
Abstract
Highlands J virus (HJV) is an alphavirus closely related to western equine encephalitis virus (WEEV) and eastern equine encephalitis virus (EEEV). HJV is an avian pathogen with the potential for disruption of poultry operations, but is not known to cause human or equine disease. HJV has only been identified in the eastern United States and is thought to have a transmission cycle similar to that of EEEV involving Culiseta melanura mosquitoes and birds. However, HJV is more genetically similar to WEEV and it remains unclear if it may be transmitted by Culex species mosquitoes like WEEV. Seven strains of HJV were characterized to assess this potential. Phylogenetic analysis of whole genome sequences revealed four distinct HJV lineages (lineages 1–4), and vector competence studies in Cx. tarsalis with four of the HJV strains from different lineages yielded two distinct infection patterns. Lineage 1 strains had low infection rates, while lineages 2 and 4 had significantly higher infection rates similar to those previously published for WEEV. The average mosquito body viral titer was highest at 8 dpi (6.60–7.26 log10 pfu equivalents/body), and head titers at all time points ranged between 6.01 and 6.80 log10 pfu equivalents/head. Nearly 45% of mosquitoes infected with strain AB-80-9 were able to transmit virus in saliva with an average titer of 5.02 log10 pfu equivalents/saliva. A single amino acid difference between high and low infectivity phenotypes was identified at genome position 8605, in the E2 gene. A nonpolar glycine was present in the low infectivity lineage 1 strains, while an acidic glutamic acid was present in the higher infectivity lineage 2 and 4 strains. This study demonstrates HJV transmission by Cx. tarsalis mosquitoes and clearly identifies the potential for transmission in the western United States. Two infection phenotypes were exhibited, indicating the need for further studies to understand Culex species transmission patterns.
Introduction
H
Unlike WEEV and EEEV, HJV is not known to cause illness in mammalian hosts with the single exception of the death of an equine in 1964 (Jennings et al. 1966). Concurrent St. Louis encephalitis virus (SLEV) and HJV infections were reported in humans in 1990–1991; however, all symptoms of disease were attributed to SLEV (Meehan et al. 2000). In contrast to the lack of disease in mammals, HJV is an acknowledged avian pathogen with potential to significantly disrupt commercial production flocks. Experimental infection of young turkeys results in depression, somnolence, and leg paralysis, with nearly one-third of the animals dying between days 2 and 3 postinfection (Guy et al. 1993). Permanent egg production decreases of at least 50% have been reported in HJV-infected birds with surviving birds often laying small, soft, or shell-less eggs (Wages et al. 1993). Infected chukar partridges were stricken with leg weakness, tremors, and erratic behavior with morbidity and mortality reaching nearly 35% in one outbreak (Eleazer and Hill 1994).
Unlike EEEV, which has been identified in both North America and the Caribbean (Arrigo et al. 2009), HJV has only been identified in eastern North America. The primary vector for HJV is presumed to be the mosquito Culiseta melanura, with passerine and wading birds serving as the main amplifying hosts (Hayes and Wallis 1977, McLean et al. 1985). HJV has also been isolated from numerous additional mosquito species, including Aedes canadensis, Aedes cantator, Aedes vexans, Culiseta morsitans, Culex pipiens (Andreadis et al. 1998), Aedes infirmatus, Aedes atlanticus-tormentor (Wellings et al. 1972), and Aedes taeniorhynchus (Henderson et al. 1962), demonstrating the potential for spread of HJV outside of the confines of the geographic range of Cs. melanura.
Although HJV has never been isolated from Cx. tarsalis nor has it been identified in the western United States, the close antigenic relationship between WEEV and HJV leads to speculation about whether the primary vector of WEEV, Cx. tarsalis, could also transmit HJV. No published studies of HJV transmission by Cx. tarsalis exist; however, the isolation of HJV from Cx. pipiens indicates that Culex species could be capable of HJV transmission. This current study was undertaken to assess the ability of Cx. tarsalis to transmit HJV and to identify viral genetic elements involved in modulation of Cx. tarsalis vector competence.
Materials and Methods
Virus propagation
Seven strains of HJV were obtained from the CDC Arboviral Reference Collection in Fort Collins, CO. The HJV strains used (Table 1) were passaged at a multiplicity of infection (moi) of 0.01 on Vero cells to create stock cultures. Supernatants were harvested at 48–72 h when ∼50% of the cells exhibited cytopathic effects (CPE). Supernatants were diluted 1:1 in heat-inactivated fetal bovine serum (FBS) and stored at −80°C. Virus titer was determined by plaque assay as previously described (Miller and Mitchell 1986).
