
Abstract
Ticks of the genus Dermacentor are known vectors of rickettsial pathogens in both the Old World and New World. In North America, Dermacentor variabilis and D. andersoni are vectors of Rickettsia rickettsii, while in Europe, D. marginatus and D. reticulatus transmit R. slovaca and R. raoultii, respectively. Neither the presence of R. slovaca in the Americas nor the ability of American tick species to maintain this pathogen have been reported. Here we describe detection of Rickettsia genetically identical to R. slovaca in D. variabilis, its molecular characterization, assessment of pathogenicity to guinea pigs, and vector competence of D. variabilis ticks. Ticks from a laboratory colony of D. variabilis, established from wild ticks and maintained on naïve NZW rabbits, tested positive for spotted fever group (SFG) Rickettsia by PCR. Analysis of 17 kDa gltA, rpoB, ompA, ompB, and sca4 genes revealed 100% identity to R. slovaca sequences available in the GenBank. New Zealand white rabbits fed upon by infected ticks seroconverted to SFG Rickettsia. Guinea pigs inoculated with the Rickettsia culture or infested by the infected ticks developed antibodies to SFG Rickettsia. The intensity of clinical signs and immune response were dependent on dose and route of infection. The identified Rickettsia was detected in all life stages of D. variabilis ticks, confirming transstadial and transovarial transmission. Thirty-six percent of uninfected larvae co-fed with infected nymphs on guinea pigs were PCR-positive and able to pass rickettsia to at least 11.7% of molted nymphs. To our knowledge, this is a first report of identification of a European pathogen R. slovaca or a highly similar agent in the American dog tick, D. variabilis. Considering pathogenicity of R. slovaca in humans, further laboratory and field studies are warranted to assess the relevance of the above findings to the public health and epidemiology of SFG rickettsioses in the United States.
Introduction
T
Ticks from the genus Dermacentor are known vectors of human diseases in both the Old and New Worlds. In North America, the American dog tick, Dermacentor variabilis (Say), and the Rocky Mountain wood tick, D. andersoni Stiles, are vectors of Rickettsia rickettsii, the causative agent of Rocky Mountain spotted fever. Colorado tick fever is a viral infection associated with D. andersoni at high elevations in Rocky Mountains (Florio and Stewart 1947). In Europe Dermacentor spp. ticks, namely D. marginatus (Sulzer) and D. reticulatus (Fabricius), transmit R. slovaca (Marquez 2006) and R. raoultii (Spitalska 2012). There is no demonstrated overlap between the geographical ranges of the European and North American Dermacentor species or Dermacentor-transmitted pathogens.
Rickettsia slovaca was first isolated from D. marginatus tick collected in central Slovakia in 1968 (Brezina et al. 1969), but was not considered a human pathogen until 1997 when the first human case was described (Raoult et al. 1997). Rickettsia slovaca causes a syndrome characterized by scalp eschars and neck lymphadenopathy and was initially known as TIBOLA (tick-borne lymphadenopathy) or DEBONEL (Dermacentor-borne necrotic erythema and lymphadenopathy) (Ibbara et al. 2006). Recently, the unified term SENLAT (scalp eschar and neck lymph adenopathy after a tick bite) was proposed for this clinical complex (Angelakis et al. 2010). The presence of R. slovaca in Dermacentor ticks and associated human disease have been recorded across Europe (Parola et al. 2009). Rickettsia raoultii also causes a similar illness in humans (Mediannikov et al. 2008), although current studies suggest that R. raoultii is less pathogenic than R. slovaca (Parola et al. 2009).
Natural cycles of tick-borne diseases are complex. The study of transmission, maintenance, infectivity, virulence, and pathogenicity of tick-borne agents requires advanced laboratory techniques and animal models, as well as live vectors (ticks) to reproduce a natural route of infection via a tick bite (Troughton and Levin 2007). Tick-rearing facilities serve those purposes through maintaining colonies of medically important species. Here we report the detection of a Rickettsia sp. genetically identical to R. slovaca in a colony of Dermacentor variabilis originated from wild ticks collected in the Virginia–North Carolina region of the Eastern United States.
