
Abstract
Affiliation to four phylogroups (A, B1, B2, and D) was examined, among 190 Escherichia coli strains, collected from five, wild waterbird species, including the following: the Greylag goose—Anser anser (61) and the Canada goose—Branta canadensis (33) obtained in the Netherlands, and the Mallard—Anas platyrhynchos (38), the Mute swan—Cygnus olor (37), and the Great cormorant—Phalacrocorax carbo (21) obtained in Poland. Moreover, the prevalence of 10 virulence factors: astA, iss, iucD, irp2, papC, tsh, vat, cva/cvi, stx2f, and bfp, as well as antimicrobial susceptibility to amoxicillin, enrofloxacin, and tetracycline (minimum inhibitory concentration [MIC] using E-tests) were investigated, in the examined E. coli strains. Results demonstrated that the greatest number of E. coli strains belonged to phylogenetic groups, B1 (86 strains—45.3%) and D (49 strains—25.8%), whereas 40 (21.0%) and only 15 (7.9%) isolates were classified as being of phylogenetic groups, A and B2, respectively. Among the 10 tested virulence-associated genes, 7 genes were detected in 61 examined strains (32.1%) with highly varying frequency. Virulence profiles showed that astA, iss, and irp2 genes were detected most frequently among all examined E. coli strains, isolated from every chosen bird species. Antimicrobial susceptibility, as detected by MIC for the examined antibiotics, is variable among strains isolated from different species of birds. The aim of this study was to examine the prevalence of E. coli strains, isolated from different species of wild waterbirds and determine their potential pathogenicity to the environment, other birds, and people.
Introduction
E
Phylogenetically, E. coli are traditionally assigned to four major phylogenetic groups, namely A, B1, B2, and D (Herzer et al. 1990). Usually, extraintestinal pathogenic strains belong to groups B2 and D (Picard et al. 1999, Johnson and Stel 2000), while isolates belonging to groups A and B1 are more often strictly commensal strains from human intestinal microbiota (Clermont et al. 2000). Groups A and B1 carry few virulence-associated genes (VGs), while pathogenic groups B2 and D usually possess VGs, which enhance colonic persistence and adhesion in the UT (Clermont et al. 2000). Maynard et al. (2004) reported that the majority of human isolates belong to a virulent group (group B2 or D), while strains isolated from animal tissues belong to either a commensal group (group A or B1) or a virulent group (group B2 or D) (Maynard et al. 2004).
Only some E. coli strains are endowed with virulence factors, including adhesins, iron acquisition systems, hemagglutination, hemolysins, antibactericidal factors (e.g., colicin production), and toxins, enabling them to cause disease. Although ExPEC strains are usually considered to be host adapted, similar arrays of virulence factors may be shared by isolates from different animal species (Ron 2006, Moulin-Schouleur et al. 2007). The antimicrobial susceptibility of E. coli is also an important point in the molecular characteristic of the examined strains. Fecal E. coli is regarded as a useful indicator of the spread of acquired antibiotic-resistant genes in the community (Nys et al. 2004).
Surveillance data have shown that resistance in E. coli was consistently increasing for antimicrobial agents that have been in use the longest time, in human and veterinary medicine. The bacteria were especially resistant to fluoroquinolones (US Food and Drug Administration 2010, ECDC/EFSA/EMA 2015).
The appearance of multidrug-resistant bacteria, among animals, is accompanied by cocontamination of the environment and a reduction in the chance of successful treatment, in humans (Grobbel et al. 2007, Martinez 2009). Moreover, Shobrak and Abo-Amer (2014) reported that the presence of bacteria, with potential zoonotic importance, among migrating and nonmigrating wild birds has public health significance.
Free-living waterbirds can also act as reservoirs for coliform bacteria, such as E. coli, with large differences in pathogenicity (Kobayashi et al. 2009, Radhouani et al. 2009). For this reason, the aim of this study was to evaluate the distribution of phylogenetic groups and prevalence of virulence factors in E. coli isolates from migratory wild-bird species, as well as determine the sensitivity of these E. coli strains (by determination of minimum inhibitory concentration values [MIC]) to three antimicrobials: amoxicillin, enrofloxacin, and tetracycline, selected because of their importance to poultry production and human medicine.
The Netherlands and Poland are countries with long ornithological histories and traditions. Both countries are also regarded as precious habitats for different species of migrating free-living waterbirds, living both inside cities and on the outskirts of human habitations. It begs the following question: might the differences between waterbird species and the climate of both countries impact on the diversity of examined free-living waterbird microflora and could that suggest a potential zoonotic importance?
