rodent individuals that were screened in this work, Bartonella prevalence was remarkably high in X. cheopis fleas obtained from both rat species (52.6% in gray rats and 34.3% in black rats). The high prevalence of Bartonella sp. in X. cheopis fleas obtained from black rats in that study, taken together with the absence of positive cases in this rodent species in our study, suggests that whatever is the exact nature of this (these) factor (s), it seems to be hampering the transmission of Bartonella spp. organisms from the vector to the host. Supporting this hypothesis, four of the Bartonella-negative black rats analyzed in this study were harboring infected fleas. In this sense, further studies are needed to ascertain the real zoonotic risk associated to black rats in Western Africa.
Our results could be important from a public health point of view due to the pathogenic potential of the species identified, which has been isolated from humans with severe clinical manifestations. In this sense, B. elizabethae has been reported to cause endocarditis (Daly et al. 1993), which is the most common clinical manifestation of Bartonella infection (Renesto et al. 2001), and B. rochalimae has been found in patients with bacteremia, fever, and splenomegaly (Tsai et al. 2010). Similarly, B. tribocorum has been found in a Thailand patient with fever (Kosoy et al. 2010). Therefore, the risk of human infection by Candidatus Bartonella mastomydis and the rest of Bartonella species found in this study should be studied in patients from Benin with fever of unknown origin. The risk of zoonotic transmission is underlined by the peridomestic and intradomestic environment of trapped animals, which suggests that they can come into close contact with humans with its consequent zoonotic risk.
At this point, it must be taken into consideration that Mastomys spp. and many other rodent species constitute the main source of animal food for rural populations in Benin (Assogbadjo et al. 2005). Consequently, transmission to humans is likely to be taking place in rural zones due to contacts with infected animals or through their blood-sucking arthropods. Therefore, this epidemiological study must be extended to rural areas to ascertain the incidence of Bartonella-related diseases in areas where potential Bartonella reservoirs constitute the main source of animal food.
Finally, this zoonotic risk could be exacerbated by the socioeconomic and political constraints affecting Cotonou, especially rapid demographic growth and inadequate resources for urban development, as previously described (Dossou and Glehouenou-Dossou 2007). In that sense, the inadequate waste management found in Cotonou may lead to an increase of rodent populations, a phenomenon that could imply an increase of human–rodent contacts and the number of Bartonella-infected rodents. In conclusion, our results demonstrate the importance of these mammal species as reservoirs for zoonotic Bartonella species and warrant increased awareness of physicians and healthcare workers for these pathogens, especially in unidentified febrile cases.
Footnotes
Acknowledgments
This work was supported by projects Red de Investigación Cooperativa en Enfermedades Tropicales—RICET (RD12/0018/0013), CGL 2009-07759BOS, and 2014SGR 1241 (Generalitat de Catalunya). A.M.-A. was supported by a PhD grant from “Agencia Canaria de Investigación, Innovación y Sociedad de la Información.”
Author Disclosure Statement
No competing financial interests exist.