
Abstract
Members of the order Rickettsiales are small, obligate intracellular bacteria that are vector-borne and can cause mild to fatal diseases in humans worldwide. There is little information on the zoonotic rickettsial pathogens that may be harbored by dogs from rural localities in South Africa. To characterize rickettsial pathogens infecting dogs, we screened 141 blood samples, 103 ticks, and 43 fleas collected from domestic dogs in Bushbuckridge Municipality, Mpumalanga Province of South Africa, between October 2011 and May 2012 using the reverse line blot (RLB) and Rickettsia genus and species-specific quantitative real-time PCR (qPCR) assays. Results from RLB showed that 49% of blood samples and 30% of tick pools were positive for the genus-specific probes for Ehrlichia/Anaplasma; 16% of the blood samples were positive for Ehrlichia canis. Hemoparasite DNA could not be detected in 36% of blood samples and 30% of tick pools screened. Seven (70%) tick pools and both flea pools were positive for Rickettsia spp; three (30%) tick pools were positive for Rickettsia africae; and both flea pools (100%) were positive for Rickettsia felis. Sequencing confirmed infection with R. africae and Candidatus Rickettsia asemboensis; an R. felis-like organism from one of the R. felis-positive flea pools. Anaplasma sp. South Africa dog strain (closely related to Anaplasma phagocytophilum), A. phagocytophilum, and an Orientia tsutsugamushi-like sequence were identified from blood samples. The detection of emerging zoonotic agents from domestic dogs and their ectoparasites in a rural community in South Africa highlights the potential risk of human infection that may occur with these pathogens.
Introduction
R
Spotted fever rickettsiosis has been recognized in South Africa since the beginning of the 20th century (McNaught 1911) with Rickettsia conorii, the agent of Mediterranean spotted fever, having long been associated with human disease in the country (Pretorius and Birtles 2002). Rickettsia aeschlimannii, which causes a Mediterranean spotted fever-like illness, has also been identified in South Africa (Beati et al. 1997). African tick bite fever (ATBF) caused by Rickettsia africae is prevalent in South Africa and is the leading cause of fever among travelers to South Africa (Jensenius et al. 2003). Game hunting and traveling to Southern Africa from November through April increase the risk for ATBF among travelers, and contact with tick-infested cattle and game in areas endemic for certain spotted fever group rickettsiae may also increase the risk of disease (Jensenius et al. 2004).
Rickettsia mongolotimonae, which causes a lymphangitis-associated rickettsiosis, has also been reported in South Africa (Pretorius and Birtles 2004). Rickettsia felis, the cause of flea-borne spotted fever (FBSF), has not yet been reported in South Africa, but has been recognized as an emerging pathogen especially in sub-Saharan Africa (Parola 2011).
Other rickettsial agents include Anaplasma phagocytophilum from the family Anaplasmataceae, which causes canine and human anaplasmosis (Parola and Raoult 2006), and Ehrlichia canis, which causes canine monocytic ehrlichiosis and is also responsible for some cases of human ehrlichiosis (Nicholson et al. 2010). A new strain of Anaplasma sp. closely related to A. phagocytophilum has also been detected in canine blood samples in South Africa (Inokuma et al. 2005). To identify and characterize rickettsial pathogens infecting dogs, we used molecular techniques to screen blood samples and ectoparasites collected from domestic dogs in the Mnisi community area in Mpumalanga Province, South Africa.
Materials and Methods
Study site
The Mnisi community is situated in the northeastern corner of the Bushbuckridge Municipal Area, Mpumalanga Province, South Africa, and located at the livestock/wildlife/human interface of the western boundary of the Kruger National Park. The study area falls within the Savannah region and is adjacent to the Andover and Manyeleti game reserves (Fig. 1). The geographic coordinates are S − 24° 39′, E 31° 20′. A Health and Demographic Surveillance System in Dogs (HDSS-Dogs) was established in the community in 2011. This study sampled dogs from 400 dog-owning compounds that are enrolled in the HDSS project. Further description of the dog population and the HDSS-Dogs can be found in Conan et al. (2015).
