
Abstract
Vector-borne protists of the genus Hepatozoon belong to the apicomplexan suborder Adeleorina. The taxonomy of Hepatozoon is unsettled and different phylogenetic clades probably represent evolutionary units deserving the status of separate genera. Throughout our review, we focus on the monophyletic assemblage of Hepatozoon spp. from carnivores, classified as Hepatozoon sensu stricto that includes important pathogens of domestic and free-ranging canine and feline hosts. We provide an overview of diagnostic methods and approaches from classical detection in biological materials, through serological tests to nucleic acid amplification tests (NAATs). Critical review of used primers for the 18S rDNA is provided, together with information on individual primer pairs. Extension of used NAATs target to cover also mitochondrial genes is suggested as a key step in understanding the diversity and molecular epidemiology of Hepatozoon infections in mammals.
Introduction
T
The life cycle of Hepatozoon sensu lato is heteroxenous, and a range of invertebrates (argasid and ixodid ticks, mosquitoes, fleas, triatomines, tsetse flies, and biting lice) serve as vectors for individual species. The vertebrate host is usually infected by ingestion of a hematophagous arthropod, although infection can be acquired also by intrauterine transmission or by predation. Infection through ingestion of an infected invertebrate vector is unlikely for many Hepatozoon s.l. from reptiles, where development can be alternatively completed through a three-host life cycle along an arthropod—paratenic host—reptile axis (Landau 1973; Sloboda et al. 2007).
In cases of infection via ingesting an infected invertebrate host, the sporozoites released from sporocysts within oocysts enter the circulation and migrate to various organs (liver, bone marrow, spleen, kidneys, lungs, intestines, and lymph nodes) where they undergo merogony. When merozoites are released from mature meronts into the bloodstream, they enter the blood cells and transform into gamonts, which are later ingested with the blood meal by a competent vector. Subsequent development includes the production of microgametes and macrogametes (in different parts of vector organs) that fuse and develop to polysporocystic oocysts (Smith 1996, Kim et al. 1998, O'Dwyer 2011).
The taxonomy of Hepatozoon s.l. is unsettled and different phylogenetic clades probably represent evolutionary units deserving the status of separate genera. Throughout our review, we focus on the monophyletic assemblage of Hepatozoon spp. from carnivores, classified as Hepatozoon sensu stricto by Karadjian et al. (2015).
Species of Importance
Hepatozoon spp. has been described in a range of mammalian hosts, mainly carnivores and rodents. While the infections in most mammalian hosts are typically subclinical, species that infect canid and felid carnivores (Hepatozoon americanus, Hepatozoon canis, Hepatozoon felis) are of veterinary importance. Currently, the ubiquitous presence of H. canis and closely related species in European foxes is attracting considerable attention (Najm et al. 2014, Hodžić et al. 2015) inasmuch as the main tick vector of this species, Rhipicephalus sanguineus, is not endemic in some areas where infection is abundant in local foxes.
H. canis infections in domestic dogs can vary from subclinical to severe, with animals manifesting extreme lethargy, cachexia, and anemia (Baneth and Weigler 1997). Infection with this species or its variants in wildlife canids has not been clearly associated with clinical disease to date. In contrast, Hepatozoon americanum infection of domestic dogs is associated with severe disease characterized by muscular pain induced by myositis, severe lameness, and subsequent muscle atrophy. Infections with H. felis primarily target striated muscle in domestic cats, but these also are mainly subclinical (Baneth et al. 2013). Nevertheless, infection with this species or a closely related variant has resulted in severe myositis in a wild Pampas gray fox (Giannitti et al. 2012).
