
Abstract
Introduction:
We previously identified tick salivary lectin pathway inhibitor (TSLPI) in Ixodes scapularis, a vector for Borrelia burgdorferi sensu stricto (s.s.) in North America. TSLPI is a salivary protein facilitating B. burgdorferi s.s. transmission and acquisition by inhibiting the host lectin complement pathway through interference with mannose binding lectin (MBL) activity. Since Ixodes ricinus is the predominant vector for Lyme borreliosis in Europe and transmits several complement sensitive B. burgdorferi sensu lato (s.l.) strains, we aimed to identify, describe, and characterize the I. ricinus ortholog of TSLPI.
Methods:
We performed (q)PCRs on I. ricinus salivary gland cDNA to identify a TSLPI ortholog. Next, we generated recombinant (r)TSLPI in a Drosophila expression system and examined inhibition of the MBL complement pathway and complement-mediated killing of B. burgdorferi s.l. in vitro.
Results:
We identified a TSLPI ortholog in I. ricinus salivary glands with 93% homology at the RNA and 89% at the protein level compared to I. scapularis TSLPI, which was upregulated during tick feeding. In silico analysis revealed that TSLPI appears to be part of a larger family of Ixodes salivary proteins among which I. persulcatus basic tail salivary proteins and I. scapularis TSLPI and Salp14. I. ricinus rTSLPI inhibited the MBL complement pathway and protected B. burgdorferi s.s. and Borrelia garinii from complement-mediated killing.
Conclusion:
We have identified a TSLPI ortholog, which protects B. burgdorferi s.l. from complement-mediated killing in I. ricinus, the major vector for tick-borne diseases in Europe.
Introduction
L
The classical, the lectin, and the alternative complement pathway are all involved in the host defence against B. burgdorferi s.l. (de Taeye et al. 2013). The lectin pathway is activated when mannose-binding lectin (MBL) or ficolins bind to carbohydrates on pathogen surfaces and activate MBL-Associated Serine Proteases (MASPs), leading to cleavage of C2 and C4 resulting in deposition of C3 convertase (C4bC2a). This will initiate the complement cascade, which triggers influx of immune effector cells, opsonophagocytosis, and direct lysis by membrane attack complexes (MAC) (Matsushita et al. 2013). Differences in complement sensitivity between B. burgdorferi s.l. genospecies are related to differential expression of complement regulating outer membrane proteins (Kraiczy et al. 2001, Kurtenbach et al. 2002, Bhide et al. 2005). In addition, Ixodes ticks introduce several salivary proteins, such as Salp15, Salp20, ISAC, and tick salivary lectin pathway inhibitor (TSLPI), which could give complement-sensitive B. burgdorferi s.l. strains a survival advantage during early skin invasion (Valenzuela et al. 2000, Daix et al. 2007, Schuijt et al. 2008, 2011a, Tyson et al. 2008, de Taeye et al. 2013).
We previously identified an I. scapularis protein, which was designated as TSLPI (GenBank acc. no. AEE89466.1), by screening an I. scapularis salivary gland protein library with tick-immune rabbit sera using yeast surface display technology (Schuijt et al. 2011b). TSLPI binds MBL and inhibits the host lectin complement pathway, mainly by inhibiting MBL-ligand binding. TSLPI plays a role in the transmission of B. burgdorferi s.s. from I. scapularis ticks to the murine host, as well as in acquisition from the host to larval I. scapularis ticks (Schuijt et al. 2011a). Moreover, silencing of TSLPI and passive immunization with anti-TSLPI antibodies reduced B. burgdorferi s.s. loads in mice after infection by I. scapularis ticks, revealing its potential as an anti-tick vaccine candidate.
Because I. ricinus is the most important vector for B. burgdorferi s.l. in Europe, we examined the presence of an ortholog of I. scapularis TSLPI in I. ricinus and investigated its effect on the MBL complement pathway and complement-mediated killing of B. burgdorferi s.l.
Materials and Methods
Identification of TSLPI in I. ricinus salivary glands
Fed I. ricinus salivary glands were collected from three adult female ticks laboratory-bred at Utrecht University, and RNA was isolated (Hovius et al. 2007) followed by a reverse transcriptase reaction, as described (Narasimhan et al. 2014). We designed primers A and B (Supplementary Table S1; Supplementary Data are available online at
Quantitative PCR analysis of TSLPI expression during tick feeding
I. ricinus nymphs were fed on a rabbit, and salivary glands of 50 nymphs were collected and pooled at 24 and 72 h of tick feeding. Salivary glands of unfed nymphs were also collected and pooled. RNA was isolated as described (Hovius et al. 2007) and purified using an RNeasy mini kit (Qiagen), and cDNA was produced as described above. A quantitative PCR (qPCR) for TSLPI mRNA expression was performed using primers C and D (Supplementary Table S1). Ixodes ricinus actin primers as described (Hovius et al. 2007) were used as a reference gene target for normalization. qPCRs were performed using the LightCycler 480 (Roche) and SYBR green dye (Roche), and four replicates were performed for each. The PCR protocol was 95°C 6 min, and 50 cycles of 95°C 10 s, 60°C 20 s, and 72°C 20 s. Results were analyzed with LinRegPCR software (Amsterdam, The Netherlands) (Ruijter et al. 2009).