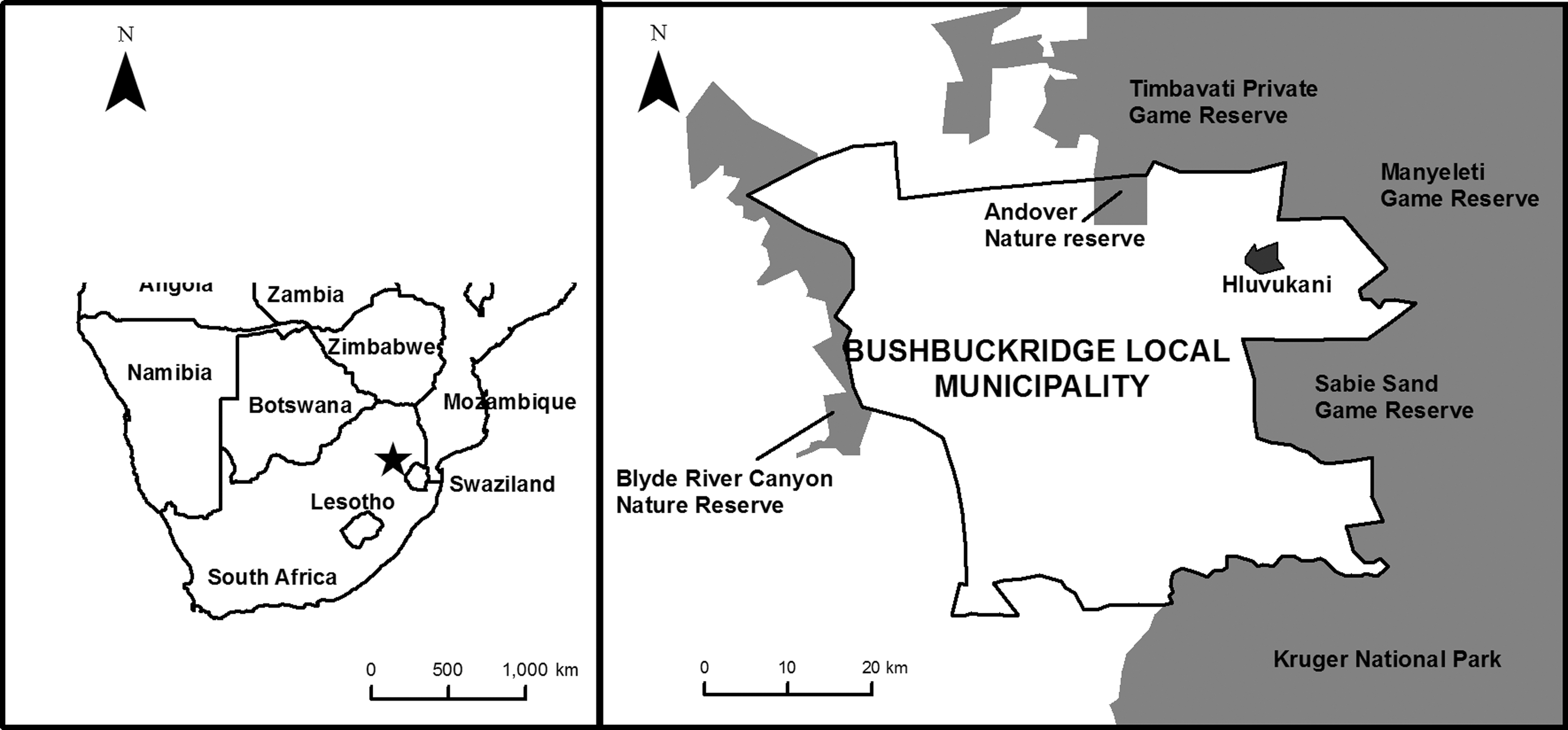
Map of study site showing area where dog-owning households are located.
