
Abstract
From 1996 through 2013, 54,546 individual birds comprising 152 species and 7 orders were banded, bled, and released at four study areas within California, from which 28,388 additional serum samples were collected at one or more recapture encounters. Of these, 142, 99, and 1929 birds from 41 species were positive for neutralizing antibodies against western equine encephalomyelitis virus (WEEV), St. Louis encephalitis virus (SLEV), or West Nile virus (WNV) at initial capture or recapture, respectively. Overall, 83% of the positive serum samples were collected from five species: House Finch, House Sparrow, Mourning Dove, California Quail, and Western Scrub-Jay. Temporal data supported concurrent arbovirus surveillance and documented the disappearance of birds positive for WEEV in 2008 and SLEV in 2003 and the appearance of birds positive for WNV after its invasion in 2003. Results of these serosurveys agreed well with the host selection patterns of the Culex vectors as described from bloodmeal sequencing data and indicated that transmission of WNV seemed most effective within urban areas where avian and mosquito host diversity was limited to relatively few competent species.
Introduction
T
WNV invaded California through the southeastern deserts during 2003 (Reisen et al. 2004), just 5 years after being discovered in New York City (Lanciotti et al. 1999), spread to every county in the state by the end of 2004 (Hom et al. 2004), and has remained continually active since this time, with human disease detected annually (Feiszli et al. 2013). WEEV and SLEV were not detected in mosquitoes from 2007 and 2003, respectively, through 2013, despite the continued testing of mosquito samples (pools) for all three viruses (Fig. 1). Widespread vaccination and natural immunization have limited the number of equine cases of both WEEV and WNV diseases reported annually.
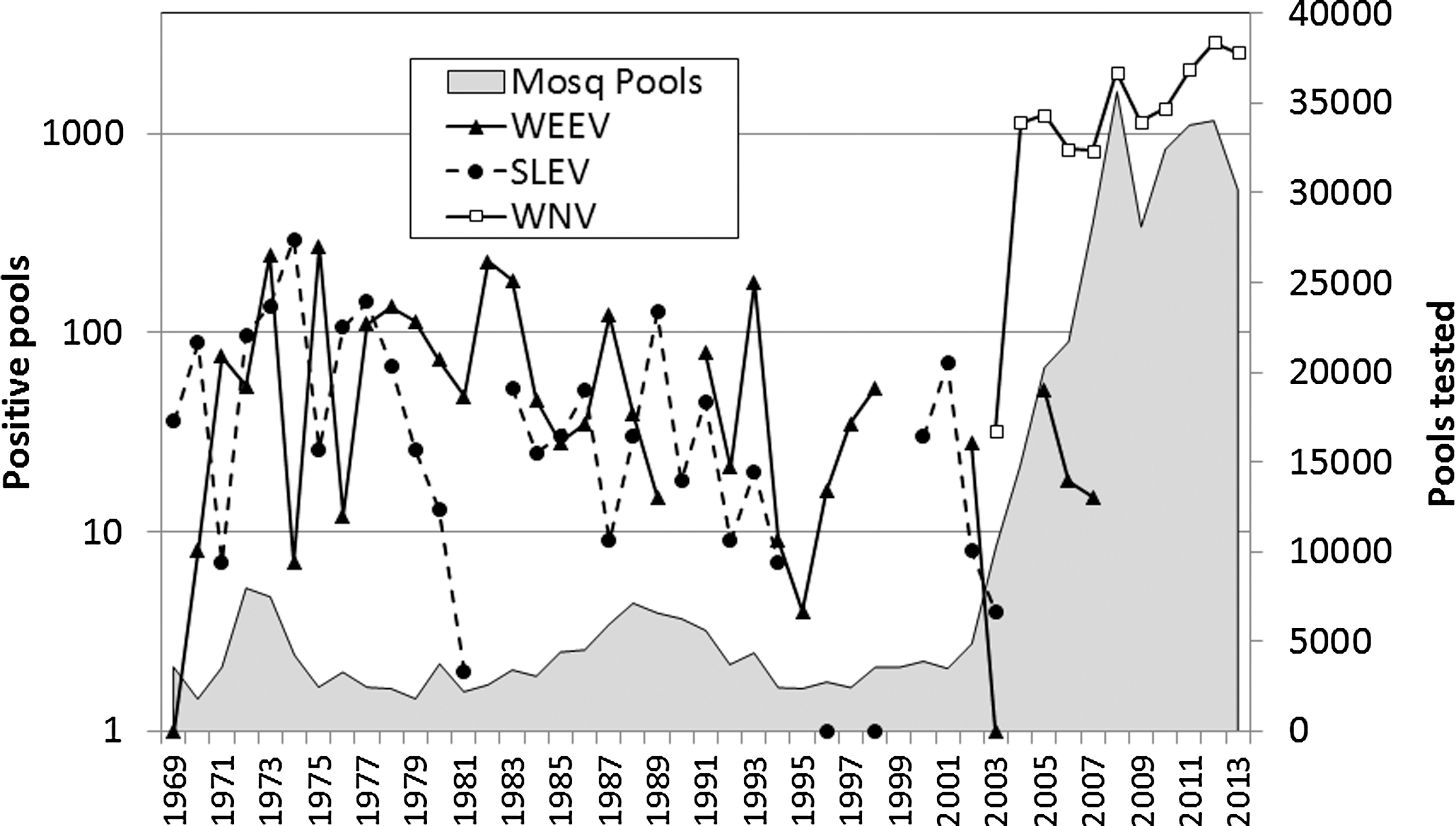
Number of pools of mosquitoes tested and positive for WEEV, SLEV, and WNV from 1969 through 2013. Data summarized from the State of California Arbovirus Surveillance Program show consistent activity of WEEV and SLEV until after the invasion of California by WNV in 2003 (Hui et al. 1999, Steinlein et al. 2003). SLEV, St. Louis encephalitis virus; WEEV, western equine encephalomyelitis virus; WNV, West Nile virus.