Number of passages in each cell type.
C, chick; DE, duck embryo; HJV, Highlands J virus; P, passage in unknown cell type; SM, suckling mouse brain; V, vero; WC, wet chick; WEEV, western equine encephalitis virus.
Virus sequencing
Primers (Supplementary Table S1; Supplementary Data are available online at
Phylogenetic analysis
A ClustalW (MEGA 5) nucleotide alignment was performed on full length genomes as previously described (Logue et al. 2009). Alignment was followed by Maximum likelihood analysis using the Tamura-Nei model for nucleotide substitutions with the ML Heuristic nearest-neighbor interchange method. Bootstrapping with 500 replications was used as the test of phylogeny.
Mosquito feeds, processing, and CPE
Mosquito feeds were performed in duplicate as previously described (Saxton-Shaw et al. 2013) using Cx. tarsalis from the Kern National Wildlife Refuge (kindly provided by Dr. Bill Reisen). Briefly, 4-day-old females were provided a bloodmeal consisting of equal parts washed goose erythrocytes, 10% sucrose in FBS, and virus suspension at a final concentration of 5.5 log10/mL. Mosquitoes were allowed to feed for 1 h at which time fully engorged mosquitoes were transferred to 28°C incubators. At days 4, 8, and 11, each mosquito head was separated from the body and individual parts were placed in 1.7 mL tubes with 400 μL of Dulbecco's modified essential medium (DMEM) supplemented with 100 mL FBS, 1 mL of Gentamycin (50 μg/mL), and 1 mL of Fungizone (1 μg/mL). Tissues were ground using microfuge pestles (Kontes/Kimble Chase), and each homogenate was passed through a 0.2 μM nylon Acrodisc 13 mm syringe tip filter (Pall). Filtrates were tested for CPE on Vero cells held at 37°C with 5% CO2 for 4 days. The percent infected was calculated as the number of CPE-positive bodies of the total number of bodies processed. Percent dissemination was calculated as the number of CPE-positive heads of the number of CPE-positive bodies.
Mosquito saliva collection
Mosquitoes were provided a bloodmeal of HJV strain AB-80-9 as above and held for 8 days at which time they were allowed to salivate into a glass capillary tube charged with 5 μL of Type B Immersion Oil (Cargille) for 1 h (Aitken 1977, Beaty and Aitken 1979). After salivation, the mosquito heads and bodies were processed as described above. The oil-filled end of the tube was broken off into 1.7 mL tubes containing 450 μL of DMEM and the tubes were centrifuged at 4500 × g for 5 min. Saliva samples were stored at −80°C until RNA extraction was performed.
Virus quantitation by real-time PCR
One-step quantitative real time RT-PCR (qRT-PCR) was performed on mosquito heads, bodies, and saliva samples to determine virus titers. Ten CPE-positive and two CPE-negative head and body pairs were chosen from each feed. RNA was extracted as above from 100 μL of each filtrate. Samples were eluted in 100 μL buffer AVE. The qRT-PCR reactions were performed in duplicate using a protocol based on the Qiagen QuantiTect probe (TaqMan) RT-PCR Kit. Average titer, reported as pfu equivalents per sample, was calculated comparing sample Ct values to a standard curve and multiplying the value from qRT-PCR by the dilution factors used to generate the sample as previously described (Logue et al. 2009).
Statistics
Statistical analysis comparing replicates was performed using Pearson p values, which were between 0.17 and 1.00, indicating that there were no statistically significant differences between the duplicate feeds or the combined dataset and individual feeds; the combined dataset is presented in this study. Pearson Chi-square and p value statistics were calculated using 2 × 2 Contingency Tables to determine if infection and dissemination rate differences were statistically significant. Fisher Exact Probability Tests (two tailed) were also calculated.
Results
Before this study, limited genetic information was available for HJV; only three complete genome sequences were available in GenBank. To further examine the genetic diversity among HJV strains, seven strains were chosen representing diverse locations, year of isolation, and isolation source for complete genome sequencing. Of the seven strains chosen, 64A-1519 was genetically most similar to the prototype strain, B230, with only 22 nucleotide differences, 17 of which were synonymous (Tables 2 –4). Strain WX3-2AP was the most divergent from B230 with a total of 167 nucleotide substitutions, eight of which resulted in a change of amino acid. The 20 total amino acid differences between B230 and all other strains were distributed throughout virtually all viral genes, but the majority was located in nsP2 and nsP3 with six mutations each. Thirteen of the amino acid mutations involved a shift that retained the characteristics of the amino acid (e.g., polar to polar), while seven mutations involved a change in characteristics. Eleven amino acid mutations were present in only a single strain, with the remaining nine mutations shared by at least two strains.