Materials and Methods
Infected tick colony
Ticks are maintained in the Medical Entomology Laboratory at Centers for Disease Control (CDC) under standard laboratory conditions as described previously (Troughton and Levin 2007). All work involving animals is done according to protocols approved by the CDC Institutional Animal Care and Use Committee. Naïve New Zealand white (NZW) rabbits (Oryctolagus cuniculus) are used as hosts for all tick species and developmental stages. Rabbits are used for only a single round of infestation for the maintenance of uninfected colonies. In order to maintain genetic diversity and avoid teratogenic effects of inbreeding, CDC colonies are periodically supplemented with ticks acquired from external laboratories or nature. Routine molecular testing of ticks and serological testing of all rabbits used in colony maintenance are performed to confirm the pathogen-free status of the CDC tick colonies (Troughton and Levin 2007). Ticks from the field are only introduced to our colonies after proved uninfected.
The CDC colony of Dermacentor variabilis originated from wild ticks collected in the vicinity of Atlanta, GA in 1999, and the pathogen-free status of this colony has been confirmed in every tick generation. In 2012, we acquired D. variabilis ticks (80 adults and 3 batches of larvae) from an external laboratory for supplementation of the CDC colony. That outside colony was derived from adult ticks collected from vegetation in southeastern Virginia and northeastern North Carolina. The newly received supplemental adult ticks were fed on rabbits, after which the engorged males were tested by PCR for the presence of rickettsial DNA, while females were kept to produce progeny.
Serological testing is performed routinely on serum drawn from rabbits 2 weeks post infestation. Indirect immunofluorescence assay (IFA) is used, as previously described (Lennette et al. 1995), to detect antibodies to multiple agents including Rickettsia, Anaplasma, and Ehrlichia. Fluorescein isothiocyanate–labelled goat α rabbit immunoglobulin G (IgG) (H+L) conjugate (KPL Inc., Gaithersberg, MD) is diluted per manufacturer's instructions. Relevant to this study, whole cell Rickettsia conorii antigen was used to detect antibodies to spotted fever group (SFG) rickettsiae in rabbit sera screened at 1:16 dilution.
Tick DNA was extracted using the DNeasy Blood and Tissue Kit (Qiagen Inc., Valencia, CA) according to the manufacturer's protocols and eluted in 100 μl of buffer (final volume). Real-time PCR was used to detect ompA gene of Rickettsia as described previously (Eremeeva et al. 2003). A plasmid of R. massiliae and distilled water were used as positive and negative controls, respectively, and included in each PCR run. No prior work had been done with either culture isolates or genomic DNA of R. slovaca in either the testing facilities at CDC or in the external laboratory that was the source of supplemental D. variabilis.
The species identity of the infected supplemental ticks was confirmed by sequencing of the 12S mitochondrial ribosomal DNA in five randomly selected specimens (Beati and Keirans 2001). Ten random PCR-positive for Rickettsia samples from supplemental D. variabilis were selected for subsequent sequencing. Multiple locus sequence typing analysis of the Rickettsia gene fragments included partial sequences of the 17 kDa surface antigen (Anderson and Tzianabos 1989), gltA (Labruna et al. 2004), ompB (Roux and Raoult 2000), ompA (Roux et al. 1996), rpoB (Paddock et al. 2010), and sca4 (Sekeyova et al. 2001) genes. An ABI PRISM 3.0 BigDye Terminator Cycle Sequencing Kit (Applied BioSystems, Foster City, CA) was used for performing sequence reactions, as recommended by the manufacturer. The amplicons were purified using the Wizard SV Gel and PCR Clean-Up System (Promega, Madison, WI) and sequenced on an Applied BioSystems 3130xl genetic analyzer. Sequences were assembled using DNASTAR lasergene 9. Homologous sequences were detected using the National Center for Biotechnology Information, by basic local alignment sequence tool application.
