
Abstract
Objective:
To understand the distribution and infection status of hantavirus in Myomorph rodents on Bolshoy Ussuriysky Island (Heixiazi Island) at the Sino-Russian border, and to provide data for the safe development and utilization of Bolshoy Ussuriysky Island.
Methods:
In 2013 and 2014, Myomorph rodents were trapped on Bolshoy Ussuriysky Island. Total RNA was extracted from rodent tissue, and it was screened for hantavirus RNA by using reverse transcription-polymerase chain reaction. Univariate and multivariate nonconditional logistic regression analysis was used to analyze the RNA prevalence rates in eight species of rodents, in relation to species, sex, age, habitat, and season. In addition, PCR amplicons were sequenced and phylogenetic analysis was performed by using Mega 5.1 software.
Results:
Six hundred forty-four rodents belonging to three orders, five families, and eight genera were trapped. Fifty-two rodents were infected with hantavirus, and the rate of RNA detection was 8.07%. The infection rates of rodents in different habitats (χ2 = 14.853, p < 0.05) and different seasons (χ2 = 16.990, p < 0.05) showed significant differences. A logistic regression analysis showed that habitat and trapping season were risk factors of hantavirus infection (p < 0.05). Phylogenetic analysis showed that the gene sequences of positive samples were Hantaan virus and Khabarovsk virus.
Conclusion:
There are two types of hantaviruses, such as HTNV (in Apodemus agrarius, Clethrionomys rutilus, Microtus fortis, Rattus norvegicus) and KHAV (in C. rutilus), among the rodents on Bolshoy Ussuriysky Island, and season and habitat are risk factors of hantavirus infection.
Introduction
H
In the 1930s, HFRS was discovered for the first time in northeast China and far-east Russia, and the virus that caused HFRS was already widespread in these areas at that time. Previous studies (Luo and Chen 2003, Kariwa et al. 2007, Zhang et al. 2010) showed that hantavirus was the causative agent of HFRS and that hantavirus infections were serious in China. The hantavirus incidence was relatively high in areas with poor living environments and high rodent densities (Yao 2007) throughout the year, especially in summer and fall. Currently, at least seven sub-species of hantavirus have been discovered in rodents in Chinese territory adjacent to far-east Russia, which demonstrates the diversity of hantavirus in this area. In China, 73 species of vertebrates have been found to be infected by hantavirus. According to previous studies, at least nine genotypes of hantavirus cause HFRS in China, and most HFRS cases are caused by HTNV and SEOV, especially SEOV (Luo and Chen 2003).
Bolshoy Ussuriysky Island (Fig. 1) is a critical and strategic island for the navigation of the Amur River and the Ussuri River, and it is adjacent to Khabarovsk, the largest city in eastern Russia, which is located across the Amur River. The island is almost undeveloped, with habitats of shrubs, grasslands, and woods. Until now, no systematic investigation of vector-borne pathogen infection status has been carried out in the island (Liu et al. 2013). In the future, more and more people will land on the island for traveling or port trade, which makes a significantly increased infection risk of vector-borne pathogens. Therefore, the surveillance of the infection status of vector-borne pathogens on the island needs to be studied.