Materials and Methods
Collection of samples from birds in the Netherlands
In total, cloacal samples were taken from 61 Greylag geese and 33 Canada geese. The geese were caught in molt catches (Voslamber et al. 2010). The birds were prompted to walk or swim in the direction of a funnel-shaped net, ending in a corral (Persson 1994). Time periods depended on the species. Greylag geese molt mainly in June, while Canada geese from mid-June until mid-July. All birds were caught in open areas, where they feed on grass. These areas can be agricultural or natural grassland, but also large lawns, in parks. The birds were caught in different parts of the Netherlands: Greylag geese: in parks, from the city of Arnhem (52.00°N, 5.90°E) in a park close to the city of Zoetermeer (52.06°N, 4.45°E) from the Molengreend nature reserve (51.16°N, 5.90°E). The Canada geese came from a park, in the city of Haren (53.17°N, 6.63°E).
Collection of samples from birds in Poland
Cloacal swabs, from the Mallard, were obtained during the hunting season, by two hunting associations, in accordance with local hunting laws, special permission (with the consent of the Regional Directorate of Environmental Protection, Wrocław, Poland, no. WPN. 6205.67.2012.MK.1), and recognized hunting programs. The hunting season occurs from mid-August until mid-December. Cloacal swabs from Great cormorants were obtained during the annual population cull, which takes place from July until the end of November, with the same special consent of the Regional Directorate of Environmental Protection. All cloacal swabs from mallards and cormorants were collected from the lakes of the Lower Silesia region (51.2667°N, 17.3333°E; 51.3044°N, 16.5939°E). Cloacal swabs from swans were collected from the lakes of Wrocław, in Poland (51.1000°N, 17.0333°E), and were obtained by ornithologists during bird ringing, with the consent of the General Directorate of Environmental Protection, Poland (nos. 253/2012 and 259/2013). The birds were caught in July (young) and January (young and adult). All of the samples were collected between September 2011 and December 2013.
Collection of bacterial strains
A total of 190 E. coli strains were studied, in this research. All of the strains were collected from two wild waterbird species, in the Netherlands: the Greylag goose—Anser anser (61), the Canada goose—Branta canadensis (33), and from three species in Poland, including the following: the Mallard—Anas platyrhynchos (38), the Mute swan—Cygnus olor (37), and the Great cormorant—Phalacrocorax carbo (21).
Isolation of E. coli
Standard methods were used for the enrichment, isolation, identification, and biochemical confirmation of Escherichia spp. isolates. Cloacal swabs were enriched in 9 mL of buffered peptone water (Merck) at 37°C, for 24 h. Subsequently, cultures were streaked with a 1 μL loop on MacConkey agar (Biomed) and incubated overnight at 37°C. Three colonies per plate, with the characteristic morphology of E. coli, were then spread onto a nutrient agar (Merck) and incubated for 24 h at 37°C. All isolates were morphologically characterized with a Gram stain. For biochemical characterization, isolates were further confirmed with analytical, profile index API 20E strips (Bio Merieux). All isolates were stored in Microbank vials (Microbank, Pro-Lab Diagnostics) at −70°C for further analysis.
Phylogenetic and virulence genotyping
One colony of E. coli strains were incubated overnight at 37°C in 3 mL of buffered peptone water (Merck). The bacterial genomic DNA was extracted using a GenElute (Sigma-Aldrich) according to manufacturer's instructions. The DNA was quantified spectrophotometrically (BioPhotometer, Eppendorf) and stored at −20°C.
All strains were assigned to one of the four major E. coli phylogenetic groups (A, B1, B2, and D) by the use of a multiplex polymerase chain reaction (PCR) based on the method described by Clermont et al. (2000). Furthermore, each strain was examined for the presence of 10 virulence genes, using multiplex PCR assays. These were then applied to the following categories of genes, based on the method described by Ewers at al. (2004 and 2005) and Vidal et al. (2004): a heat-stable enterotoxin (astA), an increased serum survival protein (iss), a gene for iron acquisition-related factors aerobactin (iucD), and iron-repressible protein (irp2) colonization factors, such as a pyelonephritis-associated pili (papC), a temperature-sensitive hemagglutinin (tsh), and a recently described APEC, vacuolating autotransporter toxin (vat), as well as a gene from the colicin V (ColV) plasmid operon genes (cva/cvi). In addition, a verocytotoxigenic/Shiga toxin called avian-specific cytotoxin (stx2f) and a gene encoding, bundle forming protein (bfp) were examined.
