
Abstract
Severe fever with thrombocytopenia syndrome (SFTS) is caused by SFTS virus (SFTSV), a novel bunyavirus reported to be endemic to central-northeastern China, southern Japan, and the Republic of Korea (ROK). To investigate SFTSV infections, we collected serum samples and ticks from wild animals. Using serum samples and ticks, SFTSV-specific genes were amplified by one-step RT-PCR and nested PCR and sequenced. Indirect immunofluorescence assay (IFA) was performed to analyze virus-specific antibody levels in wild animals. Serum samples were collected from a total of 91 animals: 21 Korean water deer (KWD), 3 Siberian roe deer, 5 gorals, 7 raccoon dogs, 54 wild boars (WBs), and 1 carrion crow. The SFTSV infection rate in wild animals was 3.30% (3 of 91 animals: 1 KWD and 2 WBs). The seropositive rate was 6.59% (6 of 91 animals: 5 KWD and 1 WB). A total of 891 ticks (3 species) were collected from 65 wild animals (9 species). Of the attached tick species, Haemaphysalis longicornis (74.86%) was the most abundant, followed by Haemaphysalis flava (20.20%) and Ixodes nipponensis (4.94%). The average minimum infection rate (MIR) of SFTSV in ticks was 4.98%. The MIRs of H. longicornis, H. flava, and I. nipponensis were 4.51%, 2.22%, and 22.73%, respectively. The MIRs of larvae, nymphs, and adult ticks were 0.68%, 6.88%, and 5.53%, respectively. In addition, the MIRs of fed and unfed ticks were 4.67% and 4.96%, respectively. We detected a low SFTSV infection rate in wild animals, no differences in SFTSV infection rate with respect to bloodsucking in ticks, and SFTSV infection for all developmental stages of ticks. This is the first report describing the detection of SFTSV in wild animals in the ROK.
Introduction
S
SFTSV was initially detected in Haemaphysalis longicornis ticks in China, which have been implicated as viral vectors (Yu et al. 2011). Moreover, in the ROK, H. longicornis is a dominant tick species and is associated with several tick-borne diseases (Kim et al. 2006, 2011, Yun et al. 2013). The Korea Centers for Disease Control and Prevention reports that the minimum infection rate (MIR) of SFTSV in H. longicornis was 0.46% (Park et al. 2014).
Transmission cycles are responsible for maintaining SFTSV pathogen in tick populations and allowing infection of people and animals. So, it is important to investigate the transmission cycles of SFTSV. To investigate the prevalence of SFTSV in animals, seroconversion and viremia of SFTSV have been demonstrated in domesticated animals such as goats, sheep, cattle, pigs, and dogs in China (Wen et al. 2012, Niu et al. 2013). However, the role of wild animals in the circulation and transmission of SFTSV remains unclear.
In this study, we investigated the infection rates of SFTSV in wild animals and ticks. This is the first study to report SFTSV in wild animals in the ROK.
Materials and Methods
Sample collections
Ticks and sera were collected from wild animals rescued by Wild Animal Rescue Center (WARC) and Wildlife Management Association (WMA) from May to October 2013 regardless of illness. Serum samples from wild animals were collected in Gangwon, Gyeongnam, and Jeonnam Provinces, and in Ulsan Metropolitan Cities. Sera were stored at −70°C until assayed. Ticks from wild animals were collected in Gangwon, Gyeongnam, Jeonnam, Gyeonggi, and Gyeongbuk Provinces, and in Ulsan and Busan Metropolitan Cities. A total of 91 serum and 891 tick samples were collected from wild animals. Based on microscopic examination, ticks were identified to the species level and classified into various developmental stages according to morphology (Yamaguti et al. 1971). Some of the collected ticks were used to detect SFTSV. Ticks (n = 823) were pooled into 433 groups by species, stage of development, and host (i.e., pooled feeding); they were also classified into 24 pools representing various larval stages (1–20 ticks per pool), 67 pools of nymphal stages (1–5 ticks per pool), and 342 pools of adults (1–2 ticks per pool). Pooled ticks were stored at −70°C until assayed.
RNA preparation
Ticks were washed using 70% ethanol once and then washed three times with sterile deionized water to decontaminate the surface. Pooled ticks were homogenized using the sterile Beadbeater TissueLyser II (Qiagen, Hilden, Germany), according to the manufacturer's instructions, with 420 μL of lysis buffer, 0.4 mg of proteinase K, and 5-mm stainless steel beads at an oscillation frequency of 30 Hz/s for 5 min, and then centrifuged at 20,000 × g for 15 min at −4°C. After centrifugation, 400 μL of supernatant was used for RNA extraction using the Gene-spin™ Viral DNA/RNA Extraction Kit (iNtRON Biotechnology, Seongnam, Korea). A total of 200 μL of serum was used for RNA extraction. The final elution volumes of ticks and sera were 60 μL.