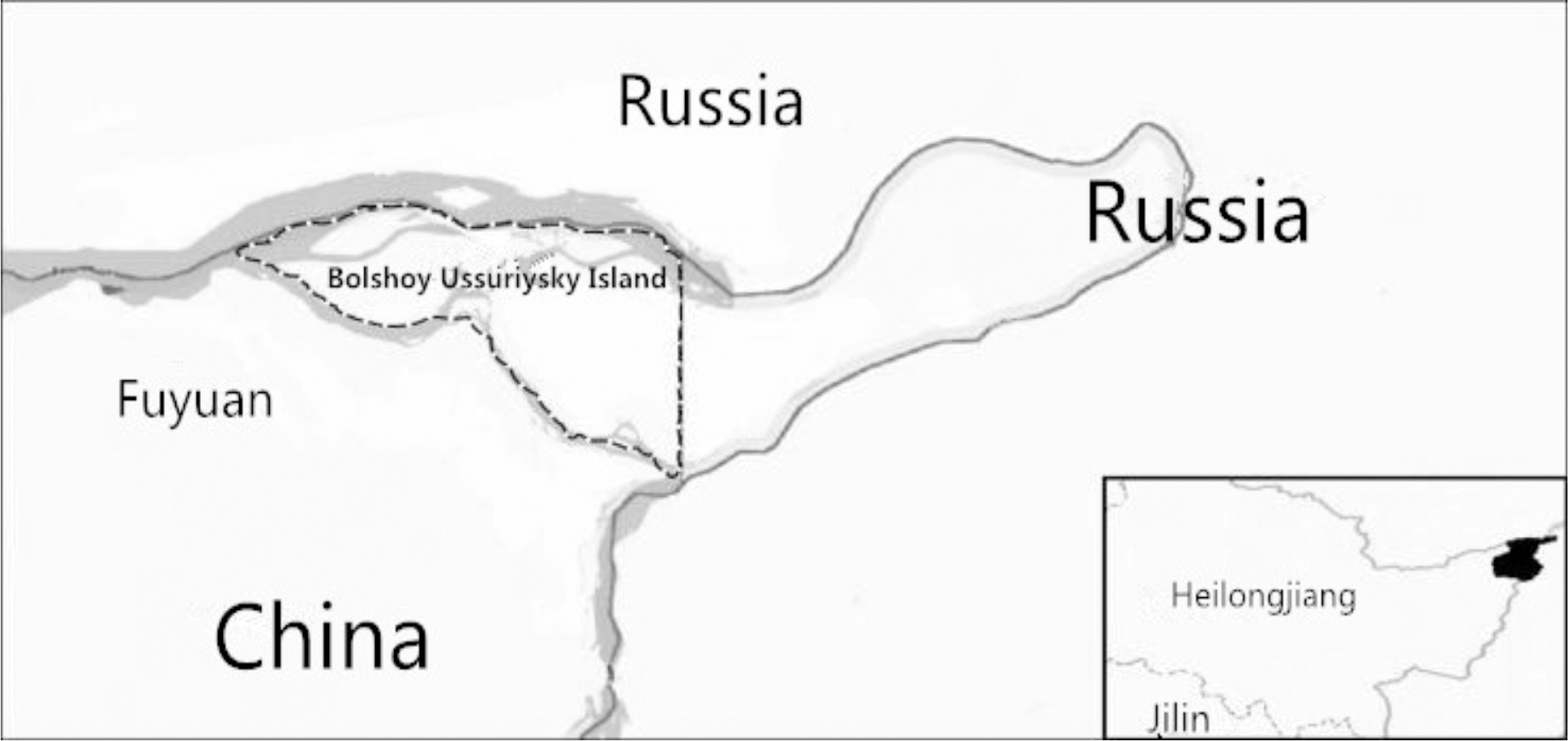
The map of Bolshoy Ussuriysky Island between China and Russia.
In this study, we examined the presence of hantavirus in rodents of Bolshoy Ussuriysky Island, which will be helpful to know and identify the factors that influence hantavirus infections in rodent populations.
Materials and Methods
Ethics statement
The handling of all rodents was conducted in compliance with the Animal Welfare Provision of the Chinese Academy of Inspection and Quarantine (CAIQ). All animal experiments were performed by following the guidelines of CAIQ.
Study site
Bolshoy Ussuriysky Island (Heixiazi Island), located in Fuyuan County of Heilongjiang Province, is the third territory to be returned to China (after the return of Hong Kong and Macao) (Fig. 1). It is 174 km2, and it ranges from 48°17′ to 48°27′N and from 134°24′ to 135°05′E.
Trapping of rodents
Rodents were trapped monthly from spring to autumn in 2013 and 2014. In 2013, rodents were trapped in April–July and in October 2013, but not in August and September because of floods. In 2014, rodents were trapped in April–October. All rodents were captured in different habitats of shrubs, grasslands, and woods. Rodents were trapped in the trap-at-night method, baited with peanut, as in Jiang et al. (2008). The rodent traps were distributed every 5 meters in the morning and were collected at night. Generally, each rodent was placed in an individual cloth bag in the field and then transported to the laboratory. After identification of their species and sex and developmental stage, the rodents were dissected. Lung, liver, and kidney tissues were extracted, stored immediately in liquid nitrogen, and transported to the laboratory for further processing.