Animal model
Guinea pigs (Cavia porcellus) were acquired from Charles River Laboratories and housed in the CDC biosafety level 3 animal facility for the duration of the experiments. Food and water were provided ad libitum. Two sets of three male guinea pigs were used to study clinical, pathological, and serological features of infection caused by the detected Rickettsia sp. The first set of three guinea pigs was needle-inoculated with the rickettsial isolate cultured from infected D. variabilis (Killmaster et al. 2016). Each guinea pig was inoculated intraperitoneally (0.8 mL) and intradermally (0.15 mL)—in the ear— with 1 × 106 rickettsiae suspended in 1 mL of phosphate-buffered saline solution. The needle-inoculated guinea pigs were kept together in the same cage for the duration of the study. The other three guinea pigs were each infested with 46 Rickettsia-infected D. variabilis nymphs (prevalence of infection determined by PCR, 100%) previously fed as larvae on a NZW rabbit. For infestation, guinea pigs were housed individually. Body temperature, clinical signs and behavior were monitored daily; ear skin biopsy, whole blood (50 μL), and serum (200 μL) samples were aseptically collected from each animal twice a week throughout the study. Two guinea pigs from each set were euthanized at day 8 and one animal from each set at day 18 post inoculation/infestation. Samples of liver, spleen, testes, lungs, heart, ear skin, and blood were obtained for PCR and immunohistochemistry (IHC) and serum was collected for serological testing.
Whole blood DNA was extracted using Qiagen FlexiGene DNA extraction kit (Qiagen Inc.) according to the manufacturer's protocol and eluted in 100 μL of elution buffer. Tissue DNA extraction and the PCR procedure for detection of rickettsial DNA were the same as those for testing tick DNA samples. Baseline sera was drawn from each guinea pig pre-infestation or inoculation, and IFA was used to detect development of antibodies to the detected Rickettsia sp. The same IFA protocol was used for guinea pigs as was for rabbits, using FITC-labelled goat α guinea pig IgG (H+L) conjugate (KPL Inc.). Guinea pig sera was initially screened at 1:16 dilution and positive samples were titrated to endpoint in a two-fold dilution series. Serologic data are reported as the reciprocal of the last dilution showing positive fluorescence. IgG titers ≥1:16 were considered positive. IHC testing was performed by the Pathology Branch at CDC using an immunoalkaline phosphatase technique with a polyclonal anti-R. rickettsii antiserum and appropriate positive and negative controls as described earlier (Paddock et al. 1999).
Vector competence study
The supplemental ticks were maintained separately from the CDC colony and fed on naïve NZW rabbits. Representative numbers of ticks from each life stage were tested to verify the presence of rickettsial DNA either individually (adults) or in pools (five nymphs or approximately100 eggs per pool). To assess the ability of D. variabilis for horizontal transmission of the detected Rickettsia sp., nymphal ticks from the infected colony were fed upon six naïve male guinea pigs. Twenty-five donor nymphs from an infected cohort were placed inside the feeding bag attached to the back of each guinea pig. Approximately 300 larvae from the uninfected colony were added 24 hours later into each bag to serve as recipients. Engorged ticks were collected and counted daily for the duration of infestation. After molting into adults, a total of 40 male and 50 female donor ticks from all six guinea pigs were tested individually for the presence of rickettsial DNA by PCR. Also, twenty-five individual engorged recipient larvae and 58 pools of molted nymphs (5 to 12 pools per animal depending on tick availability) were tested. Minimum infection prevalence (MIP) given for pooled nymphs was calculated by assuming one infected tick per pool.
The statistical analysis was conducted by calculating 95% confidence interval.
Results
Detection and molecular characterization of the Rickettsia sp. in colony-originated D. variabilis
All five specimens of adult ticks sequenced for species confirmation were identified as D. variabilis (100% identity to S83088.1 from the National Center for Biotechnology Information).