In carnivores, vectors have been identified only for two Hepatozoon species: R. sanguineus and Amblyomma ovale for H. canis (Baneth et al. 2001, 2007, Forlano et al. 2005) and Amblyomma maculatum for H. americanum (Mathew et al. 1998). The diversity of Hepatozoon species in carnivores is probably more complex than is presently known because considerable DNA heterogeneity in conserved gene sequences of Hepatozoon from carnivore hosts is frequently seen (Starkey et al. 2013). However, a lack of data on different manifestations of infection, life cycle variations, antigenic variability, and vector competence constitute a serious drawback for further taxonomic work.
Hepatozoon Detection in Blood, Tissues, and Invertebrate Vectors
Due to their complex life cycle, stages of Hepatozoon can be detected in a variety of tissues. In vertebrate hosts, blood is the most commonly used material for intravital diagnostics. Inasmuch as the gamonts are mostly present in white blood cells within mammalian hosts, however, using the buffy coat layer for microscopic and molecular diagnostics increases the sensitivity of detection (Sasanelli et al. 2010, Otranto et al. 2011). Depending on the given Hepatozoon species, life stages can be further detected by cytology or biopsy histopathology in the bone marrow, spleen, lymph nodes, or striated muscle. At necropsy, almost any parenchymal tissue can be used for parasite detection by nucleic acid amplification tests (NAATs). The spleen is nevertheless the primary target organ for the parasite's detection (Baneth et al. 2007). However, the ability of some Hepatozoon spp. to survive in tissues of hosts accidentally infected by ingestion of other infected hosts (e.g., in bats after presumed ingestion of infected arthropods) complicates the interpretation of PCR positivity in tissues (Pinto et al. 2013, Karadjian et al. 2015). Polysporocystic oocysts (or DNA) of Hepatozoon species can also be detected in bloodsucking arthropod vectors. Detection of DNA in questing ticks is highly suggestive for the vectorial role. On the contrary, the presence of Hepatozoon DNA in blood-engorged arthropods should not be considered proof of their vectorial capacity unless mature oocysts are demonstrated microscopically (Giannelli et al. 2013).
Diagnosis by Microscopy
The traditional direct diagnostic methods for rapid detection of Hepatozoon include thick and thin blood smears and buffy coat smear (Fig. 1). These methods are effective only in animals with high parasitemia. Sensitivity is noticeably diminished for subclinical infections and for those Hepatozoon species (such as H. americanum) that typically produce very low parasitemia levels during clinical disease (Vincent-Johnson et al. 1997). Romanowsky stains, including Giemsa stains and such commercial rapid stains as Diff-Quick, are suitable for detecting Hepatozoon spp. in blood. Hepatozoon life stages can be detected also in histopathology sections of tissues demonstrating the architecture of the parasite's tissue stages in a clear manner. Histopathology is also helpful in characterizing inflammatory response to infection with different Hepatozoon species in distinctive hosts (Klopfer et al. 1973, Baneth and Weigler 1997, Vincent-Johnson et al. 1997, Baneth et al. 2013).

Hepatozoon gamonts in white blood cells manifested in blood smears remain the keystone for clinical diagnosis of infected mammalian hosts.
Serological Tests
Serological assays have been developed for detecting antibodies for Hepatozoon spp. in dogs. An indirect fluorescent antibody (IFA) test for anti-H. canis antibodies using gamont antigen has been used in epidemiological studies in Israel, Japan, and Turkey (Shkap et al. 1994, Baneth et al. 1996, Inokuma et al. 1999, Karagenc et al. 2006), and an IFA assay using sporozoite antigens derived from ticks has been developed for detecting anti-H. americanum antibodies. The latter assay was found to be as sensitive as a muscle biopsy in diagnosing this infection (Mathew et al. 2000). Furthermore, an enzyme-linked immunofluorescence assay (ELISA) for anti-H. canis antibodies based on gamont antigen has also been developed (Gonen et al. 2004). A low degree of cross-reactivity was found when using the H. canis ELISA with sera of dogs infected with H. americanum (Gonen et al. 2004). In the past decade, serological tests have largely been replaced by NAATs.