One-step RT-nested PCR
PCR assay was performed according to previously described methods with some modifications (Shimojima et al. 2013). To amplify and detect the S segment of the SFTSV genome, One-Step RT-PCR Premix (Solgent, Daejeon, Korea) was used. The primers of the first PCR were NP2-F (5′-CATCATTGTCTTTGCCCTGA-3′) and NP2-R (5′-AGAAGACAGAGTTCACAGCA-3′), which amplify 461-bp products (Shimojima et al. 2013). Reverse transcription (RT) reactions were incubated at 50°C for 30 min, followed by 94°C for 5 min. The amplification was performed according to the following conditions: 20 s at 94°C, 40 s at 52°C, and 30 s at 72°C for 35 cycles, followed by a final extension step at 72°C for 5 min. The nested PCR primers were N2-F (5′-AAYAAGATCGTCAAGGCATCA-3′) and N2-R (5′-TAGTCTTGGTGAAGGCATCTT-3′), which amplify 346-bp products and were designed by authors. The second PCR was carried out with HS Prime Taq Premix (GeNet Bio, Daejeon, Korea). To minimize contamination by virus particles, the positive control was done separately from sample test. Also, filter plug tip was used in all steps. For the nested amplifications, 1 μL of the primary PCR product was used. The cycling conditions for the nested amplification were the same as those already described for the primary amplification, except that a 54°C annealing temperature and 25 cycles were used. PCR products were observed by 1.5% agarose gel electrophoresis.
Sequencing and phylogenetic analysis
SFTSV-nested PCR products were eluted from the agarose gel using the Qiagen Gel Extraction Kit and were analyzed using an ABI PRISM 3700 DNA Analyzer (Applied Biosystems, Foster City, CA). The sequence data were analyzed using Chromas software (Ver 2.33) and were aligned using CLUSTAL X (Ver 2.1). Aligned sequences were examined based on a similarity matrix. Our sequences were compared with published sequences by using the BLAST program from the National Center for Biotechnology Information Website. To assess the relationships between individual pathogens, a phylogenetic tree was constructed using the neighbor-joining method based on the maximum composite likelihood in MEGA6 (1000 bootstrap replicates).
Indirect fluorescence assay
Vero cells inoculated with SFTSV (derived from a human case in Korea, Kim et al. 2013) were treated with 0.25% trypsin–EDTA in phosphate-buffered saline and cultured on 24-well glass slides suspended in minimum essential medium supplemented with 5% fetal bovine serum and 100 units/mL penicillin and 100 μg/mL streptomycin.
After a 16-h incubation period in a CO2 incubator at 37°C, the infected cells were fixed with acetone and subjected to indirect immunofluorescence assay (IFA).
Wild animal serum samples were tested for antibodies against SFTSV by IFA. Fluorescein-labeled rabbit antideer IgG for Korean water deer (KWD) and Siberian roe deer, fluorescein-labeled rabbit antigoat IgG for goral, fluorescein-labeled goat antidog IgG for raccoon dog, fluorescein-labeled goat antiswine IgG for wild boar (WB), and fluorescein-labeled goat antichicken IgG for carrion crow were used for secondary antibodies. Sera that were reactive at a dilution of ≥1:64 were considered positive. In addition, we checked that all serum samples with SFTSV IgG were negative against uninfected cells.
Results
A total of 91 serum samples were collected from wild animals, including 21 samples from KWD (Hydropotes inermis), 3 Siberian roe deer (Capreolus pygargus), 5 gorals (Nemorhaedus caudatus), 7 raccoon dogs (Nyctereutes procyonoides), 54 WBs (Sus scrofa), and 1 carrion crow (Corvus corone). Of the 91 serum samples from wild animals, SFTSV was detected in 1 (4.76%) KWD and 2 (3.70%) WBs by one-step RT-PCR and nested PCR. In addition, 5 (23.81%) KWD and 1 (1.85%) WB samples were seropositive for SFTSV based on IFA (Table 1).
IFA-positive titers: three KWD were 1:256, two KWD were 1:128, and one WB was 1:512.