Detection of hantavirus
The tissues of the rodents were homogenized via a tissue grinder (Micro Smash MS-100 tissuelyser; TOMY, Japan), and RNA was extracted from 140 μL of the supernatant according to the instructions of the QIAamp Viral RNA Mini Kit (Qiagen, Hilden, Germany). Total RNA was reverse transcribed into cDNA according to the instructions of the A3500 Reverse Transcription System (Promega, Beijing, China). Then, we used nested PCRs to detect the highly conserved L fragment of hantavirus. Each PCR reaction was 50 μL, which included 1 μL of DNA template, 0.5 μM MgCl2, 0.2 μM dNTPs, 1 μM of each primer (Table 1), 1 U of rTaq polymerase (TaKaRa, Dalian, China), and double-distilled H2O. Distilled H2O was used as the negative control (Table 1). The PCRs were conducted by the PCR machine (Applied Biosystems, Carlsbad, CA) (Klempa et al. 2006). PCR products were run on an agarose gel; the product bands were purified by using the Omega Gel Extraction Kit (BioTek Instruments, Inc., Winooski, VT), and they were sent to Sangon Biotech Co., Ltd. (Shanghai, China) for sequencing. Sequences were identified by using a Basic Local Alignment Search Tool (BLAST) search of GenBank. We first implemented homologic analysis to sequences using Clustal X (version 2.1) (
Statistical analysis
SPSS Statistics for Windows, Version 19.0 (IBM Corp., Armonk, NY) was used for data analysis. A univariate analysis chi-squared test was used to analyze the infection rates of rodents of distinct sexes, habitats, and trapping seasons. Fisher's exact test was utilized to analyze the hantavirus prevalence of different rodents. One-way analysis of variance was used to analyze differences between the weights of rodents in the infected and noninfected groups. A multivariable, nonconditional logistic regression model was employed to analyze factors that may influence the hantavirus prevalence. Two-tailed p < 0.05 was the criterion used to determine a statistically significant difference.
Results
Rodents' constitution
In 2013 and 2014, we trapped a total of 644 rodents that belonged to three families and six genera of Rodentia, one family and one genus of Insectivora, and one family and one genus of Carnivora. Apodemus agrarius was the dominant species on Bolshoy Ussuriysky Island, accounting for 42.7% (275/644) of all trapped rodents, followed by Clethrionomys rutilus (27.64%, 178/644), and Microtus fortis (21.12%, 136/644) (Table 2).
Hantavirus infections
Fifty-two out of the 644 trapped rodents were infected by hantavirus, which was an 8.07% positive prevalence. The RNA-positive rodents were from five different species of Rodentia. The infection rates of hantavirus among different species showed no significant differences (χ2 = 2.861, p = 0.776) (Table 3). Among the 644 trapped rodents, the difference between the infection rate of males (44/476, 9.24%) and that of females (8/168, 4.76%) was not statistically significant (χ2 = 3.355, p = 0.067). The mean weight of the rodents in the viral RNA-positive group (34.16 ± 20.80 g) was not significantly different from that in the viral RNA-negative group (32.27 ± 26.03 g) (F = 0.231, p = 0.631). However, viral RNA-positive rodents trapped in different habitats (shrub: 31/237 = 13.08%; grassland: 19/298 = 6.38%; woods: 2/109 = 1.83%) showed significant differences (χ2 = 14.853, p = 0.001). The infection rates of rodents trapped in different seasons (spring: April and May, summer: June–August, and fall: September and October) also showed significant differences (χ2 = 16.990, p = 0. 000) (Table 4).
—, no detected infection.
We used species, sex, weight, trapping season, and habitat as the covariates, where the upper limit of weight (P75) was used as the criterion for grouping. Unordered, multi-categorical variables (species, trapping season, and habitat) were utilized as the dummy variables, and whether the rodent was viral RNA positive or negative was the dependent variable for a nonconditional logistic regression analysis (the inclusion standard was 0.05, and the exclusion standard was 0.1). A. agrarius, shrub, and fall were used as the indicator indices, and the analysis showed that although trapping season, habitat, and sex were used in the logistic regression model (p < 0.1) (Table 5), only trapping season and habitat were risk factors of hantavirus prevalence (p < 0.05); whereas species, sex, and weight did not influence the risk of hantavirus prevalence (p > 0.05). According to the absolute value of the standard regression coefficient, β, trapping season, and habitat were the most and second most important factors, respectively, which influenced the risk of hantavirus prevalence.