The cascade of events that have led to the above transitions in arbovirus activity remain unclear, although the California landscape has changed dramatically since the 1930s. The state human population has grown from ca. 7M in 1930 to 37M by 2010, with extensive urbanization of the Los Angeles Basin and the eastern side of the San Joaquin Valley. Extensive intermittent wetlands of the Central Valley have been converted to farmland, and vernal runoff is now managed by engineering projects along the western slope of the Sierra Nevada. Concurrently agricultural production has risen from $7.3 billion in 1974 to $45 billion/year in 2011, even though some of the original farmland was converted to housing. This increase in farm production followed the completion of the extensive Central Valley Water Project bringing Sacramento River water into the arid west side of the San Joaquin Valley through several major canal systems and to Los Angeles through the California Aqueduct (
Birds are the vertebrate hosts for these three encephalitis viruses and therefore have been studied intensively since their role in transmission was discovered in the 1940s (Hammon et al. 1943). Extensive field studies during 1946–1951 in Kern County carefully characterized bird communities in rural, agrarian, and periurban habitats, documented their seasonality and migration status, and described their nesting habits (McClure et al. 1962). A subset of these birds were tested for antibodies using a mouse protection neutralization assay, of which 17% (n = 1147) and 9% (n = 1039) were positive for WEEV and SLEV antibodies, respectively (Reeves and Hammon 1962). Of the bird species tested, House Finches, House Sparrows, and Bullock's Orioles were positive most frequently. The adaptation of the hemagglutination inhibition assay allowed for rapid and extensive testing of wild birds to detect the prevalence antibodies. During 1952–1973 in Kern County and 1969–1973 in Butte and Glenn Counties, a total of 14,713 blood samples from 131 species of wild birds were tested for antibodies, of which 4% were positive for WEEV and 3% for SLEV, respectively (Milby and Reeves 1990), a marked decline from the previous survey, although resident peridomestic passerines again emerged as the most frequently infected species.