IFA, indirect immunofluorescence assay; KWD, Korean water deer;
A total of 891 ticks (587 fed and 304 unfed ticks) were collected from 65 wild animals in seven provinces in the ROK. Of the attached tick species, H. longicornis (74.86%) was the most abundant, followed by Haemaphysalis flava (20.20%) and Ixodes nipponensis (4.94%). The mean number of attached ticks per animal was 18.2 (Table 2). The MIR of SFTSV for tick species was 4.98% (41 positive pools, 823 ticks). The MIRs of H. longicornis, H. flava, and I. nipponensis were 4.51% (27 positive pools, 599 ticks), 2.22% (4 positive pools, 180 ticks), and 22.73% (10 positive pools, 44 ticks), respectively (Table 3). The MIRs of larvae, nymphs, adult males, and adult female ticks were 0.68%, 6.88%, 8.70%, and 4.29%, respectively (Table 3). Of 424 pools (796 ticks) containing exclusively fed (298 pools, 514 ticks) or unfed (126 pools, 282 ticks) ticks, SFTSV was detected in 24 pools (MIR; 4.67%) of fed ticks and 14 pools (MIR; 4.96%) of unfed ticks (Table 4).
These data were minimum values because partial ticks (68) were not analyzed for SFTSV.
This pool was fed larvae.
MIR, minimum infection rate.
A total of 44 samples that were positive based on PCR analyses were confirmed by sequencing analyses. The sequencing results confirmed that the amplified fragments were identical to the nucleocapsid protein (NP) gene sequences of SFTSV. Excluding identical sequences, a sample of 21 representative nucleotide sequences determined in this study was deposited in the GenBank database. All SFTS sequences used in this study and the corresponding accession numbers are listed in Table 5. Virus sequences of phlebovirus species as outgroup of phylogenetic analysis are shown in Figure 1. A phylogenetic tree revealed that isolates identified in most ticks and one WB isolate (KP967531) represent a single clade (Fig. 1), and this clade was distinct from two isolates from wild animals and two isolates from H. longicornis. Furthermore, two isolates (KP967534 and KP967535) from H. longicornis were closely related to a human isolate identified in the ROK. Our isolates were clearly distinct from Japan and China isolates.
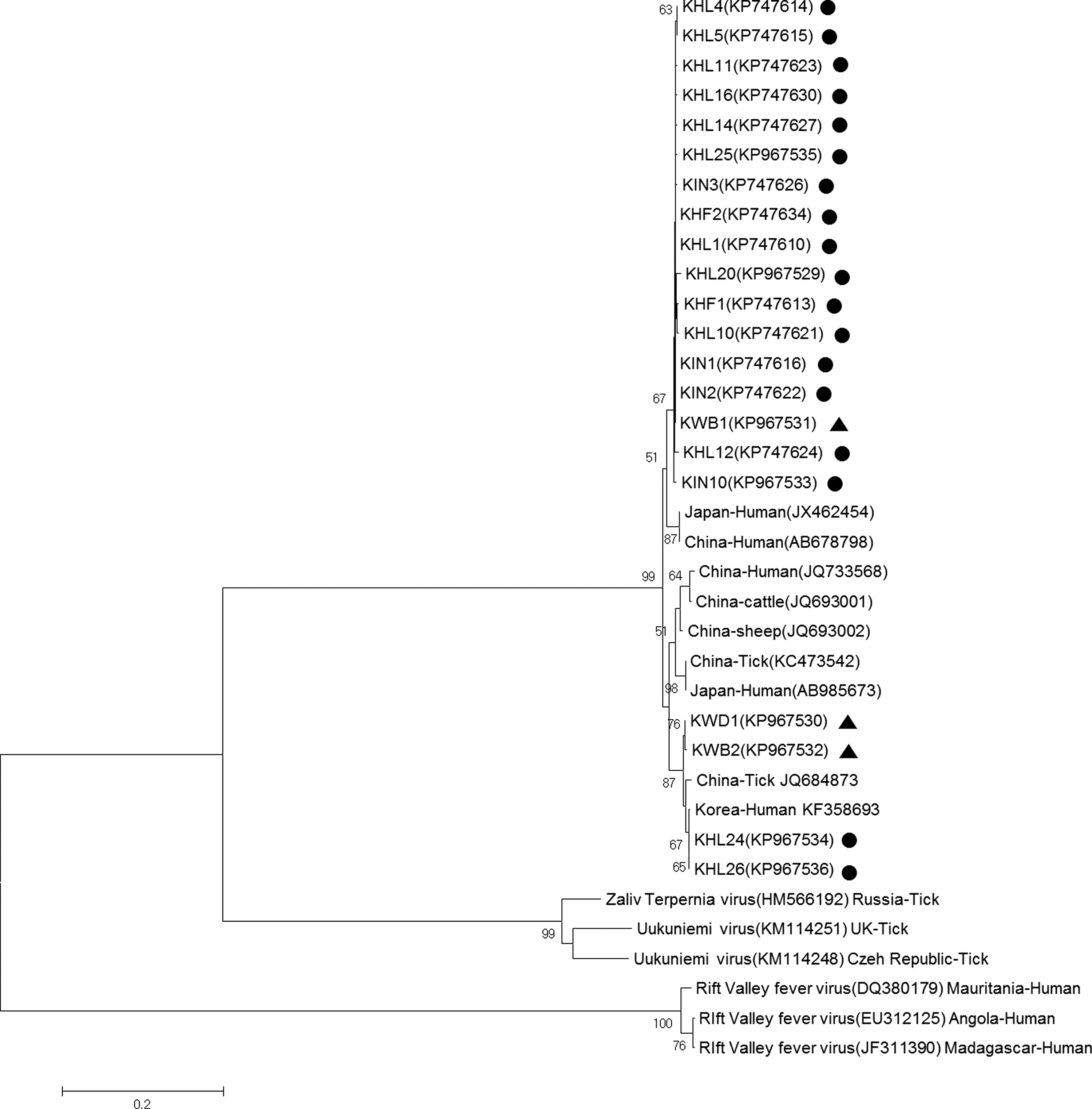
Phylogenetic analysis of severe fever with thrombocytopenia syndrome and other phlebovirus spp. viruses based on partial S segment sequences (346 bp). The tree was constructed using the neighbor-joining method based on the p-distance model in MEGA6 (1000 bootstrap replicates). The scale bar indicates nucleotide substitutions per position. Black dots (•) indicate the original sequences of the SFTSV strains obtained