Means p < 0.05.
CI, confidence interval; OR, odds ratio.
Phylogenetic analysis of hantavirus sequences
In this study, 52 positive results of 360 bp of the L gene fragment were employed to compare by MEGA5.0 software; 100% homologies were deleted, and 20 representative gene sequences were chosen to analyze by a phylogenetic analysis using neighbor-joining statistical methods by Clustal X2.1 software (Table 6). The results showed that the homologies ranged from 99.29% to 100% (Fig. 2). Different nucleic acid sequences of the L fragment of hantavirus are shown in Figure 3. MEGA 5.0 was employed to build a neighbor-joining polygenetic tree, and the results showed that the gene sequence of the L fragment of hantavirus was in the same clade with the known Fuyuan sequence KHAV-217(KJ857321), HTNV-26(KJ857334), and the Yakeshi sequence KHAV-31(KJ857314) (Fig. 4).

Identities of the 20 sequences of Hantavirus isolated on Bolshoy Ussuriysky Island in the study.
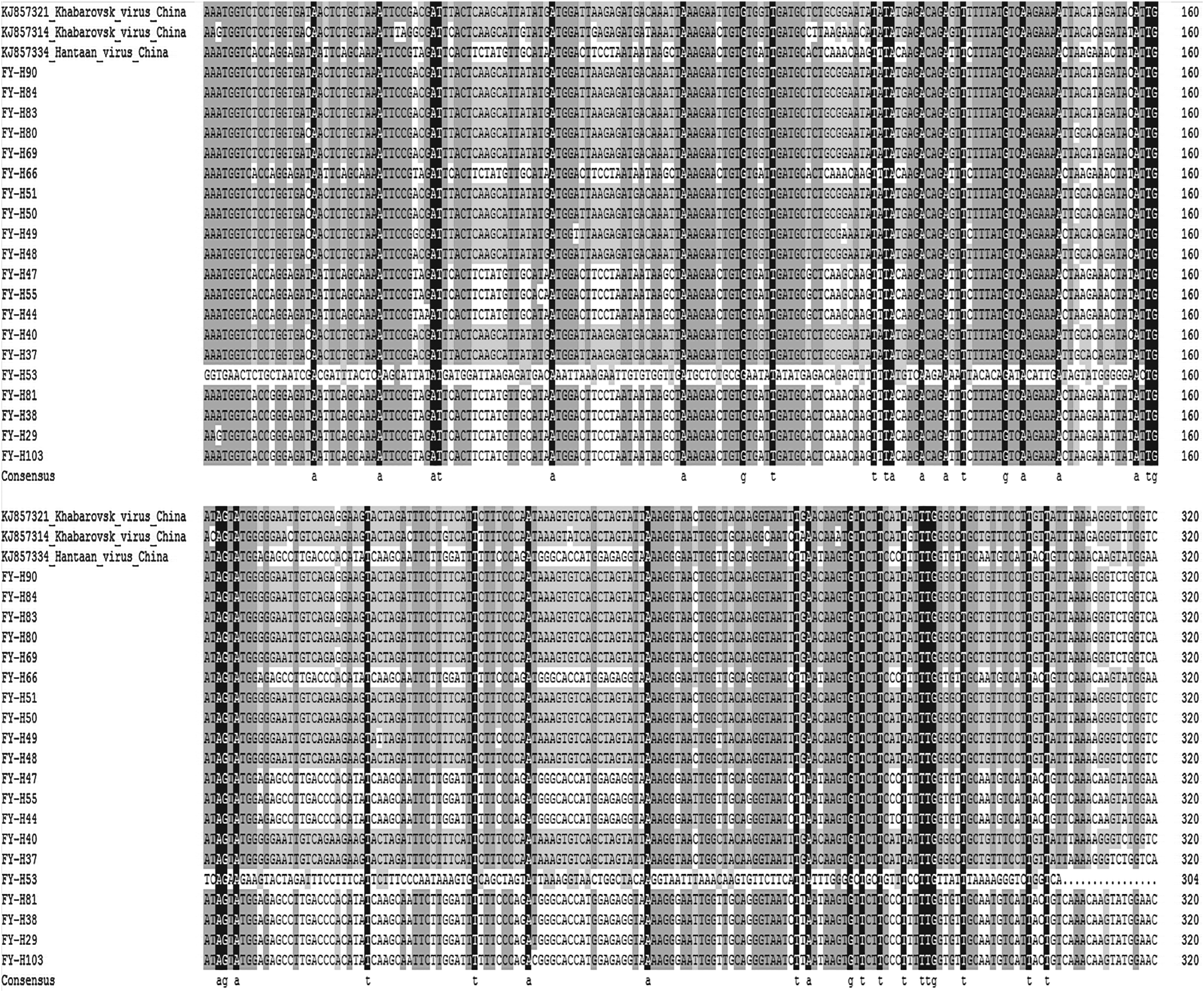
Variations of nucleotide sequences of the isolated strains of Hantavirus on Bolshoy Ussuriysky Island compared with Hantaan virus (HTNV) and Khabarovsk virus (KHAV) sequences. Figure 3 can be viewed in greater detail online at
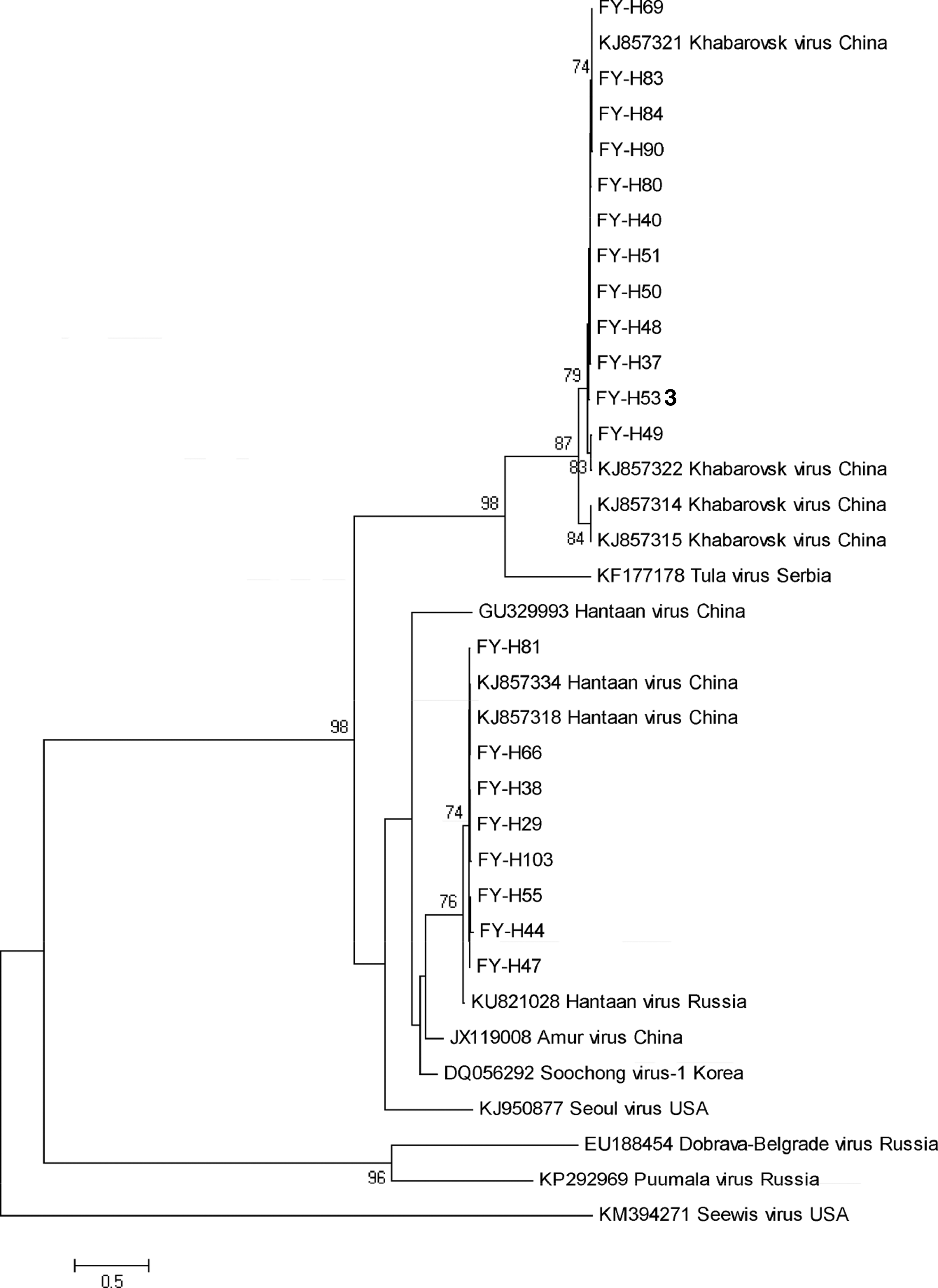
NJ phylogenetic tree based on the entire coding sequences of the Hantavirus L fragment. The accession numbers included in the tree were FY-H29(KT223834), FY-H37(KT213428), FY-H38(KT223835), FY-H40(KT213429), FY-H44(KT213430), FY-H47(KT213432), FY-H48(KT213433), FY-H49(KT213434), FY-H50(KT213435), FY-H51(KT213436), FY-H53(KT213437), FY-H55(KT213431), FY-H66(KT213421), FY-H69(KT213422), FY-H80(KT213424), FY-H81(KT223836), FY-H83(KT213425), FY-H84(KT213426), FY-H90(KT213427), and FY-H103(KT213438).