Our studies on the role of birds in the ecology of arboviruses commenced at wetlands along the Salton Sea in the Coachella Valley in 1996, following our discovery that WEEV and SLEV seemed to overwinter in this area and then disperse upland in the Coachella Valley (Reisen et al. 1995b, 1995c) and perhaps to other areas of California. The combination of agricultural, desert, and wetland habitats in this area along the Pacific Flyway has enabled one of the most diverse avifauna in North America, with >380 species listed at the Salton Sea National Wildlife Refuge (
Materials and Methods
Bird collection areas were selected near wetlands around the Salton Sea in Coachella Valley, Riverside County, in the San Joaquin Valley near Bakersfield, Kern County, and in the Sacramento Valley near Davis, Yolo County, and at multiple urban sites throughout Los Angeles. The locations of frequently used collection sites are summarized in Table 1. Additional sites were tried and then discontinued because of low collection success, vandalism, or other logistical reasons. Multiple net lanes and bird trap sites were tried at each location until productive positions were selected for repeated sampling. For netting, three sites at each of the three areas were selected each season and sampled on two consecutive days during each 2-week period, depending upon weather. Each site contained from 7 to 15 mist net locations and multiple small and large grain-baited traps, except for Los Angeles sites where a single modified crow trap was operated at each of seven fixed sites. Mist nets were mostly 2.6 × 12 meters with 38-mm mesh (some with 60-mm mesh were used to collect large birds) and were operated starting at dawn for 3–5 h each day, depending upon catch size and weather. Australian crow traps with drop-in top slits modified for sparrows and/or walk-in bottom cone entrances (McClure 1984) were baited with mixed wild birdseed and water and operated for 24 h for 2 days at each location during each sampling rotation. Additional bird serum samples were collected opportunistically, including samples from waterfowl at the Kern NWR during an avian cholera outbreak, birds taken to wildlife rehabilitation centers, heron chicks sampled at a heronry near Davis, and birds taken under depredation permits by aquiculture, pigeon, and vineyard bird removal programs.
Birds were identified to species and aged using plumage and/or skull ossification (Pyle 1997) and banded using appropriate USGS bands (
Samples were clarified by centrifugation and then stored at −20°C until screened for antibody by an enzyme immunoassay (EIA) using a conjugated anti-bird detector antibody (Chiles and Reisen 1998, Ebel et al. 2002) produced commercially in goats by Bethyl Laboratories, Inc. (Montgomery, TX). Each sample was tested in three wells using a 96-well plate format—two wells with crude viral antigen and one without antigen. Separate plates were precoated with WEEV or SLEV antigen. Using this EIA, birds experimentally infected with WNV or SLEV produced antibodies equally reactive with our SLEV crude antigen (Fang and Reisen 2006), so SLEV antigen was used to screen serum samples for antibody against both flaviviruses. Samples with the ratio of the mean absorbance of the two antigen-positive wells divided by the antigen-negative well that were ≥2 were confirmed by a plaque reduction neutralization test (PRNT). Samples were considered confirmed when endpoint titers on Vero cell culture had >90% reduction of 75–100 plaque-forming units (pfu) of WEEV, SLEV, or WNV at a dilution of ≥1:20. Because SLEV and WNV cross-reacted in our EIA and PRNT assays (Fang and Reisen 2006, Patiris et al. 2008), the purported infecting virus was required to produce an endpoint titer ≥4× the competing virus. Positive bird samples were reported once and were limited to those birds with a confirming PRNT90 titer ≥1:20 or ≥4× the competing virus.