Collection of blood samples
Blood samples were collected from owned, free roaming, apparently healthy dogs present at households visited by the HDSS-Dogs field team during routine quarterly visits. Convenience sampling of dog-owning households was done from October 2011 to May 2012. During the first two sample collection periods (October and December, 2011), blood was collected directly in capillary tubes (n = 85) and stored on FTA® filter paper (Whatman®). During the third period (April/May 2012), blood was collected in EDTA vacutainer tubes (Lasec) and thereafter transferred to FTA cards (n = 56). Blood samples stored on FTA cards were then sent to the Department of Veterinary Tropical Diseases, University of Pretoria, for analysis.
Collection of ectoparasites
Dogs were inspected for the presence of ectoparasites by brushing the hair with a plastic comb or brush, and a white paper was used for collection of fleas. Live adult ticks were removed from the animals manually using forceps. A total of 103 ticks and 43 fleas were collected. All ectoparasites were preserved in 70% ethanol and identified to species level under a stereomicroscope, according to standard morphological identification guides (Segerman 1995, Walker et al. 2003). Ticks and fleas were pooled into 10 pools of ticks and 2 pools of fleas according to their species. Large numbers of ticks from the same species were placed in separate pools to reduce possible dilution if only a small number of ticks in a pool are positive. The maximum number of ticks in a pool was 14. The tick pools were collected from 64 dogs. Dogs that had more than 1 tick collected from them were 16 in number.
DNA extraction and reverse line blot hybridization assay
DNA was extracted from 141 blood samples spotted on filter cards using the QIAamp DNA mini kit® (Qiagen) according to the manufacturer's instructions to a final elution volume of 100 μL. The tick and flea samples were homogenized using a Tissue Lyser® (Qiagen) and DNA extraction was performed using the QIAamp DNA mini kit (Qiagen) according to the manufacturer's instructions. PCR was conducted with a set of primers (Ehr-F and Ehr-R) that amplify the V1 hypervariable region of the 16S rRNA gene of Ehrlichia and Anaplasma species (Bekker et al. 2002). The PCR was performed as previously described (Gubbels et al. 1999) on the Gene Amp® PCR system 9700 (Applied Biosystems). PCR amplicons were then screened using the reverse line blot (RLB) hybridization assay as described by Gubbels et al. (1999). The Ehrlichia and Anaplasma genus-specific and species-specific oligonucleotide probes used for the assay are shown in Table 1.
Detection of rickettsiae
A Rickettsia genus-specific assay amplifying a 115-bp fragment of the 17 kDa surface protein gene of Rickettsia was used to screen blood and ectoparasites pools. Two species-specific qPCR assays targeting the ompB gene of R. africae and R. felis were used to screen ectoparasite DNA samples as previously described (Henry et al. 2007, Jiang et al. 2012, Maina et al. 2014). Species-specific assays were not performed on blood samples because they were all negative on the Rickettsia genus assay. Specific plasmid DNA was used as positive control for each assay, and PCR-grade water replaced the DNA template in negative control reactions.
Sequencing and phylogenetic analysis
Five randomly selected samples (97, 98, 107, 115, and T3) that tested positive with genus-specific probes for Anaplasma, Ehrlichia, and Rickettsia spp. by RLB or qPCR assays were further analyzed by DNA sequencing (Inqaba Biotechnologies) to determine specific spp. For sequencing reaction, the same primers used for PCR amplification were used, except that they had no biotin incorporated. Furthermore, three other samples (106, 116, and 125) were randomly selected to identify specific Anaplasma and Ehrlichia infections. These samples, which had strong signals on the RLB assay for Anaplasma and Ehrlichia, were selected for next-generation sequencing (NGS) using DNA barcoding.
The tick pool T8, which was positive for Rickettsia spp., but negative on the species assay, was also selected as well as F1, which was selected as a representative pool from the flea samples, was analyzed using the similar approach. For NGS using DNA barcoding, the 16S rRNA gene was amplified from genomic DNA of selected samples using universal primers, 341F and 785R, modified with Illumina-specific adapters (Klindworth et al. 2013). Sequencing was performed on the Illumina's MiSeq platform using an MiSeq v3 kit.