The primer sequences used in amplification studies and references are summarized in Table 1.
Antimicrobial susceptibility
Antimicrobial susceptibility to amoxicillin, enrofloxacin, and tetracycline (MIC) was evaluated for each E. coli strain, using E-Tests (BioMerieux) on Mueller-Hinton agar plates (Merck) in accordance with Clinical and Laboratory Standards Institute guidelines (CLSI 2014). CLSI breakpoints were used to interpret the results. E. coli ATCC 25922 were used as control strains. Bacterial isolates were categorized as susceptible, intermediate, or resistant to antimicrobials, using interpretive criteria published by the CLSI. Values of MIC50 and MIC90, for E. coli isolates, for selected antimicrobial agents, were evaluated.
Results
Results demonstrated that the greatest number of E. coli strains belonged to phylogenetic groups B1 (86 strains—45.3%) and D (49 strains—25.8%), whereas 40 (21.0%) and only 15 (7.9%) isolates were classified to phylogenetic groups A and B2, respectively. E. coli, belonging to the group B1, had higher prevalence in the Greylag goose (57.4%), the Great cormorant (57.1%), and the Mallard (55.3%). Most of the strains isolated from the Canada goose belonged to group A (51.5%). Group D was the most common group among the E. coli isolated from the Mute swan (59.5%) (Fig. 1).
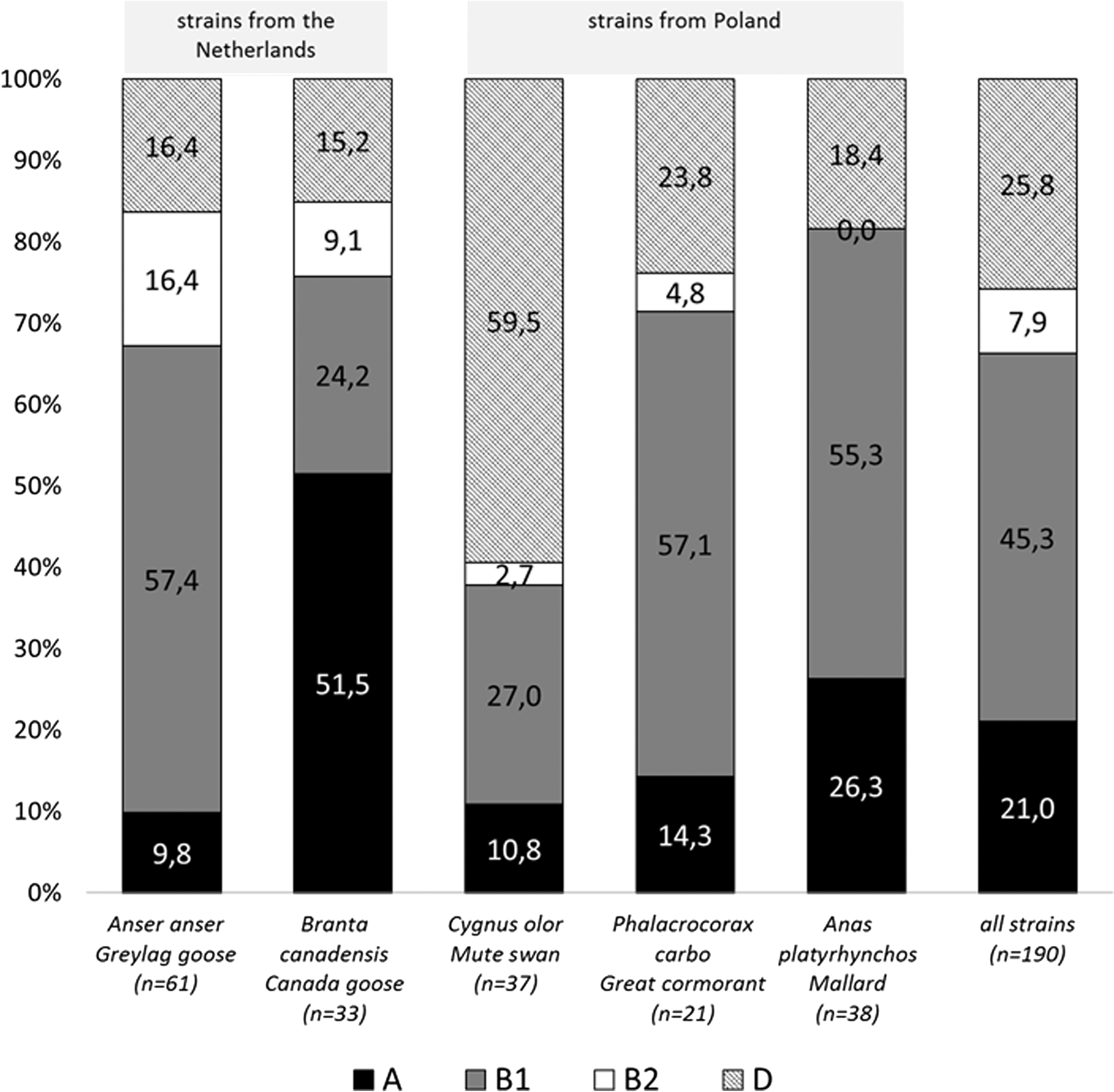
Phylogenetic groups of Escherichia coli isolated from free-living birds in the Netherlands and Poland.