Discussion
The ecological environment on Bolshoy Ussuriysky Island is not diverse, but there are different types of rodent habitats (shurbs, grasslands, and woods). Based on the results obtained from the trapped rodents, A. agrarius is the dominant species on Bolshoy Ussuriysky Island, followed by C. rutilus, which is consistent with a previous survey of rodents along the Sino-Russian border (Yang et al. 2013). However, the results from 2013 and 2014 are not consistent (we did not collect samples in August and September of 2013 because of the flood, which led to incomplete data collection in 2013). The results showed that C. rutilus was the dominant species in 2013 and that A. agrarius was dominant in 2014, which is inconsistent with results obtained in 2011, when Wang et al. (2014) showed that the dominant species from April to October was C. rutilus, followed by A. agrarius (Liu et al. 2013, Wang et al. 2013). The distribution of rodents on Bolshoy Ussuriysky Island is likely in response to environmental changes, and it may be influenced by the flood on Bolshoy Ussuriysky Island. Therefore, the distribution of rodents on this island requires further study. Nevertheless, the results of this study are roughly consistent with the results obtained in 2011, which revealed that A. agrarius and C. rutilus are the dominant species on Bolshoy Ussuriysky Island.
In this study, an analysis using the five factors associated with hantavirus RNA prevalence (species, trapping season, habitat, sex, and weight) did not demonstrate significant differences in hantavirus RNA prevalence among different rodent species, which is possibly due to the small sample size of