The 16S rRNA sequences obtained from PCR amplicons were prepared for assembly using the PreGap4 program of the Staden package (version 2.0 for Windows), and Basic Local Alignment Search Tool (BLAST) searches were performed using MegaBlast from the BLAST (
GenBank accession numbers
The sequences of the molecular isolates (97, 98, 107, 115, T3, 125, F1, 106,116, and T8) obtained from blood samples, Rhipicephalus sanguineus, Ctenocephalides felis strongylus, and Haemaphysalis elliptica pools have been submitted to GenBank with accession numbers KP823591-KP823600 for the 16S rRNA gene.
Results
Detection of hemoparasites in blood
The results of the RLB hybridization analysis for DNA prepared from 141 blood samples spotted on FTA filter cards showed the presence of Ehrlichia and Anaplasma, species detected either as single or mixed infections. Ehrlichia/Anaplasma species was detected in 70 (50%) samples, E. canis was detected in 23 (16%) samples, and 51 (36%) samples were negative.
Detection of hemoparasites from ticks and fleas
The tick species collected included Haemaphysalis elliptica (n = 30, pools T1–T3), Amblyomma hebraeum (n = 27, pools T4 and T5), R. sanguineus (n = 27, pools T7 and T8), Rhipicephalus simus (n = 18, pools T9 and T10), and one unspeciated Ixodes (T6). The fleas included Ctenocephalides felis strongylus (n = 23, pool F1) and Echidnophaga gallinacea (n = 20, pool F2). The results of the RLB hybridization analysis of DNA from pooled tick and flea samples also showed the presence of Ehrlichia and Anaplasma species. Table 2 shows results of RLB analysis of ectoparasites.
RLB, reverse line blot.
Detection of Rickettsia by qPCR
DNA samples from blood and ectoparasites were subjected to a Rickettsia genus-specific qPCR assay for detection of rickettsial infections before testing with species-specific qPCR assays, which identify R. africae and R. felis DNA. Rickettsia DNA was not detected from any of the DNA samples prepared from blood when using the Rickettsia genus-specific assay. By contrast, 7/10 (70%) tick pools and 2/2 (100%) flea pools tested positive for Rickettsia DNA using the same assay (Table 3). Analysis with species-specific qPCR assays revealed that 3/10 (30%) tick pools were positive for R. africae and 2/2 (100%) flea pools were positive for R. felis. Rickettsial DNA was not detected in three tick pools (T1, T2, and T7). The minimum infection rate (MIR), which assumes that a positive pool contains a single infected pool (Cowling et al. 1999), was calculated for the positive H. elliptica pool as 3.3%; MIR for T4, an A. hebraeum pool, was 7.7%; MIR for T5, the second A. hebraeum pool, was 7.1%; MIR for R. sanguineus pool T8 was 3.6%; and MIR for R. simus pools (T9 and T10) was 11.1%.
+, qPCR positive; −, qPCR negative.
qPCR, quantitative real-time PCR.
Sequence and phylogenetic analysis
Analysis of the 16S rRNA gene revealed that a sequence obtained from DNA of H. elliptica pool (T3) was 99% (461/466) similar to R. africae and was placed in the same clade as R. africae 95.2 (acc. no. JF949783) (Fig. 2). The sequence from R. sanguineus pool (T8) was found to have 96% (291/303) sequence identity to Rickettsia peacockii (NR118837) and was 95.7% (290/303) similar to other rickettsiae. This sequence (T8) was placed between the TGR and the STGO. Analysis showed that the sequence of one of the positive flea pools (F1) (264 bp) and blood sample 106 (265 bp) were identical and portrayed 100% sequence identity with Ca. R. asemboensis (JN315973 and JN315967). The sequence of blood sample 116 identified O. tsutsugamushi with 96.1% (247/257) homology and was 95.7% (246/257) similar to the closest/other rickettsiae. This sequence was placed in the same clade as O. tsutsugamushi Kawasaki strain (acc. no. D38625) (Fig. 2).