E. coli strains collected from geese, in the Netherlands, mostly belonged to phylogroups B1 and A, while strains obtained from birds in Poland mostly belonged to phylogroups B1 and D (Fig. 1).
Among the 10 tested VGs, 7 genes were detected in 61 examined strains (32.1%) with highly varying frequency. Virulence profiles showed that astA, irp2, and iss genes were detected most frequently, among all examined E. coli strains isolated from every bird species. The highest percentage of the irp2 gene was found in Canada geese (15.2% of isolates) and Great cormorants (14.3%). The iss gene was recorded, most often, in isolates from Great cormorants (14.3%). The astA gene was mostly detected in the Mallard (13.2%) and Greylag goose (13.1%), as well as Canada goose (12.1%). Gene encoding P-fimbriae (papC) was detected only in isolates from Poland, while a gene encoding, vacuolating autotransporter toxin (vat) was detected only in isolates from the Netherlands. None of the 190 isolates presented the cvi/cva, stx2f, or bfp genes (Table 2). Most E. coli strains had one gene, but there were single strains, from Great cormorants, with multiple virulence genes (date not shown).
Antimicrobial susceptibility, detected by MIC, for the examined antibiotics, is variable among different species of birds.
Isolates of E. coli, obtained from Mallards, showed the highest resistance to tetracycline (15.8% of all isolates obtained from this bird species). The value of MIC90 for this group of bacteria comprised 64 μg/mL, while MIC50 comprised 6 μg/mL. By contrast, all strains isolated from the Canada geese showed susceptibility to tetracycline. The value of MIC50, for this group, was 4 μg/mL, while MIC90 was 8 μg/mL. MIC50 and MIC90 for tetracycline in Mute swans were 3 and 8 μg/mL, respectively. In strains isolated from Great cormorants, values of MIC50 and MIC90 were 6 and 16 μg/mL, but in Greylag geese, 4 and 8 μg/mL, respectively (Table 3). Resistance to amoxicillin was very wide and variable among all isolates in every bird species.
