
Abstract
Bartonella species are highly endemic among wild rodents in many parts of the world. Blood and/or blood clot cultures from 38 rodents, including 27 Yucatan deer mouse (Peromyscus yucatanicus), 7 Gaumer's spiny pocket mouse (Heteromys gaumeri), 2 black rats (Rattus rattus) and 2 big-eared climbing rats (Ototylomys phyllotis) captured near Merida, Yucatan, Mexico, led to the isolation in 3–4 days of small gram-negative bacilli, which were identified as Bartonella spp. based on colony morphology. DNA extraction and PCR testing were also performed from heart samples of 35 of these 38 rodents. Overall, Bartonella spp. were isolated from the blood/blood clots of 22 (58%) rodents. All Bartonella-positive rodents were Yucatán deer mice from San José Pituch. Sequencing of a fragment of the gltA gene showed that all but one rodent isolates were closest to B. vinsonii subsp. vinsonii and one isolate was intermediate between B. vinsonii subsp. berkhoffii and B. vinsonii subsp. arupensis. Further analysis of concatenated housekeeping genes (gltA, ftsZ, rpoB, and 16S rRNA) suggests that this outlier isolate is a new subspecies within the B. vinsonii genogroup, for which we proposed the name B. vinsonii subsp. yucatanensis.
Introduction
B
Little is known on the distribution of Bartonella species in wild rodents in Mexico as only a limited number of studies were conducted in that country, mainly in northeastern Mexico (Bai et al. 2009, Rubio et al. 2014, Fernández-González et al. 2015), and no investigation of the presence of Bartonella in mammals has been conducted yet in the specific ecosystem of Yucatan. The objective of the present study was to evaluate the prevalence of Bartonella spp. in small rodents captured near Merida, Yucatan, and determine which Bartonella species were infecting these mammals.
Materials and Methods
Study area and sampling procedures
The rodents were captured at three different sites, ∼100–150 km east and southeast of the city of Merida, Yucatan, in early December 2013 (Table 1). Locations were specifically Peto (20° 7′ 33″ North, 88° 55′ 22″ West); Bokobá (21° 0′ 27″ North, 89° 10′ 49″ West); and San José Pituch (20° 56′ 59″ North, −88° 40′ 29″ West). The vegetation is characteristic of tropical savannah.
Small rodents were trapped with Sherman traps (7.6 × 8.9 × 22.9 cm; H.B. Sherman Traps, Inc., Tallahasse, FL) baited with a mix of oat and peanut butter and placed every 10 meters on each side of a transect. The traps were placed at dusk and checked and emptied at dawn. The live trapped rodents were transported to a laboratory in Merida where they were anesthetized before euthanasia. Rodents were identified to species based on external morphology. Rodents were weighed, measured, and species, sex, and age (adult, juvenile) were recorded. Blood samples were collected by intracardiac puncture before euthanasia and disposed in 2.0-mL plastic EDTA tubes (Becton Dickinson, Forest Lakes, NJ). Blood clots were also collected after euthanasia. Unfortunately, the presence of ectoparasites was not consistently recorded. Blood samples were aliquoted for the present study and for an investigation on the presence of hantaviruses performed by the Pasteur Institute, Paris, France (Noël Tordo, personal communication). Blood and blood clot samples were stored in a −80°C freezer in one of the investigators' laboratory in Mexico City, D.F., Mexico, until shipment to California.
Ethics statement
This study and sampling protocol were approved by the Institutional Animal Care and Use Committee at the School of Veterinary Medicine of the University of California, Davis (approval no. 18367), and permit from SEMARNAT Ministry of Environment and Natural Resources (Secretaría del Medio Ambiente y Recursos Naturales) permiso No-SGPA/DGVS/08283/12. Scientific collection permits were issued by the Mexico Ministry of Agriculture, Livestock, Rural Development, Fisheries, and Food (SAGARPA License FAUT 0250) and the Veterinary and Zootechnical School at the National University of Mexico (UNAM Permit B00.02.02/1026 1708).
Culture and isolation of Bartonella
Whole blood as well as blood clot samples and hearts available from the previous study were stored in a −80°C freezer at the University of California, Davis, until thawed and plated. For each rodent, both whole blood and blood clots were cultured when available. For culture of rodent blood, 200 μL of thawed EDTA blood and M199 growth medium (1:1) was plated onto fresh (<1-week-old) 5% defibrinated rabbit blood agar plates, as previously described (Koehler et al. 1992). For culture of the blood clots, 20 μL of M199 growth medium was plated on the center of a rabbit blood agar plate and was mingled with a small blood clot using a disposable loop and spread out on the plate. Agar plates were incubated at 35°C in an aerobic atmosphere of 5% CO2 for 4–5 weeks. Bacterial growth was monitored every day during the first week and then on alternate days for up to 3 weeks. After 3 weeks, the plates were checked twice a week. Bacterial colonies were presumptively identified as Bartonella spp. based on their morphology. Suspected Bartonella colonies were plated from the original plates onto secondary rabbit blood agar plates and incubated with the same conditions. When bacterial growth was observed on the secondary plates, one part of the colonies was harvested and stored in a freeze medium at −80°C and another part of the colonies was suspended in Tris buffer and kept in a refrigerator until DNA extraction.