Highlighted and bolded amino acids are different from those present in strain B230.
Phylogenetic analysis revealed four distinct lineages of HJV (Fig. 1). Lineage 1 consists of strains B230 and 64A-1519, isolated in the 1960s in Florida. Lineage 2 consists of RU-M-80-259, AB-80-9, and 73V-2540, all isolated between 1973 and 1980 in the northeastern United States. Lineage 3 includes strains 744-01, 585-01, and W17791, which form a sister clade with lineage 2. Strain WX3-2AP is the only member of lineage 4, which is the most divergent lineage identified. WEEV (Imperial 181 strain) was used as outgroup and supports the monophyletic nature of HJV.
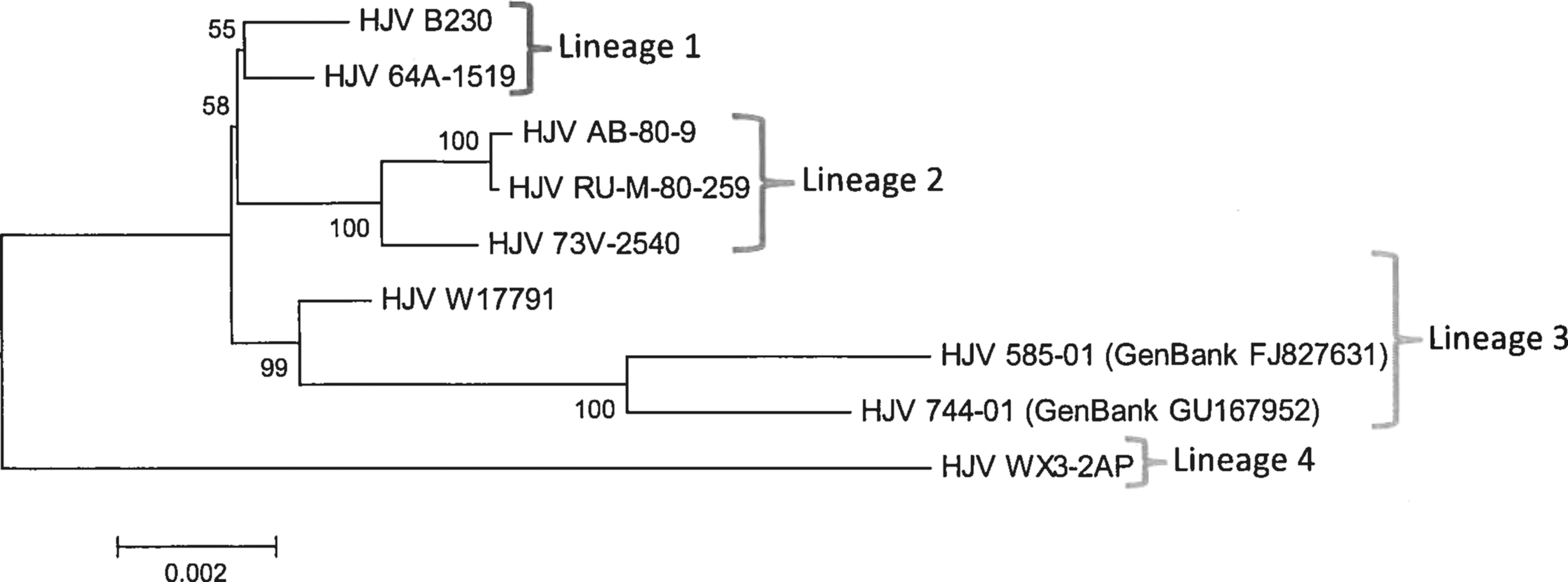
Maximum likelihood phylograms for HJV. Numbers at the nodes indicate bootstrap values based on 500 replications. HJV, Highlands J virus.