Neighbor-joining phylogenetic tree of the 16S rRNA gene sequences of Rickettsia species generated from this study together with homologous sequences from GenBank. Bootstrap analyses were performed with 1000 replications (MEGA software version 6). Gray square indicates sequences obtained from the study.
Phylogenetic analyses of the Anaplasma species 16S rRNA sequences from blood samples 97, 98, 107, and 117 showed that these sequences share 99% similarity with Anaplasma sp. South Africa dog 1076, 1108, and 1245 (acc. nos. AY570539, AY570538, and AY570540) and clustered together with the latter (Fig. 3). The sequence of blood sample 125 had a 99% identity to sequences of A. phagocytophilum strains, ApGDr1, ApGDr2, and GDR4 (acc. nos. KC800963, KC800964, and KC455366), and was placed in the same clade with A. phagocytophilum species (Fig. 3).
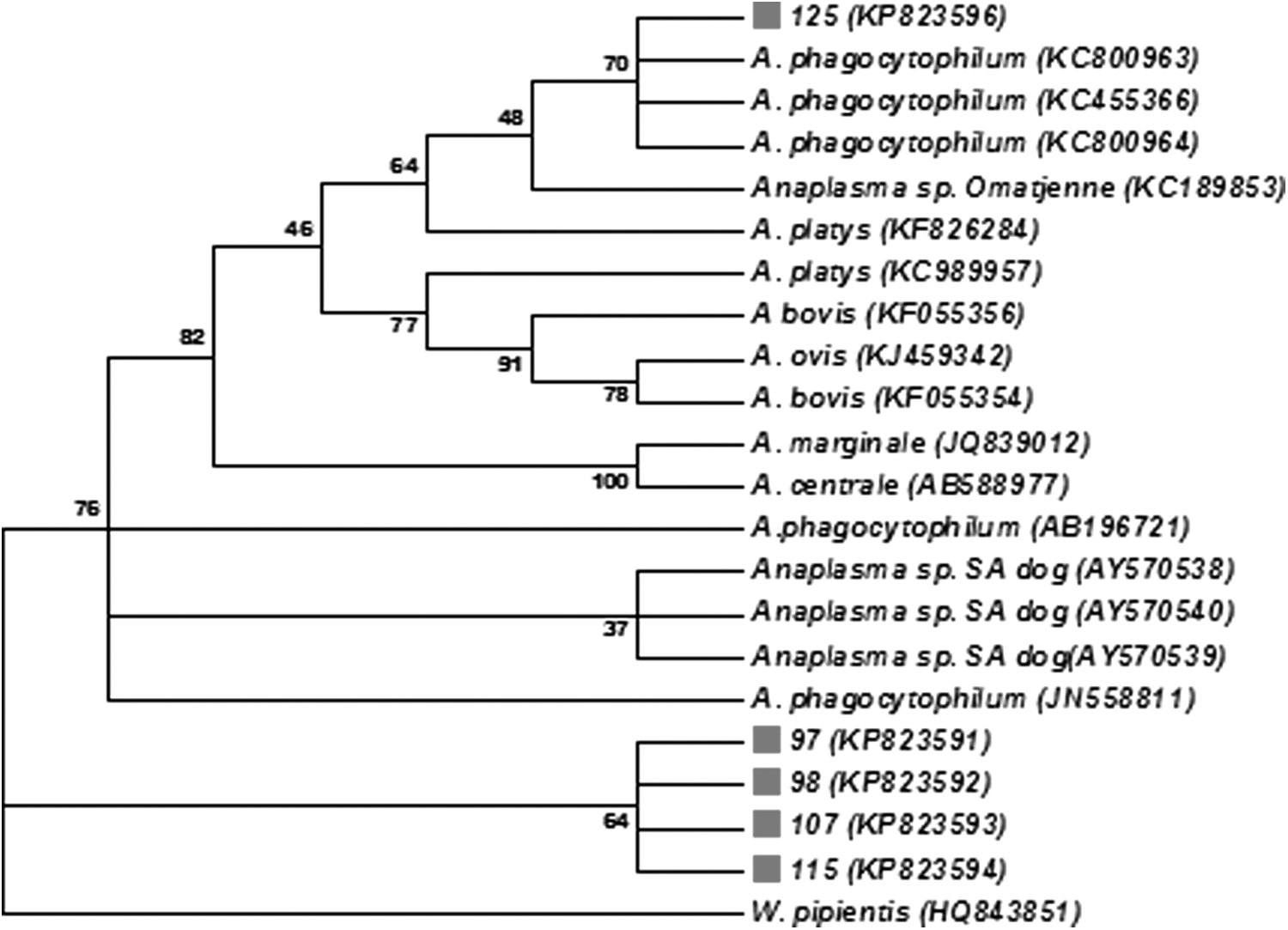
Neighbor-joining phylogenetic tree of the 16S rRNA gene sequences of Anaplasma species generated from this study together with homologous sequences from GenBank. Bootstrap analyses were performed with 1000 replications (MEGA software version 6). Gray square indicates sequences obtained in the study.