In silico sequence analysis of I. ricinus TSLPI
To assess the homology of the I. ricinus TSLPI mRNA sequence with I. scapularis TSLPI (salivary protein P8, AEE89466.1) and other known genes, we performed a Position-Specific Iterated Search (PSI-BLAST) through the NCBI website.
Recombinant protein expression
To produce glycosylated I. ricinus rTSLPI, the signal sequence was identified with Signalp 3.0 (Bendtsen et al. 2004) and PCR cloning with BGL-II and Xho1 restriction sites was performed, using Primers E and F (Supplementary Table S1) and High-Fidelity SuperMix (Invitrogen) under the following conditions: 30 s 94°C; 25 cycles of 30 s 94°C; 50 s 60°C; 25 s 72°C, followed by 5 min at 72°C. I. ricinus TSLPI was cloned into the pMT-Bip-V5-His tag vector (Invitrogen), which was transformed into Drosophila S2 cells together with pCO-Blast. Recombinant protein was expressed and purified using the Drosophila Expression System (Invitrogen) as described earlier (Schuijt et al. 2011b, 2013). After 8 weeks of blasticidin selection, the Drosophila cell culture was induced using copper sulfate for 3 days. The supernatant was filtered (0.45 μm) and run over a Ni-NTA column (Pierce) according to manufacturer's protocols using 250 mM imidazole for elution. The elution was filtered (0.22 μm), concentrated with a 9 kDa protein concentrator (Pierce), and washed with sterile PBS three times to eliminate residual imidazole. The protein concentration was assessed by a DC-Protein-Assay (Bio-Rad). Purity was checked on 18% sodium dodecyl sulfate (SDS) gel using Coomassie blue staining.
MBL-dependent C4 deposition assay
High-binding microtiter plates (Microlon; Greiner) were coated with 10 μg/mL mannan (Sigma) in a coating buffer (15 mM Na2CO3, 35 mM NaHCO3, 15 mM NaN3; PH 9.6) overnight at 4°C. Wells were blocked using a blocking buffer (10 mM Tris-HCl, 145 mM NaCl, 15 mM NaN3, 1 mg/mL bovine serum albumin [BSA]) at room temperature (RT) for 2 h. After washing with TBS/0.05% Tween 20/5 mM CaCl2, normal human serum (NHS) diluted 512× was supplemented with I. ricinus TSLPI, I. scapularis TSLPI, or BSA diluted in C4 activation buffer (4 mM Na-diethyl-barbiturate, 145 mM NaCl, 2 mM CaCl2, 1 mM MgCl2, 0.02% Tween 20, 1 mg/mL BSA), and samples were incubated for 45 min at 37°C. MBL-deficient serum (<0.02 μg MBL/mL serum) with BSA was used as a negative control. Wells were washed and incubated with biotinylated monoclonal mouse anti-human C4 IgG (0.25 μg/mL) for 1 h at RT. After washing, streptavidin-conjugated horseradish peroxidase (HRP) diluted 1:10,000 in a washing buffer was incubated for 30 min at RT. After washing, 50 μL TMB substrate and 10 μL 3% H2O2 in NaAc buffer were added and absorbance was measured at 450 nm (optical density at 655 nm subtracted) using an iMark Microplate Reader (Bio-Rad).
Complement-mediated killing assay
We performed direct complement-mediated killing assays as described previously (Schuijt et al. 2011a, Wagemakers et al. 2014). Briefly, we added 0.1 to 1.0 μg/μL of I. ricinus rTSLPI or 1.0 μg/μL BSA to 25% NHS, incubated at 37°C for 30 min, and added this to 2.5 × 105 intermediate complement-sensitive (Kraiczy et al. 2000) B. burgdorferi N40 spirochetes grown to log-phase in a sterile V-shaped enzyme-linked immunosorbent assay (ELISA) plate (Greiner). Heat-inactivated human serum (HIS) (by incubation at 56°C for 30 min) was used as a control. The plate was incubated for 90 min, wells were resuspended, and 5 μL samples were examined by dark-field microscopy by a researcher blinded to the experimental design, and 100 spirochetes in each sample were scored as either motile or immotile. Complement-sensitive B. garinii strain A87S was incubated with 12.5% NHS or HIS, and samples were examined after 60 min of serum incubation.