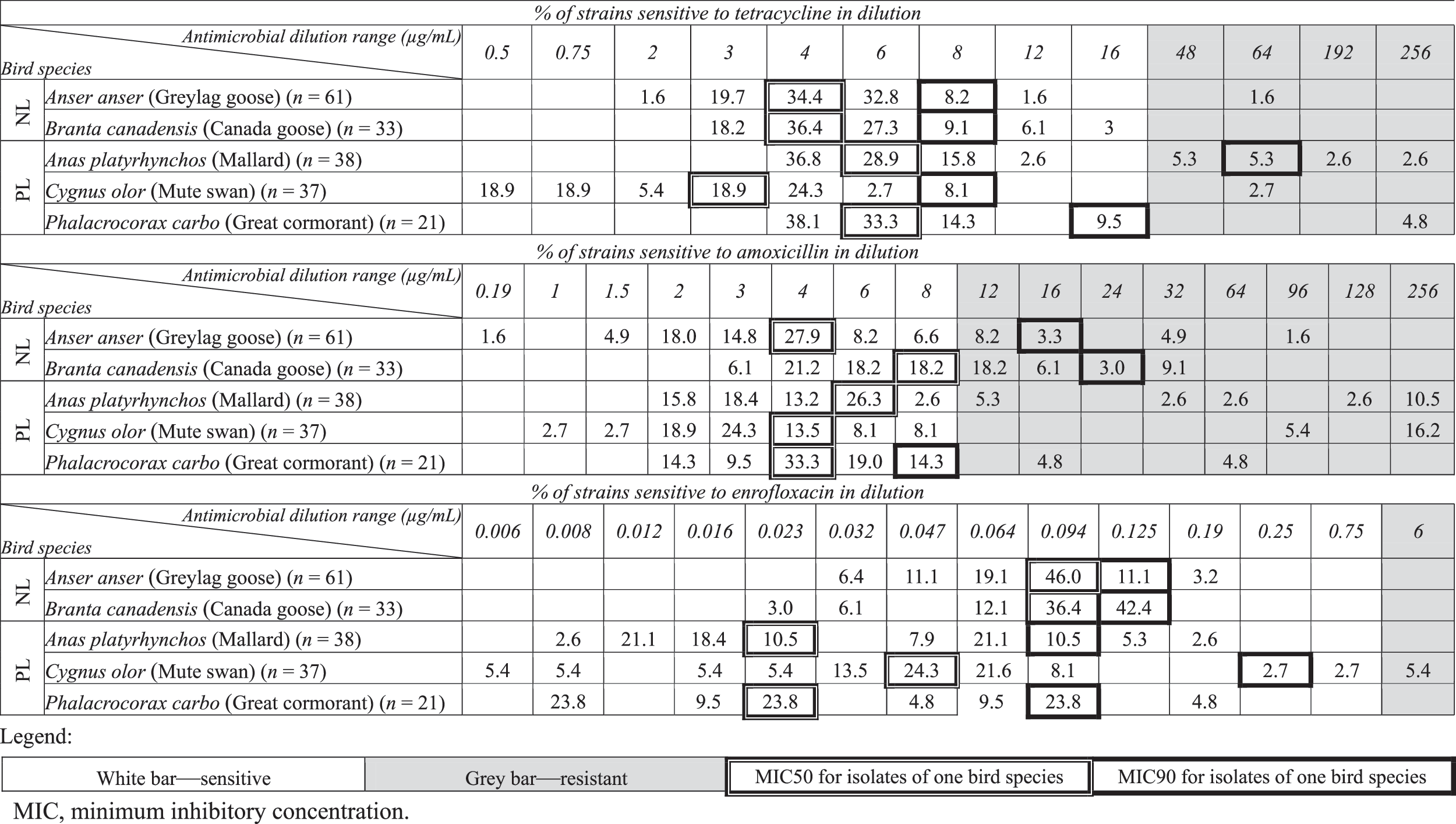
Most resistant strains were detected in the group of Canada geese (36.4% of isolates obtained from this bird species) and values of MIC50 and MIC90, for this antimicrobial, were 8 and 24 μg/mL, respectively. Moreover, among isolates obtained from Mallards (10.5%) and swans (16.2%), MIC amounted to 256 μg/mL or more, and because of that, MIC90 could not be identified exactly, using our method. Values for MIC50 for amoxicillin amounted to as follows: in Mallard, 6 μg/mL and in Mute swans, 4 μg/mL, respectively. Isolates from Great cormorants showed the value of MIC50 as 4 μg/mL and MIC90 as 8 μg/mL, while isolates collected from Greylag geese were 4 and 16 μg/mL, respectively (Table 3). Resistance to enrofloxacin was found only in isolates obtained from Mute swans. The values of MIC50 and MIC90 for this antimicrobial amounted to as follows : in Mallard and Great cormorants, 0.023 and 0.094 μg/mL (in both bird species), respectively; in Mute swans, 0.047 and 0.25 μg/mL, but in Canada and Greylag geese, the values were 0.094 and 0.125 μg/mL.
Discussion
E. coli can be easily disseminated, into different ecosystems, through faces or the fecal pollution of water. This is one of the reasons why waterbirds have been intensively used as an indicator for the fecal pollution of the environment, particularly water (Martinez 2009, Wellington et al. 2013). The molecular characteristics of bacteria obtained from migratory wild waterbirds, spending their lives on public ponds, might have an important significance for human health and livestock (Edberg et al. 2000, Machado et al. 2008).