NAATs for Hepatozoon species
Similar to other vector-borne protists, molecular diagnostic approaches relying mainly on the amplification of 18S rRNA gene fragments have prevailed during the past two decades in Hepatozoon detection and genetic characterization in mammals, while studies on reptilian Hepatozoon s.l. use ITS-1 as an additional or alternative target for both diagnostic PCR assays and phylogenetic analyses (Kim et al. 1998, Boulianne et al. 2007). To date, sequences of the 18S rRNA gene comprise the only available genetic data from Hepatozoon species infecting carnivores (Table 1, Fig. 2). The major bulk of sequences originating from different host species are ca 250–700 bp in length, and suggests a monophyletic character of Hepatozoon species from carnivores (Criado-Fornelio et al. 2006, Barta et al. 2012, Najm et al. 2014). However, the variability of the 18S rDNA within Hepatozoon species combined with the short length of the sequences used in some phylogenetic analyses complicate definitions of species status among Hepatozoon spp. (see e.g., Karadijan et al. 2015). Neighbor-joining analysis of partial 18S RNA sequences has revealed five groups of H. canis isolates from dogs based on individual point mutations, thereby demonstrating the variability of H. canis within the microenvironment in Croatia (Vojta et al. 2009). Foxes from the same country showed less variability (Dežđek et al. 2010).
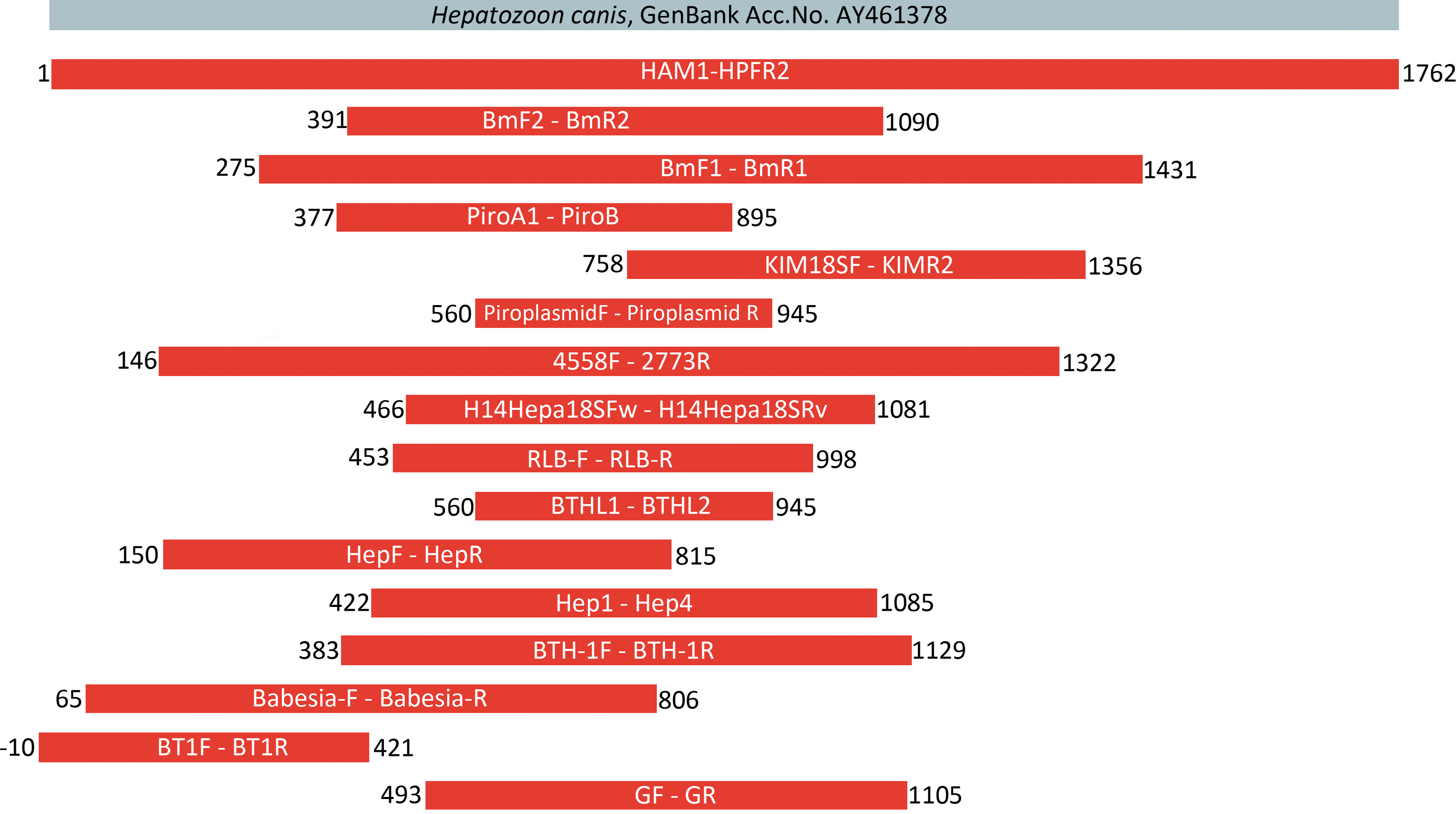