Results
From 1996 through 2013, 54,546 individual birds comprising 151 species and 7 orders were banded, bled, and released, of which 142, 99, and 1929 individuals had serum samples with neutralizing antibodies positive against WEEV, SLEV, or WNV, respectively, at either initial capture or at one or more recapture events (Table 2). Because the number of samples required to detect a positive serum varied among birds, data are presented as the number of individual birds that were detected as positive among all birds sampled. An additional 3732 birds from 137 species were bled, but not banded, of which 16, 39, and 86 were positive for WEEV, SLEV, or WNV, respectively (Table 3). These latter samples came from birds that were not authorized for USGS bands (quail, pigeons), were collected when USGS bands were not available, or were collected for experimental or other purposes. Because they were not banded, some of these birds could have been sampled more than once, confounding attempts to estimate prevalence of infection. Data from Tables 2 and 3 are combined in Figure 2 to show the temporal pattern of seropositive bird collections at Coachella and Kern study sites. Sample sizes reflected changes in landscape related to shoreline changes of the Salton Sea in Coachella Valley and to the flow of the Kern River in Kern County, as well as changes in our staffing and collection methods (arrows show when mist netting was terminated at each site). WEEV and SLEV antibodies detected during 1996–2004 in Coachella disappeared by 2005, two years after the initial detection of WNV in California (Fig. 1). Low-level activity of WEEV in Kern County ended in 2001, was renewed briefly in 2005–2007, ended in 2008, and was not detected through 2013 in birds or by other surveillance methods (Figs. 1 and 2); SLEV was not detected in Kern County throughout the study period.

Number of avian serum samples tested at multiple locations in Coachella Valley
Resident status of birds within study sites based on capture dates and additional information from the Cornell Laboratory of Ornithology website (
M, migrant; S, summer; SLEV, St. Louis encephalitis virus; V, vagrant; W, winter; WEEV, western equine encephalomyelitis virus; WNV, West Nile virus; YR, year round.
Sampling in Los Angeles and Yolo Counties was initiated after the invasion of WNV, and neither WEEV nor SLEV antibody-positive birds were detected at these locations, agreeing with concurrent mosquito and sentinel chicken surveillance. The relatively high proportion of recaptures at these locations (Table 2) reflected the extensive use of grain-baited traps and their attractiveness for passerines; for example, one male House Sparrow was collected and bled 57 times over a 4-year period at the same trap in Los Angeles (Kwan et al. 2012). Data from Los Angeles showed marked temporal variation, with high seroprevalence associated with outbreaks of WNV disease in humans during 2004, 2008, and 2012, when WNV seroprevalence increased to as high as 45% among after hatch-year and 60% among juvenile and hatch-year House Finches and House Sparrows (Kwan et al. 2012). Peak WNV antibody positivity in Kern County during 2007 coincided with the largest outbreak of human disease in this county since the arrival of WNV in 2004 (Reisen et al. 2009a). Interestingly, few human cases of mosquitoborne encephalitis have been reported from Coachella Valley, despite consistent enzootic transmission of all three viruses and an elderly human population associated with several retirement communities.
When sorted by resident status, >95% of seropositive birds and >87% of recaptures were from species classified as year-round residents (Table 4). The remaining positive birds were winter residents that likely were infected during early or late season transmission and summer residents that were infected during the transmission season. Few transient migrants coming north from Mexico or the Neotropics were seropositive, agreeing with our previous study that focused on the role of migratory birds in the movement of arboviruses through this area (Reisen et al. 2010). Some of the species that traversed through the Coachella Valley were summer residents in the Central Valley, and 4 of these 592 birds were seropositive. Overall, 15 of our captures were considered vagrants and were collected outside of their normal ranges in North America (<
Banded birds were summarized by order (Table 5). Overall, 85% of initial captures and 82% of recaptures were in the order Passeriformes, followed by Columbiformes (mostly Mourning Doves) and Galliformes (two species of Quail). Although these latter two orders comprised <20% of the samples from original and recaptured birds, they comprised 49% of the WEEV, 73% of the SLEV, and 33% of the WNV seropositives, perhaps indicating a trend for Culex vectors to blood feed on larger-bodied birds. Adults of these columbiform and galliform birds generally were low to moderately competent hosts based on experimental infection studies (Komar et al. 2003, Reisen et al. 2003, 2005a); however, nestlings were highly competent before their immune systems became fully functional (Mahmood et al. 2004, Reisen et al. 2006c). With the exception of House Finches and House Sparrows, most of the seropositive Passeriformes were fairly large-sized birds such as Western Scrub-Jays. Other corvid species such as the American Crow and Yellow-billed Magpie were infrequently collected and frequently succumbed to WNV infection (Komar et al. 2003, Reisen et al. 2013b, Foss et al. 2015), precluding their utility in serological surveys. Another group frequently positive, but infrequently sampled, was the Pelecaniformes, including the Least Bittern and Black-crowned Night Heron. Some herons and egrets form large multispecies nesting colonies during spring and early summer that may be foci of intense virus transmission (Reisen et al. 2009b), provided these colonies did not form in trees over water (Reisen et al. 2005b). Over land, Culex mosquitoes fed frequently and preferentially on these birds (Thiemann et al. 2011), some of which such as Black-Crowned Night Herons produced elevated WNV viremias as nestlings. Extensive surveillance samples from sentinel or domestic chickens were not included in this report, but serum samples from these birds frequently were positive at all four study areas.