Results obtained in this study showed that the biggest number of examined avian E. coli strains belonged to phylogenetic group B1, which may suggest that, in general, wild waterfowl transmit mostly nonpathogenic bacteria. However, there are some notable exceptions among examined bird species. Most of the E. coli strains isolated from Mute swans belonged to group D, which suggests that this bird species could be, in particular, a potential reservoir for E. coli, from this virulent phylogenetic group. The other pathogenic phylogenetic group, B2, was detected at a lower percentage, among strains obtained from all examined bird species, except Mallard.
Comparing the isolates of E. coli obtained from Mute swans, Great cormorants, and the Mallard (in Poland) with strains from Greylag geese and Canada geese (in the Netherlands), it should be emphasized that phylogroup D was dominant among strains from species of birds examined in Poland, while group B2 was more often detected in strains isolated from bird species analyzed in the Netherlands. The presence of both phylogroups D and B2 as pathogenic may indicate the presence of pathogenic E. coli in these environments. Such results might be interesting, compared to results reported by Ahmed et al. (2011) who examined E. coli isolated from rainwater tanks. Rainwater also fills up ponds, where wild waterbirds spend their time. These authors detected that 22.5% of isolates belonged to phylogenetic group B2, while 29.5% belonged to group D. Other authors have reported that distribution between phylogenetic groups B2 and D in bird isolates was 18.8% and 35.9%, respectively (Escobar-Paramo et al. 2006). These authors reported that one of the main forces which shapes the genetic structure of E. coli populations, among hosts, is domestication. In accordance to a report by Tenaillon et al. (2010), domesticated animals have a decreased proportion of B2 strains, compared to wild animals, and an increased proportion of A strains. This important fact, in turn, is also visible in results obtained in our study and might explain the high percentage of E. coli strains, from phylogroup A, among isolates from Canada geese, because the Canada geese, in the Netherlands, originate from birds that have been in captivity for several generations.
In previous studies, for example, in Portugal, the prevalence of E. coli, of phylogroups A and B1, was observed in wild waterbirds, such as seagulls (Radhouani et al. 2009), while research conducted by Simoes et al. (2010) showed that 37% of all extended-spectrum Beta-lactamases producing E. coli, isolated from seagulls, belonged to the B2 or D phylogroups. Results obtained by Radhouani et al. (2009) in wild birds and by Machado et al. (2008) in chickens and swine showed that E. coli isolates from phylogenetic groups A and D were predominant. As reported by Gunther et al. (2011) in Germany, phylogenetic typing of E. coli isolates, from wildlife, revealed that a large proportion of multiresistant isolates belonged to group B2 (46%). Results of more recent studies indicate that a high percentage of migrating bird samples contained E. coli and the same samples showed the presence of selected virulence factors (Shobrak and Abo-Amer 2014). In this study, a low percentage of isolates (only 32%) showed the presence, if any, of examined virulence factors. This is lower than results obtained by Camarda et al. (2006) in other wild waterbirds, like gulls, where 62.5% of examined strains were positive for one or two genes.
Virulent genes examined by us, such as astA, iss, iucD, irp2, tsh, vat, and cvi/cva, as well as papC, are characteristic and pathogenic to poultry and are commonly detected among chickens with colibacillosis (Maynard et al. 2004).
One of the most important factors, which defines the APEC pathotype, is the ColV plasmid. Besides bacteriocin ColV, this plasmid encodes other traits, such as, iss, the aerobactin operon, and tsh, which are responsible for virulence (Johnson et al. 2006). Previous investigations have indicated that the prevalence of a ColV plasmid, among isolates from chicken with colibacillosis, is estimated to be from 35.5% to 62.7% (Ewers et al. 2004).
None of the isolates examined, in this study, had the ColV plasmid (gene cvi/cva). However, the investigated strains showed the variable prevalence of the iss, tsh, and iucD genes. Moreover, the other siderophore gene encoding yersiniobaktin (irp2) showed the highest prevalence, among all investigated genes. An enteroaggregative heat-stable toxin gene (astA) was detected among strains from every bird species, while a vacuolating autotransporter toxin (vat) was found only in isolates collected in the Netherlands.
In the group of gene-encoding factors characteristic of enterotoxigenic E. coli, which are able to affect people, none of the stx2 and bfp genes were found. Also, a low amount of isolates (E. coli strains isolated only in Poland) showed the presence of the papC gene, characteristic for uropathogenic E. coli. Compared to other results available in literature, obtained from wild Starlings or Common buzzards, such a low prevalence of the gene papC is similar (Gaukler et al. 2009, Radhouani et al. 2012). Kobayashi et al. (2009), who examined the prevalence of the gene stx among isolates from many different bird species, did not detect this gene in any waterbird. However, the situation is a little different, when compared to poultry. It has been shown that P fimbriae (expressed by the papC gene) are expressed in vivo, in the air sacs, lungs, kidneys, blood, and pericardial fluid (Kariyawasam et al. 2006).