Discussion
We detected R. africae in A. hebraeum and H. elliptica ticks tested with the R. africae-specific qPCR assay; this is consistent with previous molecular studies in Africa where the organism was detected in Chad, Burundi, Ethiopia, Senegal, and in southern Africa (Jensenius et al. 2003). R. africae is the cause of ATBF, an emerging infectious disease transmitted by ticks (Jensenius et al. 2003). Many cases of human infections with R. africae have been acquired from South Africa, which has many wildlife tourist centers usually situated in areas in which tick vectors are endemic (Chmielewski et al. 2013). In South Africa, R. africae is transmitted mainly by A. hebraeum (Jensenius et al. 2003). Other tick species have been found to harbor R. africae (Parola et al. 2001, Mediannikov et al. 2012); however, this is the first time that the organism has been detected in H. elliptica, thereby extending the known host range of the organism. The detection of R. africae from A. hebraeum, a tick known for readily biting humans, and H. elliptica, one of the most common ticks infesting domestic dogs in South Africa, underpins the potential risk of human infection with this pathogen in the study area.
This is the first report of members of the R. felis clade being detected in South Africa. R. felis is an emerging zoonotic agent that has been found in several countries globally, causing FBSF in humans (Parola 2011). The cat flea, C. felis, is the only biological vector and reservoir of R. felis so far identified (Reif and Macaluso 2009). In Africa, R. felis has been detected in humans and arthropods in Gabon, Tunisia, Egypt, Congo, Algeria (Parola 2011), and Kenya where substantial work on the organism has been carried out (Jiang et al. 2013). Sequence analysis of one of the flea pools (F1) positive for R. felis on the species-specific qPCR assay, however, revealed a 100% homology to Ca. R. asemboensis, a newly described rickettsia and an R. felis-like organism (RFLO) previously identified from fleas in Kenya (Jiang et al. 2013). In that study, Ca. R. asemboensis was detected in 60% of flea pools tested.
Detection of Ca. R. asemboensis in pool F1 in our study shows that the R. felis species-specific qPCR assay (Henry et al. 2007) is not specific to R. felis, but is able to detect other RFLOs. We found the same organism in a dog blood sample (sample 106)—the first time that this putative new species has been detected in a canine host. Recently, this agent has been detected in the blood of cynomolgus monkeys without signs of infection (Tay et al. 2015). Sample T6 (Ixodes sp.) and tick pools, T9 and T10 (R. simus), were positive with the Rickettsia genus-specific assay, but negative on the two species-specific assays used, implying that they could be positive for species of Rickettsia other than R. africae or R. felis. Sequencing would have to be performed in follow-up studies to determine the specific species. Ixodes ricinus is a known vector and reservoir host of Rickettsia helvetica (Parola et al. 2005), while R. simus is more known as a vector of Anaplasma marginale and Anaplasma centrale (Potgieter et al. 1983).