Of the 54,546 birds banded and released, only 128 were reported to us as foreign encounters; that is, reported to the USGS Bird Banding Laboratory by individuals from outside of our research program. Of these, 90 were Mourning Doves, of which 68 were reported as shot and 15 as found dead during the hunting season and relatively near to where they were banded and released. In addition, 12 Western Scrub-Jays and 3 Yellow-billed Magpies were reported as found dead. Five birds were reported from regions outside of California, including a Red-winged Blackbird banded in Bakersfield in 1997 and reported from Alberta, Canada, in 2001, a White-crowned Sparrow banded at the Tracy Ranch in Kern County in 2003 and reported in British Columbia in 2005, a Brown-headed Cowbird banded at the Kern NWR in 1997 and reported from Oregon in 2006, and a Western Wood Pewee banded at the Kern NWR in 1997 and reported from Nevada in 1999. In addition, a Common Yellowthroat banded in Coachella Valley in 1997 was encountered in Santa Cruz on the California Coast in 1999. Interestingly, we did not document the movement of banded birds between our study areas and made only a single foreign band recovery: a Brown-headed Cowbird netted in Lamont near Bakersfield in November 2000 that was originally banded in the San Joaquin Valley (data not available).
Discussion
Disappearance of WEEV and SLEV
Temporal trends in avian seroprevalence agreed well with results from arbovirus surveillance programs in California using standard metrics of human and equine passive case detection and mosquito infection and sentinel chicken seroconversion, and confirmed the disappearance of both WEEV and SLEV from California after 2008 and 2004, respectively, through 2013. Conversely, WNV has remained active each summer after its introduction in 2003, utilizing the same mosquito vectors and generally the same avian hosts as WEEV and SLEV. SLEV reappeared in Coachella Valley, California, and Arizona during the summer of 2015 (unpublished data), but WEEV has not been detected to date. Unfortunately bird sampling ceased in 2013, so the antibody status of the avian community during the reemergence of SLEV is not known.
It was difficult to understand these viral displacements. Possibly very high viremia levels in susceptible avian hosts and moderate Culex vector competence for WNV provided a more effective transmission system than lower avian viremia and greater vector competence by SLEV and WEEV (Hardy and Reeves 1990b, 1990c, Reisen et al. 2005a, Maharaj et al. 2014). Although cross-immunity between SLEV and WNV was demonstrated experimentally in avian hosts (Fang and Reisen 2006, Patiris et al. 2008), SLEV seroprevalence levels in Coachella Valley during 2002 and 2003 (Reisen et al. 2008b) apparently were inadequate to impede the invasion of WNV in 2003 and then all of California by 2004. The establishment of WNV was followed by the immediate and coincidental disappearance of SLEV from California and most of North America (Reisen 2014). Previous infection with WNV imparted sterilizing immunity against SLEV in House Finches (Fang and Reisen 2006) and presumably other birds, and this combined with more efficient WNV transmission may have contributed to the disappearance of SLEV.