Feeding of the supplemental adult ticks on naïve NZW rabbits resulted in the development of antibodies reactive to SFG rickettsial antigen. Forty-five out of 46 (97.8%) DNA extracts obtained from D. variabilis males were positive for the presence of SFG rickettsial DNA by PCR. To identify the Rickettsia species, partial sequences of ompA gene of 10 PCR-positive tick samples were obtained and analyzed. All 10 sequences were identical to each other, but their relatedness to rickettsial agents such as R. montanensis or R. rickettsii—both of which could be expected in D. variabilis–was low (Table 1). Unexpectedly, the 608 bp fragment of ompA (GeneBank accession number KR559552) was 100% identical to that of R. slovaca str. D-CWPP (GeneBank accession number CP003375.1) and R. slovaca str. 13-B (CP002428.1). Additional partial sequences of 17 kDa surface antigen (KR559550), gltA (KR559551), rpoB(KR559549), ompB (KR559553), and sca4 (KR559554) genes were obtained and compared with those of R. slovaca using basic local alignment search tool. All sequences were found to have 100% identity to those of R. slovaca str. D-CWPP (GeneBank accession number CP003375.1) and R. slovaca str. 13-B (CP002428.1). Conservative genes, such as 17 kDa antigen and rpoB, were also found to be 100% identical to those of some other SFG rickettsiae (Table 1).
Ricksettia slovaca, b R. rickettsii, c R. africae, d R. parkeri, e R. montanensis, f R. japonica, g R. sibirica sibirica, h R. endosymbiont of A. tuberculatum,
R. philipii str. 364D, j R. conorii, k R. sibirica mongolotimonae.
Sequences of ompA and ompB genes of D. variabilis-derived Rickettsia were only 92% and 93% identical to those of R. montanensis respectively; other gene targets demonstrated 98–99% identity (Table 1). Percent identity among sequences of the detected Rickettsia sp. and R. rickettsii ranged from 97% for ompA gene and 98% for ompB and sca4 genes to 99% for 17 kDa surface antigen and gltA and rpoB genes (Table 1).
Infection of guinea pigs: culture inoculation versus tick bite
Three inoculated guinea pigs did not exhibit any behavioral changes and remained afebrile for the duration of the study. However, all three animals developed scrotal edema and vasculitis in ears and foot-pads, as well as desquamation of skin on the feet (Table 2). At necropsy, localized hepatocellular necrosis or widespread vascular hemorrhages in lungs were observed in all 3 guinea pigs (Fig. 1). Over the duration of the experiment, rickettsial DNA was detected in two out of four blood and three out of four skin samples collected from one of the animals, and five out of seven skin samples from another (Table 2). Rickettsial DNA was not detected by PCR in internal organ samples collected from the needle-inoculated guinea pigs. IHC tests performed on the samples from the same organs indicated presence of rickettsial antigen in testes and skin samples from two out of three animals. All three guinea pigs seroconverted by day 8 post inoculation with IgG titers ranging from 128 to 256. An increase in the IgG titer to 512 was observed in one needle-inoculated guinea pig kept until day 18 post inoculation (Table 2).
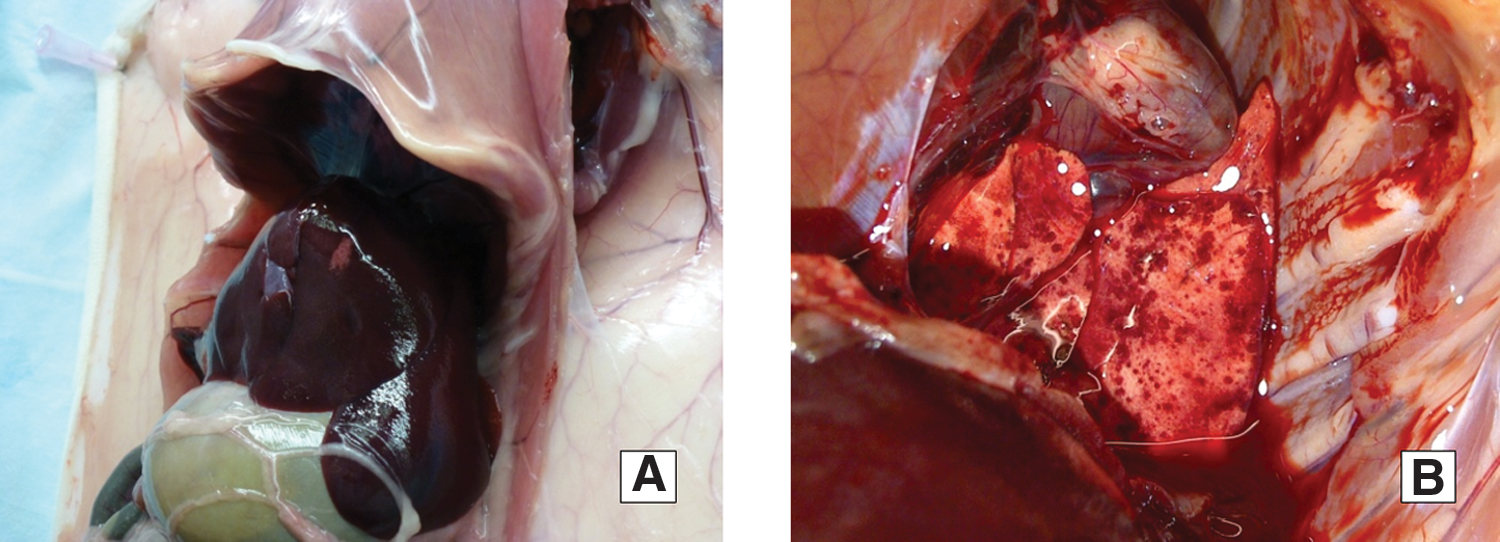
Hepatocellular necrosis
Organs used for PCR testing: liver, spleen testis, lungs, and heart.