Statistics
Data are represented as mean ± SEM. Comparisons were made using a Student's t-test in GraphPad Prism 5.0, with Statistical significance: p ≤ 0.05 (*), p ≤ 0.01 (**), and p ≤ 0.001 (***).
Results
A TSLPI ortholog is expressed in I. ricinus ticks and is upregulated during tick feeding
To identify a TSLPI ortholog, a PCR on I. ricinus salivary gland cDNA with primers based on the I. scapularis TSLPI signal sequence and 3′UTR resulted in a 315 bp product, which was ligated into a pGEMT-easy vector and eight clones were sequenced. The TSLPI sequence in I. ricinus was based on five identical clones and submitted to GenBank (acc. no. KF451926). Mature I. ricinus TSLPI is encoded by a 210 nucleotide coding region, which is 93% and 89% identical to I. scapularis TSLPI at the mRNA level and amino acid level, respectively. Next, we measured TSLPI mRNA expression in I. ricinus salivary glands during tick feeding to explore its significance in the feeding process. Similar to its ortholog in I. scapularis ticks (Schuijt et al. 2011a), I. ricinus TSLPI mRNA expression was not present in unfed nymphs and was upregulated during the course of feeding (Fig. 1A). In silico analysis using a PSI-BLAST revealed various homologous salivary proteins in I. scapularis and I. pacificus (Fig. 1B), including Salp9 putative anticoagulant (93% homology), anticoagulant Salp11 (90% homology), Salp14 (81% homology), and basic tail salivary proteins (BTSPs, e.g., 80% homology to BTSP-4).

Ixodes ricinus expresses a TSLPI ortholog during tick feeding.
Notably, factor Xa inhibitor Salp14 also appears largely homologous to TSLPI, but has an additional C-terminal basic tail. It therefore appears that I. ricinus TSLPI is part of a larger family of proteins across multiple Ixodes species.
Functional characterization of I. ricinus TSLPI
To investigate the function of I. ricinus TSLPI, we produced I. ricinus rTSLPI in a Drosophila expression system, resulting in a ∼16 kDa protein (Fig. 2A). I. ricinus TSLPI showed a dose-dependent ability to inhibit MBL-dependent C4 activation, similar to its counterpart in I. scapularis, further confirming their functional homology (Fig. 2B). To investigate whether I. ricinus TSLPI inhibits complement-mediated killing of B. burgdorferi s.l., we performed in vitro killing assays in the presence of I. ricinus rTSLPI, in essence as previously described (Schuijt et al. 2011a). As expected, I. ricinus rTSLPI dose-dependently protected B. burgdorferi strain N40 from complement-mediated killing by NHS (25%) (Fig. 2C). Complement-mediated killing of the complement-sensitive strain B. garinii A87S by NHS (12.5%) was also diminished in the presence of I. ricinus TSLPI (Fig. 2D).
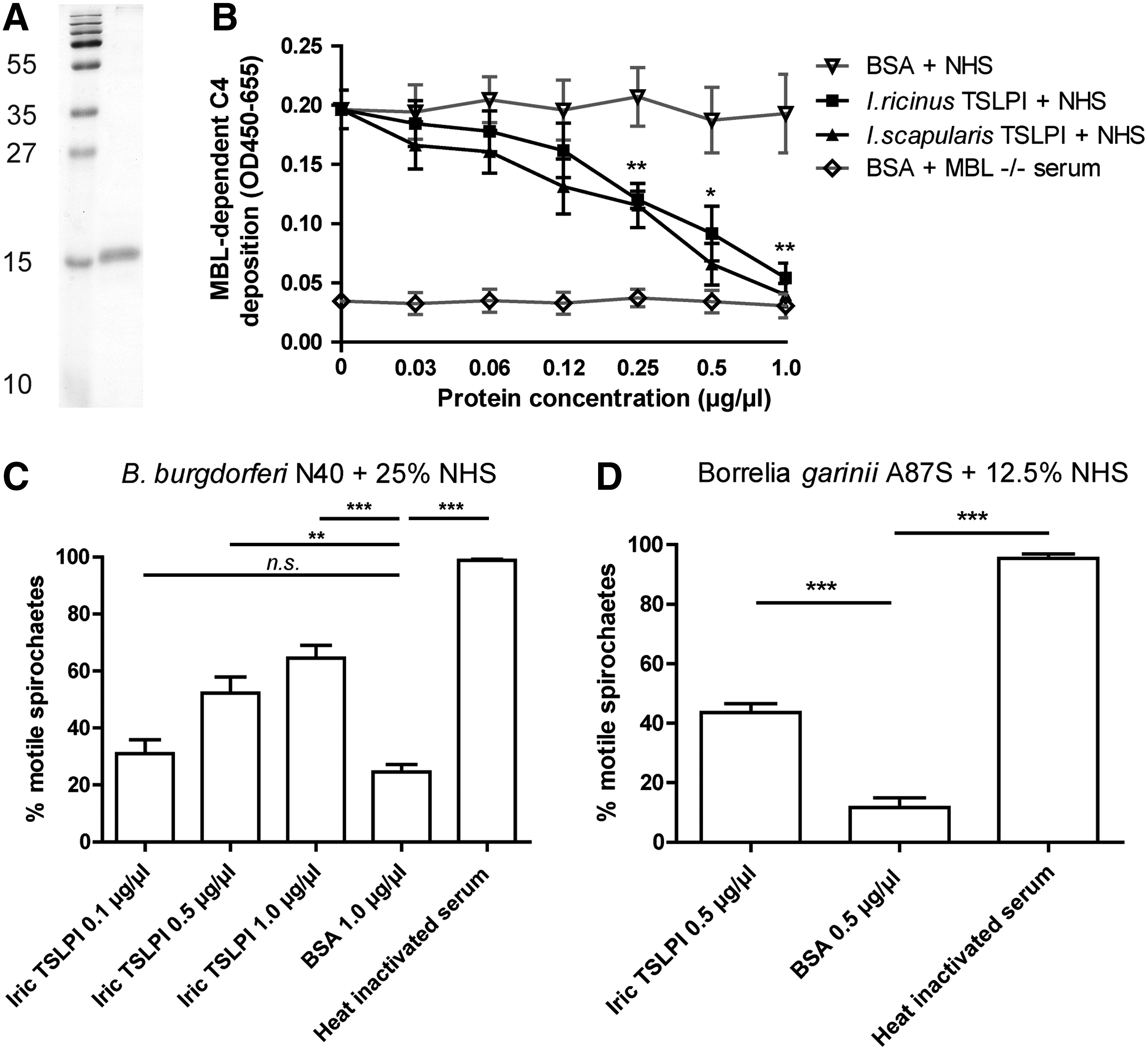
I. ricinus rTSLPI protects Borrelia burgdorferi s.l. from complement-mediated killing.