Coincidental with the invasion of WNV, WEEV also disappeared from California and North America. WEEV is an alphavirus and WNV is a flavivirus, and therefore there is no cross-immunity. These two viruses utilize many of the same avian hosts, but WEEV viremia was not impaired experimentally by WNV infection when birds were coinfected with both viruses (Reisen and Hahn 2007). Previously, Reeves et al. (1994) speculated that WEEV may disappear due to global climate warming. In support, the median infectious dose of WEEV required to infect field-collected Culex tarsalis was found to increase by two to three orders of magnitude from early spring to midsummer (Hardy et al. 1990a, Reisen et al. 1996b), perhaps indicating the negative impact of warm temperature on susceptibility to infection. However, the last location of active WEEV transmission in California was in the SE deserts, the hottest area of California, where temperatures average 3–5°C warmer than the San Joaquin Valley where WEEV was previously endemic (Reisen et al. 2006b). In addition, WEEV replicates more effectively in mosquitoes at cold temperatures than does WNV (Reisen et al. 1993, 2006b), thereby theoretically extending the transmission season. Previously, it was proposed that floodwater Aedes mosquitoes and mammals (especially Lagomorphs) composed an early season transmission cycle for WEEV that preceded the well-documented Culex–bird cycle (Burton et al. 1966, McLintock et al. 1970). However, our studies in southern California did not support this paradigm as few Aedes were found infected (Reisen 1984, Reisen et al. 1990, 1992a). In addition, we have failed to detect temporal declines in WEEV replicative fitness as indicated by experimental infections in mosquito or passerine hosts (Reisen et al. 2008a) or mosquito and avian cell culture systems (Zhang et al. 2011). Decreased murine neurovirulence was reported for one WEEV strain (IMP181-05) isolated from Imperial County, California, in 2005 (Logue et al. 2009).
The arrival of WNV in California was accompanied by marked wildlife, veterinary, and human health impacts that raised media attention and funding levels for surveillance and mosquito control at the local and national levels. In combination with several drought years during which cropping strategies were altered and irrigation practices modified to increase water-use efficiency, enhanced mosquito control perhaps reduced vector abundance below the entomological thresholds required for efficient avian infection and the amplification of WEEV and SLEV, but not WNV.
Avian host selection
The presence of antibodies against arboviruses in avian hosts depends largely upon the host selection patterns of the mosquito vectors and the ability of the avian host to survive infection and produce detectable and persistent antibodies. Although Cx. tarsalis fed more frequently upon mammals than did members of the Cx. pipiens complex, both vectors blood fed frequently upon a wide variety of birds (Tempelis et al. 1965, Tempelis and Washino 1967, Molaei et al. 2010, Montgomery et al. 2011, Thiemann et al. 2011, 2012, Campbell et al. 2013, Reisen et al. 2013a). Related behavioral studies showed the importance of vegetative ecotones, landscape heterogeneity and composition, and mosquito flight paths during host-seeking flights on the frequency of vector–host contact (Lothrop and Reisen 2001).
A striking example of the importance of landscape was the near absence of pelecaniform bloodmeals from Culex (Thiemann et al. 2012) or antibody in ardeid birds (Reisen et al. 2005b) collected in southern California, where these and other waterbirds such as gulls and cormorants nested and roosted in snags over water or on sand bars. When the same ardeid species nested in trees over land near Davis in the Central Valley, they were frequently infected with WNV (Reisen et al. 2009b) and were the preferred bloodmeal host for Culex (Thiemann et al. 2011).