GP, guinea pig; IHC, immunohistochemistry; IFA, immunofluorescence assay; Pos, positive; Neg, negative; NI, needle-inoculation; sampl, number of samples (total); TI, tick-infestation.
Less prominent infection was observed in the tick-infested group. Except for dermatitis signs in one of the three guinea pigs, no other clinical and behavioral signs were recorded. Also, no pathological changes or rickettsial antigen were detected at necropsy and IHC. All DNA samples from blood, skin, and organs from all three animals were PCR negative. All three guinea pigs developed antibodies to spotted fever group rickettsiae by day 8 post infestation; however, titers were lower than those in inoculated group, ranging from 16 to 64. No antibodies were detectable in a tick-infected guinea pig at 18 days post infestation (Table 2).
Vertical and horizontal transmission of R. slovaca-like agent by D. variabilis ticks
Rickettsial DNA was detected in all developmental stages of D. variabilis. After oviposition, 20 egg batches were tested by PCR and all of them were positive. Larvae hatched from infected egg batches were fed on a naïve NZW rabbit in the next feeding round. All five pools of molted nymphs were positive. After feeding as nymphs, 10 of 10 males and 10 of 10 females were PCR-positive for the presence of rickettsial DNA (Fig. 2).
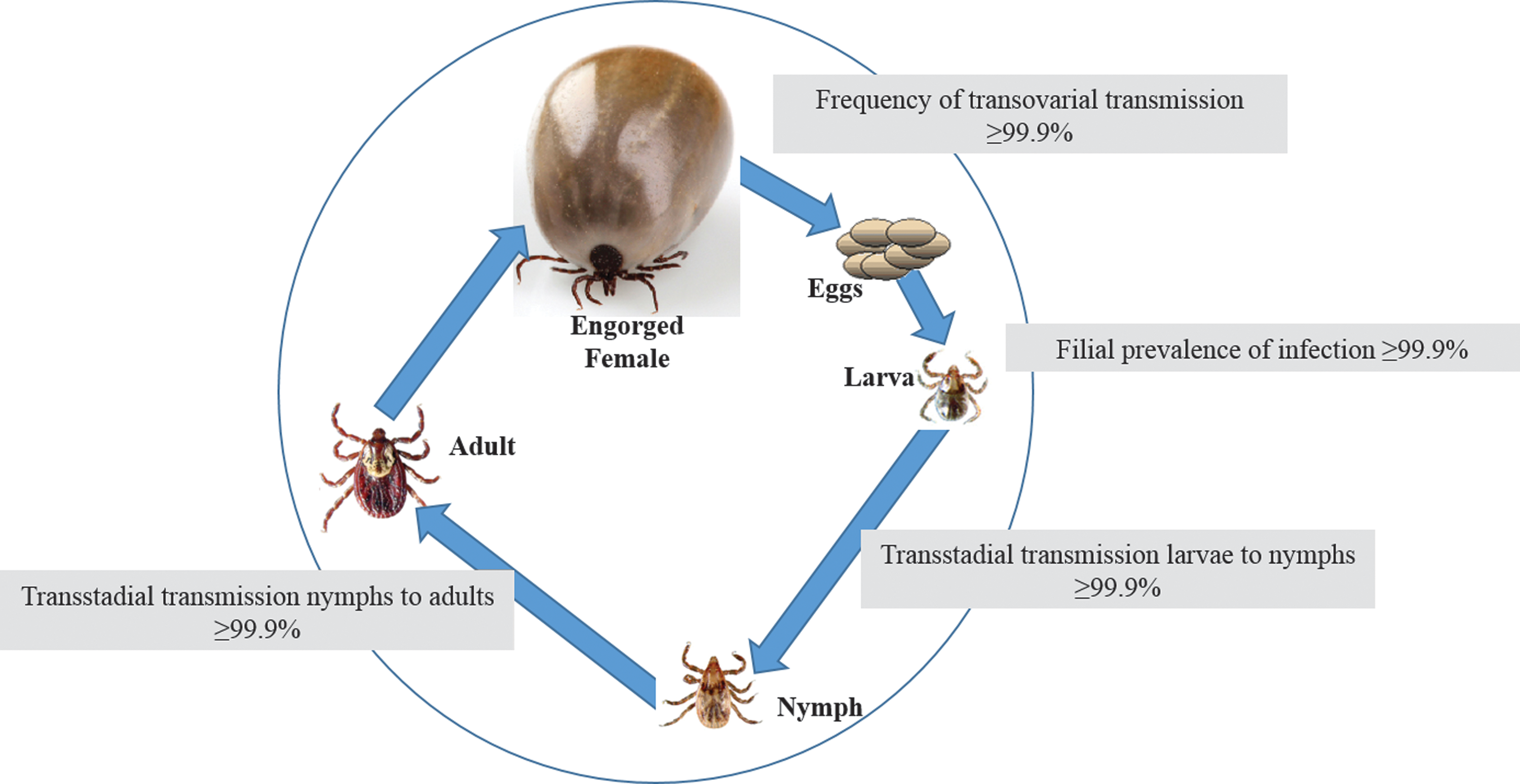
Vertical transmission of R. slovaca-like agent by D. variabilis ticks fed on naïve rabbits.
For assessment of horizontal transmission of R. slovaca-like agent between infected and uninfected ticks, nymphs that were originally received and fed upon rabbits as infected larvae were placed on naïve guinea pigs simultaneously with uninfected D. variabilis larvae (Fig. 3). A majority of the donor D. variabilis nymphs and recipient larvae successfully fed to repletion. Nymphs (n = 101) were allowed to molt into the next developmental stage. Among resulting adults 100% of males (n = 40) and 100% of females (n = 50) contained rickettsial DNA (Fig. 3). Prevalence of infection among engorged recipient larvae was 36 ± 7.7% (54/150) by PCR. From 58 nymphal pools, 34 were PCR-positive, resulting in 11.7 ± 3.7% MIP (Fig. 3).