Sequence analysis of the 16S ribosomal RNA gene using high-throughput sequencing technology detected rickettsiae sequences in two dog blood samples. The samples were positive for the Ehrlichia/Anaplasma genus-specific probe only on the RLB hybridization assay, but were negative on the Rickettsia genus-specific qPCR assay based on the 17-kDa antigen gene (Jiang et al. 2012). This means that the qPCR assays used in this study may not be as sensitive in detecting Rickettsia spp. as the 16S rRNA gene high-throughput sequencing technology using the Illumina platform. The Rickettsia qPCR assays (Henry et al. 2007, Jiang et al. 2012, Maina et al. 2014) are able to detect 3–10 copies of the target DNA fragment per reaction, whereas NGS, using the MiSeq Illumina platform, is more sensitive because of its high sequencing depth of about 1.6 gigabases per run/60 mega bases per hour, so the detection threshold is far above the qPCR assays, especially in cases of multiple pathogen infections.
The inability to amplify rickettsial DNA from blood samples collected from dogs using the Rickettsia genus-specific qPCR and species-specific qPCR assays could also be attributed to several factors. It is possible that rickettsial DNA was present in the samples tested, but at a copy number that was below the detection limits of the assays used (Hawley et al. 2007). Rickettsial DNA may be cleared from the blood of the dogs by a fast and effective immune response as has been hypothesized by (Bayliss et al. 2009). It is also possible that the dogs were infected with rickettsial pathogens, but the organisms were enclosed in other tissues such as the vascular endothelium, dermis, or spleen (Hawley et al. 2007).
An agent closely related to O. tsutsugamushi strain Kawasaki was detected in a dog blood sample. O. tsutsugamushi causes scrub typhus, a febrile disease transmitted by larval-stage mites Leptotrombidium akamushi and Leptotrombidium deliense commonly called chiggers (Parola and Raoult 2006). The disease occurs in the Asia-Pacific region of the world, which includes the north of Australia, India, Korea, Japan, Papua New Guinea, and Pakistan. However, two recent reports indicate that scrub typhus occurs outside this region, one case occurred in a patient visiting UAE and Chile (Izzard et al. 2010, Balcells et al. 2011). Moreover, there have been reports suggesting that scrub typhus occurs in Africa (Osuga et al. 1991, Ghorbani et al. 1997) with the most recent report from Kenya (Thiga et al. 2015).
Infected mites serve as the vector and reservoir of this organism, although the vector/host for the Orientia spp. detected in scrub typhus patients from UAE (Orientia chuto) and Chile (Orientia sp.) is unknown (Izzard et al. 2010, Balcells et al. 2011). Clinical signs of the infection in humans include fever, headaches, maculopapular rash, eschar, lymphadenopathy and neurological signs, cough, and interstitial pneumonia in some cases (Parola and Raoult 2006). There is a recent case report of O. tsutsugamushi infection in a lethargic dog in an area of Japan, showing that dogs can be naturally infected and may play a role as a host of the organism (Namikawa et al. 2014). To our knowledge, this is the first report of an O. tsutsugamushi-like organism in South Africa.
An R. peacockii-like agent was detected in the 16S rRNA gene sequence obtained from the R. sanguineus pool with a 96% sequence identity to R. peacockii and 95.7% similarity to other rickettsiae. R. peacockii, first detected in Dermacentor andersoni ticks in Montana, United States, is generally considered to be a nonpathogenic spotted fever group rickettsia and is mainly transmitted transovarially from one tick generation to the next (Simser et al. 2001). An endosymbiont of D. andersoni ticks, its presence in the tick is related to the reduced prevalence of Rickettsia rickettsii, the cause of Rocky Mountain spotted fever in dogs and humans in the Americas (Felsheim et al. 2009). Phylogenetic analysis of the 16S rRNA gene sequence obtained from the R. sanguineus tick pool against other homologous rickettsial sequences published from the GenBank showed that the sequence was placed between O. tsutsugamushi and an SFGR, but in a separate clade, suggesting that it may potentially be a new species of Rickettsia. Further characterization will, however, be needed to determine if this is indeed a new species of Rickettsia and to determine whether this organism may be pathogenic to dogs and/or humans.