DNA extraction
A suspension of Tris buffer and harvested colonies was heated for 15 min at 100°C, followed by a vortex for 10–15 s from the Bartonella cultures. After 5 min of cooling down, the samples were centrifuged at 12,500 rpm at 4°C for 10 min. One hundred microliters of the supernatant containing the genomic DNA to be used as DNA template in the PCR was then pipetted into a sterile 1.5-mL (stock) vial containing 100 μL of 2 × TE buffer. A working solution of 1:10 was made by adding 5 μL of the stock vial into a new 1.5 vial containing 45 μL of TE buffer. These vials were stored in a −20°C freezer. DNA extraction from 35 rodent heart tissues was performed using a Qiagen DNeasy tissue and blood kit (Qiagen, Valencia, CA) according to the manufacturer's instructions.
Conventional PCR
A PCR amplification of the extracted DNA from the heart tissue and cultures was performed using primers, CSH1F and CSH1137, to generate a 379-bp amplicon of the Bartonella citrate synthase gene (gltA) (Norman et al. 1995). For the suspected positive cultures, not only a PCR on the gltA fragment was performed, but the ftsZ (APT0257 and APT0258), rpoB (APT0244 and APT0245), and 16S rRNA (fD1 and rP2) gene regions were also amplified (Regnery et al. 1992, Norman et al. 1995, Renesto et al. 2001, Zeaiter et al. 2002). However, primer sets targeting these other regions were not applied on heart tissue samples. For each PCR reaction, negative and positive controls were distilled water and the strain Bartonella henselae type II (U4 isolate), respectively. Comparison between PCR-positive blood clots or whole blood samples and heart samples was also performed.
Sequencing and phylogenetic analysis
Amplified PCR products were cleaned using a Qiagen QIAquick PCR Purification Kit according to the manufacturer's instructions. Purified PCR products were sequenced at the College of Biological Sciences UCDNA Sequencing Facility at the University of California, Davis. A phylogenetic tree was constructed from gltA sequences using the MrBayes plugin in Geneious version 8.1.7 with a 1,100,000 MCMC and 100,000 burn-in length (Huelsenbeck and Ronquist 2001, Kearse et al. 2012). Similarly, a concatenated phylogenetic tree of four rodent isolates from Yucatan based on concatenated sequences of four housekeeping genes (ftsZ, gltA, rpoB, and 16S rRNA) was constructed. The phylogenetic tree was constructed from partial sequences using a neighbor-joining method with 100,000 bootstrapped replicates using the Tamura distance model in Geneious version 8.1.8. To determine the taxonomic classification of the four rodent isolates, we used the criteria previously proposed by La Scola et al. (2003), in which the gltA and the rpoB gene sequences from each strain were compared with those of validated published Bartonella species or subspecies.
Results
The Yucatan deer mouse (Peromyscus yucatanicus) was the most prevalent rodent species, accounting for 71% (n = 27) of the 38 rodents captured. Seven Gaumer's spiny pocket mice (Heteromys gaumeri), two black rats (Rattus rattus), and two big-eared climbing rats (Ototylomys phyllotis) were also collected (Table 1). A total of 35 hearts, 13 blood samples and 32 blood clots, were available, as shown in Table 2. Among these 38 rodents, 30 (79%) were adults and 8 (21%) were juveniles and 19 (50%) were males.
PCR positive, but sequence was not usable, so it was not counted as a positive result.
J, juvenile.
The Bartonella colonies took 3–4 days to grow on the agar plates. Four (30.8%) of the 13 blood samples and 21 (65.6%) of the 32 blood clots yielded Bartonella suspected colonies. From the 38 rodents, 22 (58%) yielded a positive reaction using the primer set targeting the Bartonella gltA gene region. This percentage was identical when the 16S rRNA region was targeted instead of the gltA, but was 55% (21) when ftsZ and rpoB primer sets were used. Fourteen (40%) of the 35 rodents for which heart tissue was available were PCR positive for the gltA gene. Overall, 5 (62.5%) of the 8 juvenile rodents were Bartonella positive compared with 16 (53%) of the adults and 11 males were Bartonella positive. Neither age nor sex was identified as a specific risk factor.