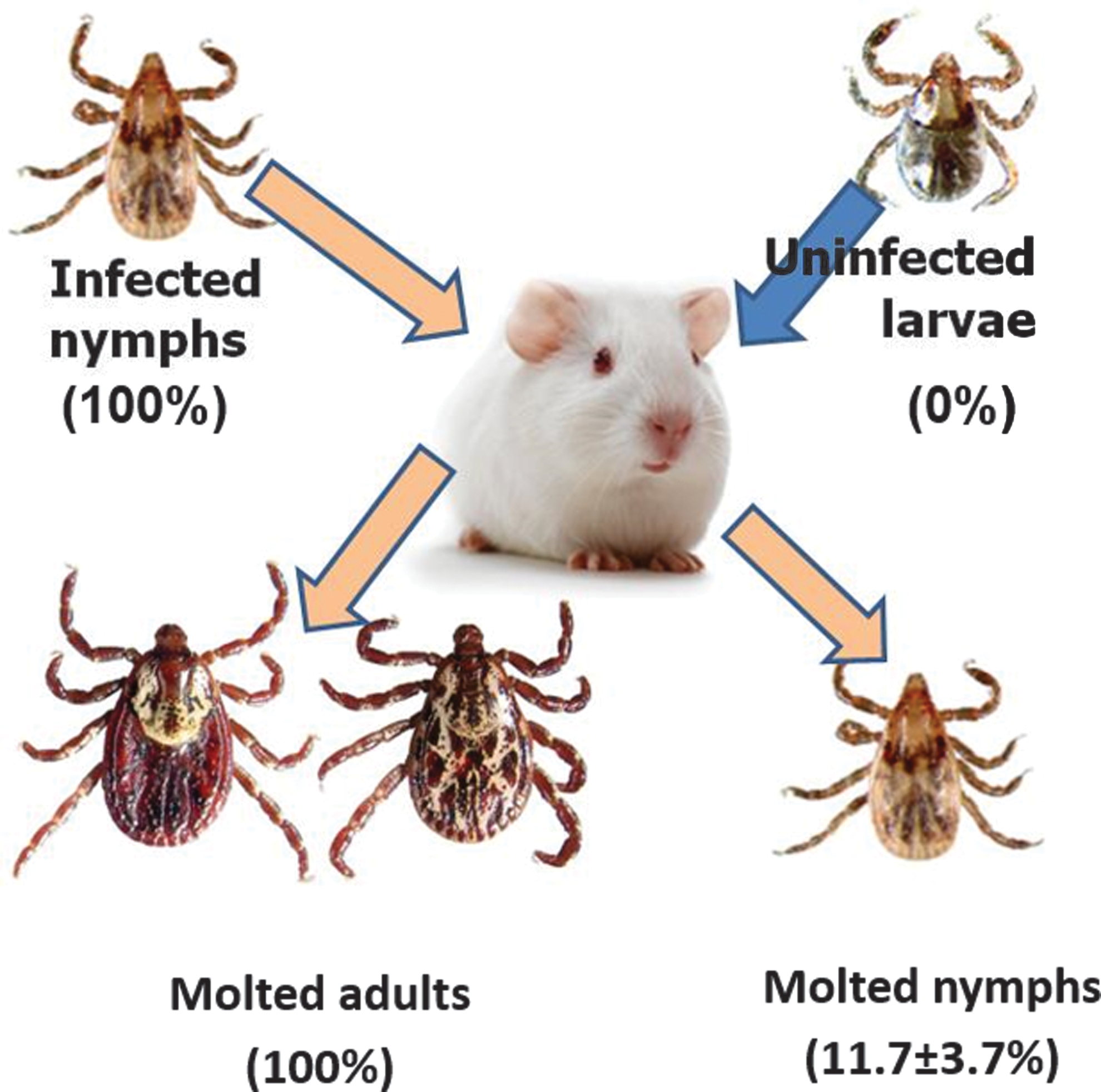
Horizontal transmission of R. slovaca-like agent from guinea pigs infected by D. variabilis nymphs to uninfected D. variabilis larvae.
Discussion
For the first time, we report the detection of a Rickettsia sp. genetically identical to R. slovaca in a colony of Dermacentor variabilis derived from wild ticks collected from the Virginia and North Carolina region. Eight Dermacentor spp. are found in the United States including D. (A) nitens Neumann, D. albipictus (Packard), D. andersoni, D. halli McIntosh, D. hunteri Bishopp, D. occidentalis Marx, D. parumapertus Neumann, and D. variabilis (Yunker et al. 1986), many of which have medical and veterinary importance. SFG Rickettsia spp. known to be associated with Nearctic vectors of the genus Dermacentor so far include R. rickettsii, R. montanensis, and recently identified 364D Rickettsia (Rickettsia phillipi, proposed) (Ammerman et al. 2004, Shapiro et al. 2010). Other known tick-borne rickettsial agents found in the United States include the human pathogen R. parkeri as well as several endosymbiotic bacteria: R. amblyommii, R. andeanae, Rickettsia endosymbiont of A. tuberculatum present in Amblyomma ticks, and rickettsial endosymbiont of Ixodes scapularis (Paddock et al. 2004, Ferrari et al. 2012, Zemtsova et al. 2012). The only species of SFG Rickettsia that have been reported to date from both sides of the Atlantic Ocean are Rhipicephalus-borne R. massiliae (Eremeeva et al. 2006, Oteo et al. 2006) and Amblyomma-borne R. africae (Kelly 2006). The presence of R. slovaca in the Western hemisphere has not been reported to date.