Based on the current serological data, previous bloodmeal analyses, and dead bird surveillance (Foss et al. 2015), larger birds such as doves, quail, and domestic chickens seemed to be selected more frequently than smaller birds such as warblers and wrens. This agreed with the general overselection of thrushes (family Turdidae) by Culex in eastern United States (Kilpatrick 2011) and Europe (Lundstrom et al. 2001). Adult galliform and columbiform birds generally are poor to moderately competent hosts for WNV (Reisen et al. 2003, 2005a, Wheeler et al. 2009), and along with mammals, their frequent selection by host-seeking mosquitoes most likely dampened virus transmission. Conversely, the corvids (especially American Crows) present an interesting conundrum because they are large-bodied periurban birds, are highly competent for North American strains of WNV and usually die from infection (Komar et al. 2003, Brault et al. 2004), and are frequently associated epidemiologically in time and space with the transmission of WNV to humans (Eidson et al. 2001, Reisen et al. 2006a, Nielsen and Reisen 2007), but seemed to be infrequently fed upon by Culex (Molaei et al. 2010, Thiemann et al. 2012), except under certain conditions of host availability (Thiemann et al. 2011, Campbell et al. 2013). With the exception of Western Scrub-Jays, the high mortality rate among infected corvids and their difficulty in collection made them unsuited for routine serological sampling. House Finches, House Sparrows, and perhaps Oak Titmice seemed to be outliers to this general trend in bird size, because although they are relatively small-sized birds, they frequently were fed upon by Culex (Thiemann et al. 2012), were frequently antibody positive in serosurveys (current and historical studies), produced viremias adequate to infect Culex (Reisen et al. 2005a, 2006b), were linked closely to viral amplification and outbreak patterns in urban settings (Kwan et al. 2012), and frequently tested positive at necropsy when found dead in nature (Foss et al. 2015).
Sampling biases
Although avian serology may be informative and compliment other surveillance measures such as infection prevalence in dead birds for tracking arbovirus activity, our sampling approach and the resulting data had multiple limitations and biases. The diversity of species collected and therefore tested was constrained by sampling. Based on historical studies, we chose to focus on mist netting and grain-baited trap collections that worked well for passeriform, galliform, and some columbiform birds; however, these methods did not collect all species equally and often birds seen within our study areas were rarely collected, making us reluctant to use the numbers captured as an index of abundance.
In addition, these methods did not collect potentially important bird groups such as the psittaforms, anseriforms, and pelecaniforms. Many cities in California now have large resident populations of escaped psittaform birds, and New World species have been found to be highly susceptible to WNV infection (Komar 2003), but with the exception of an occasional Budgerigar, these birds were never collected by our sampling program. The role of anseriform birds in arbovirus amplification remains poorly understood. Although adult mallard ducks were relatively refractory to WEEV and SLEV (low viremia response), 6-day-old ducklings produced peak viremias of 3.3 to 4.3 pfu/mL and 4.4 to 6.7 pfu/mL following needle inoculation with Kern County strains of WEEV and SLEV, respectively, and produced a strong antibody response (unpublished data). Adult mallards infected with WNV produced a peak viremia of 6.7 pfu/mL; ducklings were not tested (Komar et al. 2003). Our few blood samples from anersiforms were taken from birds collected for other purposes and none were antibody positive. Repeated sampling at a heronry consisting of nesting Black-crowned Night Herons, Great Egrets, Snowy Egrets, and Cattle Egrets showed frequent infection of nestlings that had fallen from nests, with elevated viremia and high antibody prevalence (Reisen et al. 2009b), but adult birds in the canopy could not be sampled.
Another sampling problem precluding the estimation of prevalence was that individual birds were sampled on different numbers of occasions. This created variation in the sensitivity of detection of infection among birds within and among species. Obviously birds collected on multiple occasions over several transmission seasons would have a greater probability of being detected as antibody positive than birds collected once in a single occasion. In agreement, after hatch-year birds had a higher prevalence of infection than hatch-year birds.
In addition to biases related to avian species sampling, our data also were limited to those species or individuals that became infected and survived to produce antibody and therefore may not represent infection prevalence. Although most species survive WEEV and SLEV infection (Reisen et al. 2003), many species succumb to WNV infection (Komar et al. 2003, Reisen et al. 2005a, Wheeler et al. 2009). Because the mortality rate from WNV infection varied widely within and among avian species, this contributed to the difficulty in calculating and interpreting seroprevalence data. Therefore, to appreciate the range of avian involvement in WNV transmission, the current study should be compared with the results of concurrent dead bird surveillance (Foss et al. 2015) as well as experimental host competence studies (Wheeler et al. 2009).