hantavirus-positive rodents. Therefore, the results need to be confirmed by a study with a larger sample size. However, hantavirus RNA prevalence is significantly associated with trapping season: Summer is the season of highest incidence (12.96% report incidence value here). One reason is likely due to the high temperature, which is suitable for the growth, diffusion, and direct or indirect hantavirus RNA prevalence, and it is the breeding season during which the aggression of rodents is increased. The other reason is abundant rainfall, which is suitable for increasing rodent densities because of having more vegetation, food, and water in this season (Lokugamage et al. 2004). Viral RNA prevalence was higher in 2014 than in 2013, which is likely due to the flood on Bolshoy Ussuriysky Island in 2013 that greatly changed the distribution and activity of the rodents on the island. First, although areas of the habitats may have been greatly reduced by the flood, rodents may have migrated and the rodent density in relatively high-elevation areas may have increased. Second, the constitution and quantity of the rodent population may have drastically changed. The percentage of healthy and agile adult and sub-adult rodents is likely to have increased. Third, the percentage of rodents carrying the pathogen probably increased for several factors, including the mix of rodents from distinct habitats, the mix of domestic and wild rodents, and enhanced aggression due to the lack of food, which may lead to increased cross-infections of pathogens. All of these changes may increase the risk of outbreaks of rodent-associated diseases (Glass et al. 1988, Cai et al. 2014). Further long-term studies may clarify these relationships.