Wild birds do not naturally come into contact with antimicrobials; however, they may be infected with antibiotic-resistant bacteria by the acquisition of resistant bacteria from human sources, agricultural facilities, and associated contaminated environments (Dolejska et al. 2007, Alroy and Ellis 2011). Antimicrobial-resistant E. coli isolates, originating from wild bird species, were reported for the first time, by Sato et al. (1978) and Kanai et al. (1981). Nowadays, the number of studies describing the occurrence of antimicrobial resistant E. coli has increased.
Results obtained in this study show that E. coli, isolated from bird species in Poland, were more resistant to three examined antimicrobials than strains obtained from geese in the Netherlands. One suspected reason for this might be the difference between the location and environment, in both countries, where the samples were collected. All strains in Poland came from birds, which spend their lives on public ponds, where many people and whole families spend their free time, often feeding the birds. Samples from the Netherlands were collected from the populations of geese, which live both in and outside cities, some of them having limited contact with people. In most studies (Middleton and Ambrose 2005, Dolejska et al. 2007), the wild bird populations have relatively frequent interactions with habitat influenced by human activities, although Sjolund et al. (2008) demonstrated antimicrobial-resistant E. coli originating from Arctic birds, in a region of the world where human influence on the ecology of antimicrobial resistance was expected to be minimal.
Cole et al. (2005) detected high rates of resistance (64%) to tetracycline in E. coli isolated from migratory Canadian geese. The presence of E. coli strains, resistant to tetracycline, was also found by Camarda et al. (2006) in gulls from Italy. The percentage of resistance among examined strains was 22.9%. Compared to our study, this percentage of resistance to tetracycline was significantly higher. Results obtained by Middleton and Ambrose (2005) in Canada geese, from the USA, suggested that all 63 E. coli strains obtained from these birds were resistant to β-lactam antibiotics. In our study, 36.4% of isolates from Canada geese were resistant to amoxicillin. Isolates from other wild waterbirds, like gulls, were resistant in 27.1% of isolates (Camarda et al. 2006).
Moreover, isolates from double-crested cormorants, collected by Dobbin et al. (2005) in Canada, showed full susceptibility to enrofloxacin. This is similar to the results obtained in our study where E. coli from the examined Great cormorants also showed full susceptibility to enrofloxacin. Resistance to enrofloxacin was also detected only in 5.4% of strains isolated from Mute swans. Isolates from gulls, in Italy, were resistant to a similar percentage (Camarda et al. 2006).
Most of the clinically important antimicrobial agents, such as macrolides, sulfonamides, fluorochinolones, and tetracyclines, have already been detected in soils, surface water, and groundwater, and it has been proven that tetracyclines and fluorochinolones are able to persist in the environment for months and even years (Hirsch et al. 1999, Boxall et al. 2006, Milic et al. 2013, Monteiro and Boxall 2010). Such a situation might be important for increasing antimicrobial resistance among bacteria in wildlife
Conclusion
In conclusion, these studies suggest that wild waterbirds, in the Netherlands and Poland, could be vectors for potentially virulent E. coli strains, including antimicrobial-resistant variants. These strains may contribute to the pollution of superficial waters, as well as natural grasslands and parks, often attended by people.
Footnotes
Acknowledgments
This research project was financed by the National Centre for Research and Development, project number, no. 12 0126 10 (part led in Poland), and by the Dutch Faunafonds and the Dutch province of Limburg.
Publication was supported by the Wroclaw Centre of Biotechnology, the Leading National Research Centre Programme (KNOW) for years 2014–18.
The authors are grateful to Jan H. Beekman, Jorina Boer, Loes van den Bremer, Hennie van den Brink ( † ), Marian Crombach, Annita Logotheti, Gerard Muskens, Phillip van de Ven, Małgorzata Pietkiewicz, and people from the “Odra” Bird Ringing Group, Mieczysław Łyskwa, Piotr Zajiczek, and Stanisław Rusiecki for their help with collecting the samples.
Author Disclosure Statement
No competing financial interests exist.