from wild animals in this study; black triangles (▴) indicate the original sequences of the SFTSV strains obtained from ticks in this study. Among the 21 representative strains identified in this study, the Korean strains were detected from Haemaphysalis longicornis (KHL), Haemaphysalis flava (KHF), Ixodes nipponensis (KIN), Korean water deer (KWD), and Korean wild boar (KWB). Numbers at nodes indicate bootstrap values. SFTSV, severe fever with thrombocytopenia syndrome virus.
This study.
Discussion
In this study, of a total 91 serum samples collected from wild animals in four provinces in the ROK, we detected SFTSV RNA in KWD (4.76%) and WBs (3.70%) and seropositivity of SFTSV-specific antibodies in KWD (23.8%) and WBs (1.85%). Previous studies have detected seropositive goats, sheep, pigs, cattle, dogs, and chickens for SFTSV, and SFTSV RNA was detected from these animals in China (Wen et al. 2012, Niu et al. 2013, Ding et al. 2014). Domestic animals used in these studies in China grazed on pastures or hills and roamed freely in fields. Thus, they had opportunities for exposure to SFTSV-infected ticks. However, most domestic animals raised in the ROK are housed indoors. As tick hosts, wild animals may be more important than domestic animals in the ROK.
Recent studies of domestic animals in China have shown that the seroprevalence of SFTSV, based on enzyme-linked immunosorbent assays (ELISA), is relatively high in goats (74.8–83%), sheep (69.5%), cattle (57.1–60.5%), dogs (37.9–52.1%), chickens (35.9–47.4%), and pigs (3.1%) (Wen et al. 2012, Niu et al. 2013, Ding et al. 2014), whereas the prevalence of SFTSV RNA is low in sheep (3.8%), cattle (4.2%), dogs (5.3%), pigs (2.6%), and chickens (1.7%) (Niu et al. 2013). We confirmed the SFTSV seropositivity in KWD (23.8%) and WBs (1.85%) by IFA. These estimates were lower than previous estimates for domestic animals in China. This difference might be caused by host specificity of SFTSV, or methodological differences, among other explanations. According to a recent study, NP in SFTSV that were used for ELISA had potential serological cross-reactivity with the other phleboviruses NP (Wu et al. 2014). Also, our study has some limitations. It was possible that IgG signal was false positive in IFA because of cross-reaction with other pathogens.
In this study, the prevalence of SFTSV in wild animals in the ROK was low. Although we confirmed SFTSV infection in wild animals, three SFTSV-positive samples were verified in the nested step of PCR, but not in the first PCR (data not shown). These results indicated that wild animals may have a low level of viremia.