Discussion
We have identified an ortholog of TSLPI in I. ricinus ticks and demonstrated that it is part of a larger family of Ixodes salivary proteins, comprising Salp9pac, Salp14, and BTSPs, among others. In addition, we showed that I. ricinus TSLPI is upregulated during I. ricinus tick feeding and that it inhibits the lectin pathway and thus protects both B. garinii and B. burgdorferi s.s. spirochetes from killing by the human complement system.
We showed for the first time that I. ricinus TSLPI aids B. burgdorferi s.l. to evade the host lectin complement system. Previously, other mechanisms have been described by which I. ricinus is able to inhibit the host complement system, namely by ISAC and IRAC, which inhibit the alternative pathway, and by Salp15, which by binding to its outer surface protects B. burgdorferi s.l. from MAC formation (Schroeder et al. 2007, Couvreur et al. 2008, Schuijt et al. 2008). In I. scapularis saliva, Salp15, Salp20, Isac, and TSLPI have been previously identified as anticomplement proteins (Valenzuela et al. 2000, Schuijt et al. 2008, 2011, Tyson et al. 2008). These proteins, in combination with its intrinsic complement evasion techniques, enable B. burgdorferi s.l. to evade killing by the host complement system (Pausa et al. 2003, Kraiczy et al. 2004, Pietikainen et al. 2010, de Taeye et al. 2013, Koenigs et al. 2013). Only two of the TSLPI/Salp14 family have been previously described in more detail; TSLPI (a lectin complement pathway inhibitor) (Schuijt et al. 2011a) and Salp14 (a factor Xa inhibitor) (Narasimhan et al. 2002, 2004). It is currently unknown whether TSLPI paralogs exert anticomplement activity. TSLPI, however, does not influence the coagulation cascade in contrast to Salp14, suggesting that the basic C-terminus of Salp14 is involved in factor Xa inhibition (data not shown). Moreover, the striking homology between TSLPI and several other Ixodes saliva proteins suggests that antibodies against TSLPI, which have previously been demonstrated to inhibit B. burgdorferi transmission by ticks, might also be directed against several of its paralogs. This mechanism might add to the effectiveness of TSLPI as the basis for an anti-tick vaccine candidate, and we are currently exploring different approaches to use TSLPI as an anti-tick vaccine.
Conclusions
I. ricinus ticks express TSLPI upon feeding, which is able to protect B. burgdorferi s.l. spirochetes from complement-mediated killing. TSLPI appears to be part of a larger protein family conserved among different Ixodes species.
Footnotes
Acknowledgment
This work was supported by a VENI stipend (91611065) from the Organization for Health Research and Development (ZonMw) in the Netherlands.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.