Length and position of 18S rDNA fragments amplified by various primer pairs. The nucleotide positions are extrapolated according to the reference sequence of H. canis AY461378 (shown in gray). Color images available online at
Expected approximate size of PCR product for Hepatozoon spp. sequence.
Primers used in cited literature in nested PCR protocol.
Yes = primers specific only for Hepatozoon spp. sequences; no = primers with broader range of amplified species.
Reference to first use of primers for Hepatozoon spp. sequence amplification, original annotation of primers in brackets.
Sequence of primer HEP-1 was originally published as 5′-CGC GAA ATT ACC CAA TT-3′. As this very probably contains a mistyping error, we suggest here the correct sequence to anneal to Hepatozoon 18S.
Molecular detection of Hepatozoon spp. is highly influenced by the specificity of the primer pair and the PCR cycle conditions that are used. Most of the primers used are universal enough to amplify either a broad range of apicomplexan protists (e.g., BTHL 1 and 2, BTH-1F and −1R, BmF and R) or piroplasmids (e.g., Piroplasmid F and R, Piro A1 and B, BT1F and R). It should be recognized that several primer pairs amplify Hepatozoon 18S despite mismatches in target sequence (GF2 and GR2, BmF and R, Babesia-F and -R) (Table 1). For this reason, it is always recommended to confirm the identity of PCR products by sequencing a representative amount of samples from each particular set. The difference of ca 40 bp in length of the 18S PCR products for piroplasmids and Hepatozoon species can be helpful in distinguishing them when universal primers are used (Fig. 3).