Therefore, when D. variabilis ticks acquired from an external laboratory tested PCR-positive for a SFG Rickettsia, we expected to find either R. montanensis or R. rickettsii. Surprisingly, the detected Rickettsia had only 92–99% of sequence homology with these anticipated agents. On the other hand, multiple locus sequences demonstrated 100% identity of the detected Rickettsia to European strains of R. slovaca. The available data are not sufficient to ascertain whether the rickettsial agent reported here represents one of the known European strains or a unique strain of R. slovaca as only partial sequences of six genes covering a small part of the whole rickettsial genome were characterized. It is noteworthy however, that neither the external laboratory providing supplemental ticks, nor our own laboratory at CDC has ever worked with either R. slovaca or any European ticks of the genus Dermacentor prior to this finding. Therefore, the original source of infection in American dog ticks remains unknown at this point. As a working hypothesis for further research, we theorize that infected ticks, or the pathogen, or both had been imported to the United States from Europe with either domestic dogs or pigs, which are known hosts for D. marginatus. Once imported into the New World, the pathogen had to be picked up and proliferated by the local population(s) of D. variabilis. Any further speculation regarding the introduction in the United States would be unsupported without additional field-collected data.
An animal model was needed to assess infectivity and immunogenicity of the R. slovaca-like agent. We used guinea pigs as model animals because they are known to be susceptible to infection with various rickettsial agents (Walker et al. 1977). We evaluated two different routes to infect laboratory animals. Needle inoculation with the culture is a widely used technique to deliver a controllable amount of live organisms, while infection via a tick bite is a natural route of transmission (Rehacek 1984). In our study both routes provided us valuable information. When guinea pigs received a large dose of the rickettsial agent, they developed clinical signs, had pathological changes observed upon necropsy, and demonstrated relatively strong antibody response. Infestation by infected ticks resulted in a mostly unapparent infection with minor clinical manifestation in only one animal. However, seroconversion in both groups of guinea pigs as well as in NZW rabbits indicates that ticks were capable of transmitting this agent to vertebrate hosts during feeding.
In this study, infected D. variabilis were able to maintain R. slovaca-like agent by both transstadial (larvae to nymphs and nymphs to adults) and transovarial routes while feeding on either naïve rabbits or guinea pigs. Our results also demonstrate successful transmission of this Rickettsia between infected and uninfected D. variabilis feeding simultaneously on guinea pigs. Potential transmission routes include acquisition of rickettsial agent from systemically infected animal and co-feeding. Considering that several skin biopsy, blood, and organ samples from guinea pigs were PCR-positive, uninfected ticks might have acquired this Rickettsia from an infected host. On the other hand, co-feeding is a highly efficient way of transmission of rickettsial agents between uninfected and infected ticks even in the absence of systemic infection (Zemtsova et al. 2010). Regardless of the specific acquisition route, a high proportion of engorged larvae contained detectable level of rickettsial DNA. At least 11.7% of resulting nymphs remained infected after the molt demonstrating that the agent acquired by feeding ticks was also transstadially transmitted. The represented MIP assumes that only one of five pooled ticks was infected; in reality, the infection prevalence among molted nymphs might have been higher.
To our knowledge, this is the first report of identification of a European pathogen R. slovaca or a highly similar agent in the American dog tick, D. variabilis. Whether this finding is relevant to the public health and epidemiology of SFG rickettsioses in the United States will have to be evaluated in the future research regarding the presence of this agent in natural populations of D. variabilis, its pathogenicity to wildlife vertebrate hosts and a potential risk for humans.
Footnotes
Acknowledgments
The authors thank the partners for providing supplemental ticks, and CDC colleague Alyssa Snellgrove for her invaluable help with the maintenance of ticks and testing samples. They gratefully acknowledge Dr. Chris Paddock for performing IHC and Dr. Sandor Karpathy for his contributions to this study.
Author Disclosure Statement
No competing financial interests exist.