Data from foreign recapture encounters were unrewarding. Our sole foreign encounter was a single Brown-headed Cowbird. In addition, we did not encounter banded birds moving among our four study areas, even during migration periods. For example, we banded and released 7294 White-crowned Sparrows and recaptured 2788 that we banded. This winter resident arrived at our study areas during fall and departed in spring, thereby avoiding most of the virus transmission season, although some were found antibody positive for WNV. Many of the birds we collected in the San Joaquin Valley purportedly nest in Alaska (Hardy et al. 1965), a journey of over 3000 miles each way. However, they returned each fall to the exact same wintering grounds, and banded birds were never recaptured at different sampling sites. Most foreign encounters of our banded birds were by hunters during dove season: 83 of 90 encounters of Mourning Doves were shot or found dead. Overall, 12 of 12 Western Scrub-Jay reports were from dead birds that may have succumbed from WNV, but were not submitted for testing. Only five birds were reported from outside of our region. These data certainly indicated the difficulty in understanding bird migration routes and survival, and the low frequency of band encounters along the Pacific Flyway.
Relatively few bird species were repeatedly positive for antibodies against these three arboviruses. We sampled 152 different species of birds; however, 83% of the positive individuals were from 5 key species. From a WNV epidemiological standpoint, information on cumulative seroprevalence among two peridomestic maintenance hosts (House Finches, House Sparrows) sampled by grain-baited traps seemed most informative. In Los Angeles, when herd immunity or the proportion of antibody-positive birds collected during January–March was >20%, the probability of a summer outbreak of human disease was low (Kwan et al. 2012), and these data now are used for early season planning by local mosquito control agencies. In contrast, our attempts to establish similar trapping programs at our Kern County and Coachella Valley study areas were less successful because we failed to establish high trap collection success and trap fidelity (Fig. 2), and seroprevalence rates in these species generally were not predictive of virus amplification (Reisen et al. 2008b, 2009a). Passive surveillance using dead birds reported by the public seemed to be a more cost-effective and efficient method of monitoring WNV activity (Foss et al. 2015).
The intensive and widespread enzootic transmission of WNV to birds and improved genetic methods of bloodmeal host identification provided a more extensive appreciation of encephalitis virus transmission patterns in California than could be extracted from studies when there was low-level activity of WEEV and SLEV. Although these viruses occasionally infected a wide range of avian hosts, only a few species seemed critical for maintaining transmission, and the simplification of diversity in periurban landscapes allowed effective virus amplification to outbreak levels. As suggested previously (Reeves and Hammon 1962), key avian species were abundant and widespread during the summer transmission season in habitats frequented by questing Culex, frequently survived infection, produced elevated viremias following infection with small amounts of virus, had a high reproductive or replacement rate ensuring a continued number of immunologically naïve birds, and were fed upon frequently by Culex vector species. In California, House Finches, House Sparrows, and Western Scrub-Jays seemed to best fit these criteria, although comparable adequate data were not available on all other possible species.
Footnotes
Acknowledgments
The authors gratefully thank VM Martinez, BD Carroll, and staff of the Arbovirus Field Station in Kern County, R Cusack, J Lundstrom, and HD Lothrop in Coachella Valley, S. Kluh, J. Kwan, and staff of the Greater Los Angeles County Vector Control District in Los Angeles, and V. Armijos and S. Wright in Yolo County for help with the collection, banding, and bleeding of birds, and S. Garcia, Y. Fang, R. Chiles, and staff of the Center for Vectorborne Diseases Arbovirus Program for testing serological samples. Data maintenance and summaries were provided by BD Carroll. This research was supported by grants from the Coachella Valley Mosquito and Vector Control District, the Mosquito Research Program of the University of California, and grants, R01AI55607 and RO1AI32939, from the National Institute of Allergy and Infectious Diseases, NIH.
Ethics
The collection and sampling of wild birds was done under protocols approved by the Institutional Animal Care and Use Committee of the University of California, Davis, California, Resident Scientific Collection Permits by the State of California Department of Fish and Game, and United States Department of the Interior U.S. Geological Survey Station Bird Banding permit 22763. The use of arboviruses for neutralization assays was approved under Biological Use Authorization #0554 and #0873 by the Institutional Biosafety Committee of Environmental Health and Safety of the University of California, Davis.
Author Disclosure Statement
No competing financial interests exist.