Initial characterization of HJV mosquito infection patterns in Cx. tarsalis focused on the two most genetically divergent strains: B230 and WX3-2AP. Strain B230 infection rates were low, ranging from 39.7% at 4 days postinfection (dpi) to 27.6% at 11 dpi (Table 5). Dissemination rates ranged between 67.7% and 84.6%, indicating that once B230 was able to infect the mosquito midgut, the virus was able to establish a disseminated infection. In contrast, strain WX3-2AP exhibited both high infection and dissemination rates, equal to those rates previously published for WEEV strains (Mossel et al. 2013). Infection rates were already high (72.6%) at 4 dpi reaching 78.5% at 11 dpi, and dissemination rates likewise increased from 47.2% at 4 dpi to 90.2% at 11 dpi. There is a highly significant statistical difference between the infection rates of the two strains (4 dpi bodies p≤0.0001, 8 dpi bodies p≤0.0001, and 11 dpi bodies p≤0.0001), but no statistically significant difference in the dissemination rates (4 dpi heads p = 0.07, 8 dpi heads p = 0.65, and 11 dpi heads p = 0.12).
Because disparate mosquito infection phenotypes were initially identified, the lineage 2 strain AB-80-9 was added to aid in determining which of the two phenotypes was more common among HJV strains. Strain AB-80-9 exhibited high infection rates of 84.0% at 4 dpi and 73.0% at day 11, similar to WX3-2AP levels (8 dpi bodies p = 0.25 and 11 dpi bodies p = 0.47). Dissemination rates rose from 44.4% at 4 dpi to 95.7% at 11 dpi and were statistically indistinct from WX3-2AP dissemination rates (4 dpi heads p = 0.76, 8 dpi heads p = 0.25, and 11 dpi heads p = 0.30). Both AB-80-9 and WX3-2AP started with low dissemination rates that increased to nearly 95% by day 11, while B230 dissemination rates stayed fairly stable between 67.7% at 4 dpi, 84.6% at 8 dpi, and 76.2% at 11 dpi. These data suggest the presence of a vector competence-attenuating factor in strain B230 in Cx. tarsalis.
Utilizing the HJV whole genome sequence data, two amino acids were identified as being potentially responsible for the differences in mosquito infection. Both residues reside in the E2 glycoprotein, which is responsible for recognition, attachment, and infection of the host cell. The first residue was located at genome nucleotide position 9243 (E2 glycoprotein amino acid 282) and was found to be valine (nonpolar) in strain B230 and isoleucine (nonpolar) in all other strains studied here. The second residue was located at genome nucleotide position 8605 (E2 glycoprotein amino acid 69) and was a glycine (nonpolar) in both lineage 1 strains and glutamic acid (acidic) in all other strains studied here. Based on this data, strain 64A-1519 was chosen to discern which of the two amino acid differences was likely responsible for the attenuated phenotype of B230. Strain 64A-1519 contained glycine at nucleotide position 8605 (like B230) and isoleucine at nucleotide position 9243 (unlike B230) (Table 4).
Strain 64A-1519 was found to exhibit limited infection rates of 30.7% at 4 dpi and 31.3% at 11 dpi, similar to those of B230 (4 dpi p = 0.20, 11 dpi p = 0.62), with dissemination rates ranging between 65.2% at 4 dpi and 96.0% at 11 dpi. This data indicated that the glycine at E2 amino acid 69 found only in lineage 1 strains is likely to be responsible for the observed attenuation of infection due to a midgut infection barrier. The valine at position 9243 in B230 may have a modulating effect on dissemination since it is the only residue unique to B230; an isoleucine is present at that position in all other strains tested.
To investigate whether there was a difference in viral titers between the permissive lineage 2 and 4 strains and the less-permissive lineage 1 strains, qRT-PCR was performed on a subset of samples from the CPE testing (n = 490). For the bodies, all of the strains tested exhibited similar titer profiles with slightly lower titers at 4 dpi followed by a rise in titer on 8 and 11 dpi (Table 5). WX3-2AP reached the highest average titers, but for all strains titers ranged between 6.60 log10 and 7.26 log10 pfu equivalents/body. Titers for the heads ranged between 6.01 log10 and 6.80 log10 pfu equivalents/head for all strains tested. Interestingly, while lineage 2 and 4 strains had increasing titers at later time points, 64A-1519 exhibited the lowest mean titers in the head at day 11. Of note, all strains of HJV were able to replicate to high titers regardless of the strain phenotype.