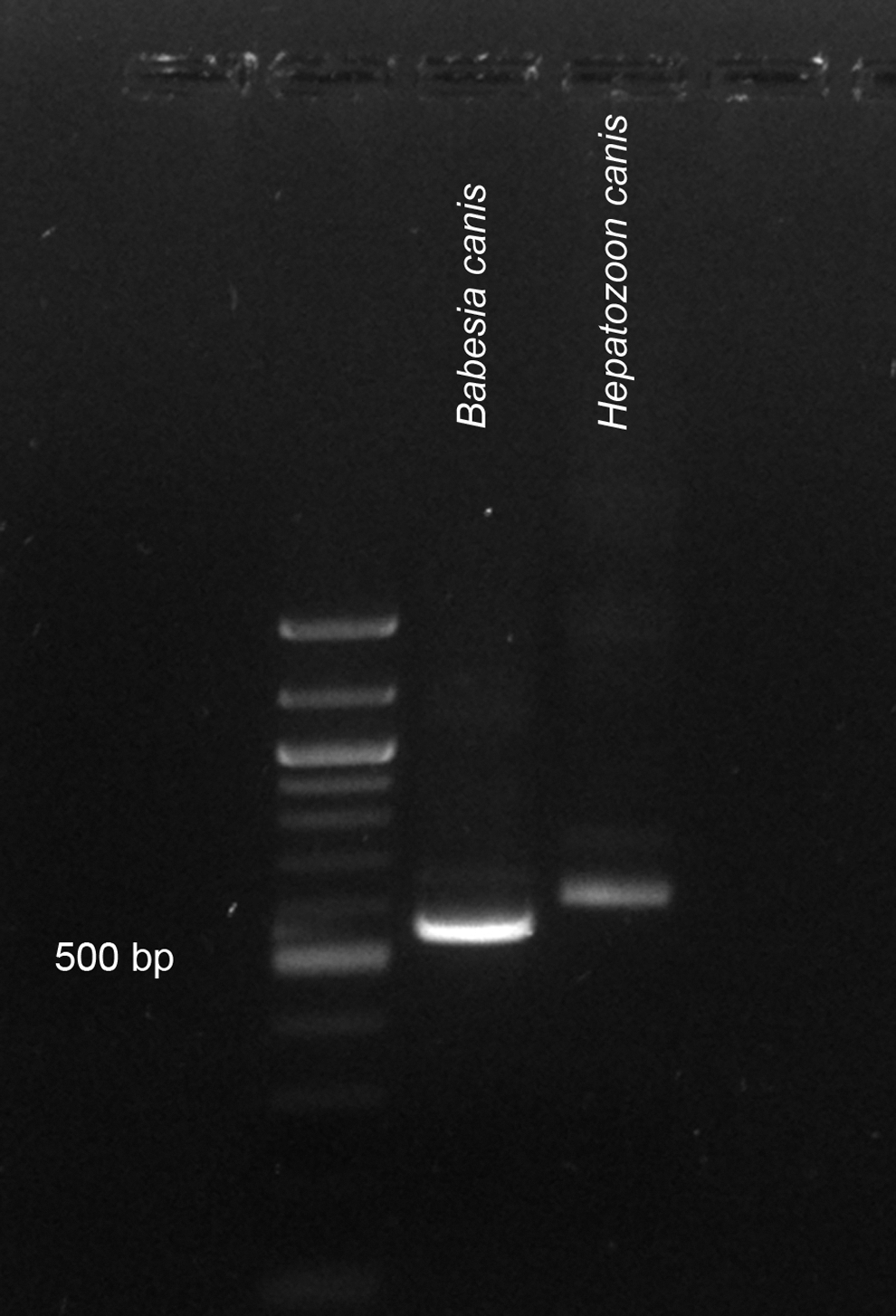
Demonstration of H. canis and Babesia canis 18S PCR products amplified by nested PCR using nonspecific primers (BTH-1F, BTH-1R, and GF2, GR2), showing ca 40 bp length difference between the amplicons; 100 bp DNA Ladder (New England BioLabs).
The choice of primers should reflect the purpose and design of the particular study. For diagnostic PCR, specific primers amplifying short PCR products are recommended (e.g., H14Hepa18SFw and Rv, Hep-1 and -4, HepF and R). High sensitivity can be achieved using a nested PCR protocol (e.g., a combination of universal BTH-1F and −1R with more specific Hep-1 and -4 or H14Hepa18SFw and Rv; Fig. 2), but this may come at the cost of increased contamination risk. For phylogenetic analyses and species determination, amplification of longer fragments (using, e.g., HAM1-HPFR, 4558F −2773) is advisable (Table 1).