We investigated SFTSV infections from attached ticks in wild animals. Wild animals were infested with an average of 18.2 ticks, and 32.3% (21/65) of wild animals were infested with SFTSV-positive ticks. According to a recent study, 18 people were bitten by ticks infected with SFTSV and none had any clinical signs or symptoms of SFTS (Yun et al. 2013). Our results did not indicate that 21 wild animals were potential SFTSV positive. This may reflect the frequency of exposure to SFTSV in wild animals.
A total of 891 ticks were collected from 65 wild animals (9 species). H. longicornis (74.86%) was the most abundant species, followed by H. flava (20.20%) and I. nipponensis (4.94%). A previous study revealed the following distribution of questing tick species in the ROK: H. longicornis (90.8%), H. flava (8.8%), I. nipponensis (0.3%), and Ixodes persulcatus (0.05%) (Park et al. 2014). H. longicornis was undoubtedly the most dominant species of questing tick and attached tick in wild animals and humans (Kim et al. 2006, 2011, Yun et al. 2013). In this study, the MIR of SFTSV in attached ticks in wild animals was 4.98%. The MIRs of H. longicornis, H. flava, and I. nipponensis were 4.51%, 2.22%, and 22.73%, respectively. In the ROK, the minimum field infection rate of H. longicornis collected by dragging or sweeping in the environment is 0.46% (912 pools, 11,856 ticks) (Park et al. 2014), the MIR of ticks collected from humans is 6.9% (189 pools, 261 ticks) (Yun et al. 2013), and the MIR of ticks collected from wild animals in this study was 4.98% (447 pools, 823 ticks). In these studies, the mean numbers of tested ticks per pool were 13, 1.38, and 1.84, respectively. The relatively low MIR of questing ticks (Park et al. 2014) may be due to the large number of tested ticks per pool. The MIRs of unfed and fed ticks collected from wild animals in this study were 4.96% and 4.67%, respectively. Based on this result, we speculated that SFTSV infections in wild animals are not acquired during feeding by ticks. Therefore, our results indicate that wild animals may not be essential host reservoirs, but feeding sources for SFTSV infected ticks in SFTSV circulation.
In the case of Anaplasma phagocytophilum, which is a tick-borne pathogen like SFTSV, a variety of mammals are susceptible to infection and can act as host reservoirs to maintain a source of infected ticks (Nicholson et al. 2010). In previous studies, A. phagocytophilum is detected in 63.6% of KWD (Kang et al. 2011), 26% of small mammals (Kim et al. 2006), and 1.94% (506 pools, 1395 ticks) questing ticks (Oh et al. 2009). Unlike A. phagocytophilum, the infection rate of SFTSV in wild animals was not high. Nevertheless, questing ticks in grass were detected at similar rates for A. phagocytophilum and SFTSV. We surveyed larvae that were collected within 0.5 km of the residence of an SFTS patient (2013) in Seok-Mo Island located in Incheon by the flagging method. Samples were collected during October, when larvae increase explosively in the ROK. Using one-step RT-PCR and nested PCR, we confirmed that 0.32% of larvae (MIR, 30 larvae/pool, 1560 total larvae) were infected with SFTSV (unpublished data). In addition, SFTSV has been detected in H. longicornis at all developmental stages in China (Wang et al. 2015).
In this study, we reported the SFTSV infection rate from wild animals and ticks attached to wild animals. This is the first report of SFTSV in wild animals in the ROK. Our results provide information that may be useful to understand the transmission cycles responsible for the maintenance of SFTSV in tick populations and the infection of wild animals.
Conclusions
We reported the SFTSV infection rate from wild animals and ticks attached to wild animals. The SFTSV infection rate in wild animals was 3.30% and seropositive rate was 5.5%. The MIRs of fed and unfed ticks were 4.67% and 4.96%, respectively. Our results provide information that may be useful to understand the transmission cycles responsible for the maintenance of SFTSV in tick populations and the infection of wild animals.
Footnotes
Acknowledgments
This study was supported by the National Institute of Environmental Research (2013) and was partially funded by the BK21 PLUS Program for Creative Veterinary Science Research, Research Institute for Veterinary Science, and College of Veterinary Medicine, Seoul National University. The authors sincerely appreciate the members of the Wild Animal Rescue Center and Wildlife Management Association for providing serum samples and ticks collected from wild animals.
Author Disclosure Statement
No competing financial interests exist.