High infection and dissemination rates may not directly translate into the ability of the mosquito to transmit the virus if a salivary gland barrier exists. To assess the potential for transmission, saliva was collected at 8 dpi from mosquitoes infected with strain AB-80-9. Of 49 mosquitoes provided an infectious bloodmeal, 39 (79.6%) became infected and 27 (69.2%) established a disseminated infection (data not shown). Twenty-two of the 27 (81.5%) with disseminated infection expectorated detectible levels of virus (Table 6), viral titers ranged between 1.68 and 5.81 log10 pfu equivalents/saliva, and the log average titer was 3.59 log10 pfu equivalents/saliva indicating that no salivary gland escape barrier exists.
Discussion
Genomic sequencing of HJV showed a high level of conservation among strains. Even between the most diverse strains identified, 99.62% amino acid identity was found. This level of identity allowed the identification of individual viral genetic elements potentially responsible for phenotypic differences that would be difficult to discern among more diverse species. While this study focused on the amino acids with a nonconservative nature, other nucleic acid differences in the nontranslated regions or differences that result in conservative amino acid changes should still be considered.
It has been assumed that HJV was never isolated in the western United States because the distribution of the presumed vector, Cs. melanura, was limited to the eastern third of the United States. In addition, numerous other mosquito species from which HJV has been isolated have a similarly restricted geographic distribution. Even if HJV could use vectors from western geographic regions, there has been a lack of knowledge regarding minimum infection rates necessary to allow transmission. Currently, the infective dose required for transmission of HJV is unknown even for the eastern mosquito vectors. Previous studies with Cs. melanura and EEEV revealed that bloodmeal doses of 4.8 log10 pfu/mL resulted in dissemination in 100% of mosquitoes by day 7 (Weaver et al. 1990). WEEV vector competence in Cx. tarsalis KNWR mosquitoes is dose responsive, with 5.8 log10 pfu/mL bloodmeals leading to 2.6-fold greater infection, 2.8-fold greater dissemination, and fivefold greater transmission rates than 3.6 log10 pfu/mL meals, and infected mosquitoes were capable of expectorating an average of 2.1 log10 pfu (Mahmood et al. 2006). While the precise volume of saliva expectorated in each saliva sample in this study could not be measured, Cx. tarsalis mosquitoes were clearly capable of developing salivary HJV titers greater than or similar to mosquitoes infected with WEEV, for which Cx. tarsalis is the primary vector. These results clearly show that Cx. tarsalis has the ability to serve as a vector for HJV.
As with all laboratory studies using colonized strains of mosquito (such as Cx. tarsalis KNWR strain, which has been in laboratory colonization since 2004), the transmission results could potentially be distinct from those of more recent field populations. However, clear differences in infection patterns were identified among different HJV strains strengthening the argument that vector competence and transmission potential of this species are not due to laboratory adaptation.
The results of this study raise interesting questions about HJV and WEEV surveillance and the possible impact of HJV transmission by Cx. tarsalis in the western United States. Cross-reactivity between HJV antibody and WEEV antigen has been widely reported in enzyme immunoassay, complement-fixation, hemagglutination-inhibition, radioimmune precipitation, and plaque-reduction neutralization tests (Jennings et al. 1966, Karabatsos 1985, Trent and Grant 1980, Calisher et al. 1988). This cross-reactivity makes these tests potentially unreliable for the serological laboratory diagnosis of WEEV and HJV. Molecular-based diagnostic tests are specific enough to differentiate the two viruses, but virus must be present in the blood of the patient at the time of diagnosis, often limiting the use of these nucleic acid detection methods. Because HJV has never been identified in the western United States and because human cases of WEEV have all but disappeared since the 1990s, laboratory testing for WEEV is not routinely conducted and the possibility of HJV causing infections in the western United States has not been explored.
The unexpected finding of Cx. tarsalis vector competence for HJV indicates that additional research is needed to further clarify the role of this mosquito in the natural transmission of HJV. While E2 amino acid 69 appears to be the single essential element for Cx. tarsalis infectivity, confirmation using infectious clone technology is needed. In addition, HJV has traditionally been an understudied alphavirus, but its relationship with the human and equine pathogen WEEV and the potential for transmission by WEEV vectors indicate the importance of further characterization of this alphavirus.
Footnotes
Acknowledgments
The authors thank Andrea Peterson for maintaining the Culex tarsalis KNWR mosquitoes used in this study. The authors also thank Dr. Sandra Quackenbush and Dr. Chaoping Chen for valuable insight and helpful discussions throughout the project. The findings and conclusions in this report are those of the author(s) and do not necessarily represent the official position of the Centers for Disease Control and Prevention.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.