The results of the RLB hybridization assay showed that 49% of the blood samples and 30% of tick pools were positive for the genus-specific probes of Ehrlichia/Anaplasma species. Sequence analysis of the 16S ribosomal RNA gene of four blood samples positive for the Ehrlichia/Anaplasma genus-specific probes revealed a 99% sequence homology to an Anaplasma sp. (South African dog strain) closely related to A. phagocytophilum (Inokuma et al. 2005). In another blood sample positive for Ehrlichia/Anaplasma genus-specific probes, next-generation sequence analysis of the 16S rRNA gene revealed a 99% sequence identity to A. phagocytophilum strains, ApGDr1 (KC800963), ApGDr2 (KC800964), and GDR4 (KC455366). A. phagocytophilum was first detected in dogs in the United States in the 1980s (Madewell and Gribble 1982) with several genetic variants subsequently described based on molecular typing.
Ixodes spp. ticks are the vectors implicated for transmitting A. phagocytophilum (Woldehiwet 2010). However, in this study, only one of 103 ticks collected from domestic dogs was identified as an unspeciated Ixodes. This suggests that other genera of ticks may be responsible for the transmission of Anaplasma spp. to dogs in Bushbuckridge. A. phagocytophilum is a known zoonotic pathogen causing human anaplasmosis (formerly known as human granulocytic ehrlichiosis) with disease in humans characterized by fever, myalgia, headache, an increase in liver function enzymes, thrombocytopenia, and disorientation (Bakken and Dumler 2008). Anaplasma spp. are transmitted to dogs and humans by tick vectors (Nicholson et al. 2010). The diseases caused by Anaplasma spp. can be prevented by effective tick control coupled with awareness of ticks and tick bite prevention (Nicholson et al. 2010). Doxycycline is the drug of choice for treatment of A. phagocytophilum infections (Nicholson et al. 2010). In the present study, almost half of the samples were positive for Anaplasma spp. as confirmed by sequencing. The finding of A. phagocytophilum suggests a potential risk of human infection with this rickettsial pathogen in the study area.
Conclusions
The detection of rickettsial agents, Ca. R. asemboensis and an O. tsutsugamushi-like agent, in canine blood suggests that dogs may play a role in the life cycle of rickettsiae. Our study provides preliminary information about the occurrence of zoonotic rickettsiae in domestic dogs and their ectoparasites in a South African rural community and highlights the potential risk of human infection with these pathogens. Further work is needed to characterize Anaplasma sp. South Africa dog strain and Ca. R. asemboensis to determine their pathogenic potential.
Footnotes
Acknowledgments
The authors would like to thank Dr. Luther van der Mescht of the University of Stellenbosch for ectoparasite identification and the National Research Fund (NRF) South Africa for the research funds used in the study. The authors would also like to thank Dr. Anne Conan of the Ross University School of Veterinary Medicine for the map showing the sampling area in relation to the surrounding game reserves. The authors are grateful to the students from the veterinary schools of Norway, Pretoria, and Utrecht, and to the environmental Monitors of the Mnisi Community Programme, University of Pretoria, who assisted with the sample collection from dogs. The study was approved by the University of Pretoria Animal Ethics Committee (protocol no. VO48-13). The HDSS-Dogs platform (protocol no. VO33-11) was supported by funding to Darryn Knobel from the Morris Animal Foundation, United States (grant no.D12CA-312). This publication has not been reviewed or endorsed by the Foundation, and the views expressed herein do not necessarily reflect the views of the Foundation, its officers, directors, affiliates, or agents. Drs. A.N. Maina and A.L. Richards were supported by funding of the Global Emerging Infections Surveillance and Response System, work unit # A1402. The opinions and assertions contained herein are the private views of the authors and are not to be construed as official or reflecting the views of the Department of the Navy, Department of Defense, or the U. S. government. This is the work of a U.S. government employee (ALR) and may not be copyrighted (17 USC 105).
Author Disclosure Statement
No competing financial interests exist.