Quantitative PCR
In recent years, quantitative PCR (qPCR) has been frequently used in studies because it enables quantification and detection of mixed infections from various biological sources, without laborious evaluation of results by gel electrophoresis. This notably has not been the case, however, for studies of Hepatozoon species. Only two qPCRs assays targeting the conserved 18S rRNA gene have been developed to estimate the prevalence and intensity of Hepatozoon infection in carnivore species (Criado-Fornelio et al. 2007, Li et al. 2008). Using the primers HEP1 and HEP2, Criado-Fornelio et al. (2007) failed to separate species infecting canids and felids by melting peak temperature analysis, while qPCR combined with FRET revealed single target nucleic copies with 100% specificity and enabled differentiation of dog species (H. canis and H. americanum) from canine blood (Li et al. 2008, Kelly et al. 2013). The first and to date only qPCR protocol based on melting curve analysis that enables differentiation between mixed infection of adeleorinid (Hepatozoon s.l.) and eimeriorinid (Schellackia and Lankesterella) protists was developed to study their presence in reptiles (Maia et al. 2014).
Relevance of Various Detection Techniques for Clinical Diagnosis in Dogs and Cats
The assays used to diagnose Hepatozoon infections in dogs and cats include detection of gamonts in blood smears, cytology of tissues, serology, and PCR. While detection by microscopy of blood smears is easy and rapid when parasitemia is sufficient, as in most clinical infections with H. canis, it is not sensitive enough to detect H. americanum and H. felis infections. Furthermore, follow-up treatment for H. canis should be carried out using PCR and buffy coat microscopy because detection by microscopy is considerably less sensitive than is PCR (Sasanelli et al. 2010, De Tommasi et al. 2014). A study from Turkey demonstrated that detection of H. canis by PCR is far more sensitive than is examination of blood by light microscopy. In that study, the prevalence of infection among 349 dogs was 10.6% according to blood smear evaluation and 25.8% according to blood PCR (Karagenc et al. 2006). Additionally, a comparative study on the bone marrow, blood, and buffy coat of dogs infected with H. canis has found that buffy coat and blood are the best tissues for detecting H. canis infection in dogs using PCR (Otranto et al. 2011). Sensitive and specific real-time qPCR assays are able to detect both H. americanum and H. canis while distinguishing between the two species (Criado-Fornelio et al. 2007, Li et al. 2008). Serological assays for canine hepatozoonosis are indicative of exposure to the parasite and can be used for epidemiological surveys, but they lack the sensitivity and specificity of real-time PCR.
Conclusions and Future Perspectives
A plethora of studies employing molecular tools for detecting Hepatozoon s.l. in domestic and free ranging mammals have been published in the past decade. To date, both molecular diagnosis and phylogenetic analyses rely solely on nuclear 18S rDNA data. Extending the range of available molecular markers to plastid or mitochondrial genes will greatly improve the understanding of diversity within the genus Hepatozoon and intergeneric relationships between individual clades of hemogregarines. The recently published mitochondrial genome of Hepatozoon (Bartazoon) catesbianae (Leveille et al. 2014) opened a range of opportunities in the search for alternative phylogenetic markers and diagnostic primers also for mammalian Hepatozoon spp. Inasmuch as the prevalence assessed by PCR in clinically healthy dogs and cats can reach more than 25% in areas of endemic occurrence, and because extremely sensitive diagnostic PCR also detects subclinical carriers, direct demonstration of gamont presence in circulating white blood cells (in blood smears or buffy coat smears) remains an important clinical diagnostic tool.
Footnotes
Acknowledgments
The international collaboration of all authors was facilitated by COST Action TD1303. The University of Veterinary and Pharmaceutical Sciences team was supported by grant COST CZ LD14048 and the project CEITEC 2020 (LQ1601) with financial support from the Ministry of Education, Youth and Sports of the Czech Republic under the National Sustainability Programme II. The microphotographs of Hepatozoon were kindly provided by Martina Gallusová (Fig. 1b) and Andrei Mihalca (Fig. 1c). Barbora Mitková kindly contributed to the review of primers used in the literature (![]() ).
).
Author Disclosure Statement
No competing financial interests exist.