Hantavirus prevalence in different habitats was significantly different: The infection rate in rodents from shrub ecosystems is significantly higher than that in grasslands and wood ecosystems. During the flood, rodents were concentrated in relatively high-elevation areas, that is, shrublands. The infection rates of different sexes are not significantly different, but this may be because the number of male rodents collected was much larger than that of females.
A multivariate logistic regression analysis further indicates that trapping season and habitat are the main factors that influence hantavirus prevalence, and that trapping season is relatively more influential (Kariwa et al. 2007). Although the regression analysis does not show that the species of rats were associated with hantavirus RNA prevalence, Apodemus and Rattus are the main hosts of hantavirus. Apodemus is the main host of HTNV, and Rattus is the main host of SEOV according to a previous report (Wang et al. 2000). However, in this study, we found that Apodemus peninsulae has a higher hantavirus RNA prevalence than the others. A previous study reported that A. peninsulae is the main host of AMRV (Zhang et al. 2007). This may be because of the original habitat of Bolshoy Ussuriysky Island, and/or the relatively smaller sample size of A. peninsulae, and/or the flood on Bolshoy Ussuriysky Island in 2013. The flood resulted in a decreased absolute number of rodents; Rattus norvegicus and A. agrarius can float and swim for 60–72 h in 35°C water, which allows them to migrate to high-elevation areas, thus possibly leading to increased local rodent density (Liu et al. 1992). A previous study has found spillover of SEOV in M. fortis for the first time within the Mudanjiang area in Heilongjiang Province, China (Cheng et al. 2015). Therefore, it is not surprising to detect high infection rates among A. peninsulae; although it is unclear whether this species is a reservoir host or an incidental host, this finding needs to be verified with further studies.
The sequence results were identified as two types of Hantavirus: HTNV and KHAV. KHAV was found in M. fortis, which is consistent with other reports. KHAV has been detected in M. fortis in Khabarovsk (Horling et al. 1996) and Microtus maximowiczii in Yakeshi (Zou et al. 2008a and 2008b), and it was subsequently found on Bolshoy Ussuriysky Island in 2011 as well (Wang et al. 2014). Therefore, the results of this study further demonstrate that there is a new KHAV virus lineage on Bolshoy Ussuriysky Island (include nucleotide homologies to support this claim), and, furthermore, that M. fortis and M. maximowiczii are likely reservoirs of KHAV on Bolshoy Ussuriysky Island. This may be related to the fact that Bolshoy Ussuriysky Island is adjacent to Khabarovsk, Russia, and that there was a certain degree of migration of KHAV in far-east Asia. According to previous studies, KHAV is remarkably different from HTNV and SEOV in terms of its nucleotide and amino acid sequences, antigenicity, and secondary structure (the amino acid sequences of the nucleoprotein (NP) are the same, but the amino acid sequences of the glycoprotein (GP) are significantly different). Thus, theoretically, the current HFRS vaccine will not effectively prevent KHAV infections. It is necessary to strengthen the monitoring of KHAV on Bolshoy Ussuriysky Island (Plyusnin 2002, Cong et al. 2012).
Conclusions
This study revealed high infection rates of hantavirus in rodents on Bolshoy Ussuriysky Island, and hantavirus was identified as types of HTNV and KHAV; season and habitat are risk factors of hantavirus RNA prevalence in rodents. Therefore, it is necessary to constantly monitor the infection rate of hantavirus in rodents on Bolshoy Ussuriysky Island.
Footnotes
Acknowledgments
This study was supported by the Project of the Public Welfare Program (201310072 and 201410020), National Project of International Cooperation in Scienceand Technology (2012DFA30540), and CAIQ Research Project (2016JK031). The authors acknowledge the Chinese Academy of Inspection and Quarantine for the use of their laboratories. They thank all of the health facility staff at the project sites for the support they offered during the sample collection.
Author Disclosure Statement
No competing financial interests exist.