Of the seven Yucatan deer mice for which both blood and blood clots were available, three were culture positive and three were culture negative for both samples. One culture was positive for only the blood clot for one rodent. There were 30 rodents for which we had both a blood clot and a heart sample (Table 2). Of these, 14 (46.7%) were culture and PCR positive on both samples, 5 (16.7%) were only culture positive from the blood clot, and 11 (36.7%) were negative for both samples. All PCR-positive heart samples were also positive by culture of whole blood or blood clot. Based on the gltA partial sequence, all but one sample clustered together and were identified as B. vinsonii subsp. vinsonii (Fig. 1). A concatenated alignment of the ftsZ, gltA, rpoB, and 16S rRNA genes showed that one isolate was distinct from B. vinsonii subsp. vinsonii, clustering within the B. vinsonii genogroup, but as an outlier from B. vinsonii subsp. berkhoffii and B. vinsonii subsp. arupensis. This isolate corresponds to a new B. vinsonii subspecies, proposed to be named B. vinsonii subsp. Yucatanensis, as it shares more than 96% sequence homology for the gltA gene, ranging from 96.34% with B. vinsonii subsp. arupensis to 98.15% with B. vinsonii subsp. berkhoffii, but less than 95.4% sequence homology for the rpoB gene, ranging from 91.03% for B. vinsonii subsp. arupensis to 92.83% for B. vinsonii subsp. vinsonii (Fig. 2). This isolate was only obtained from rodent blood (Table 2).
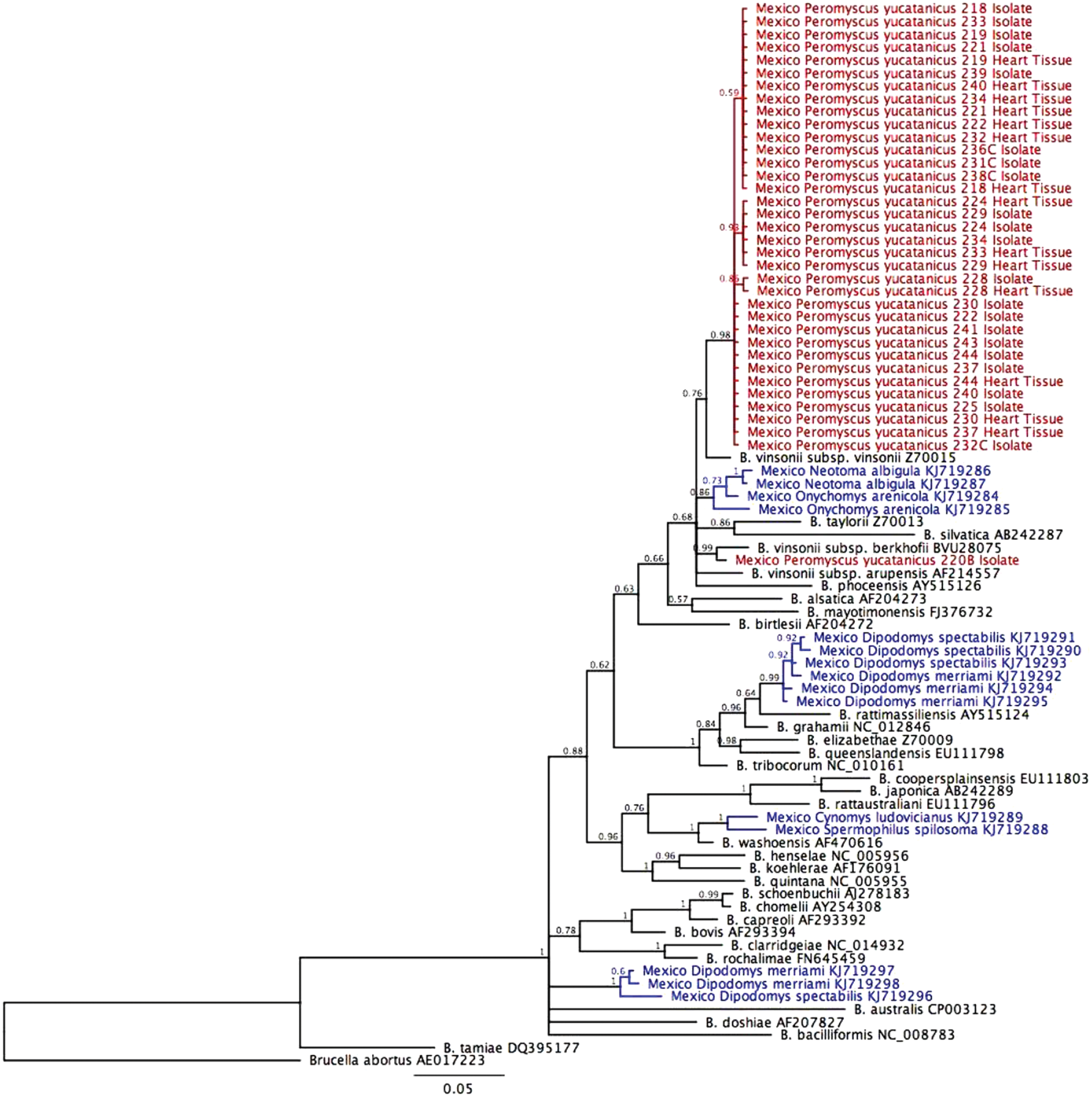
Dendogram of Bartonella strains identified by partial sequencing of the gltA gene. The phylogenetic tree was constructed from gltA sequences on Bayesian neighbor-joining method, using the MrBayes plugin in Geneious version 8.1.7 with a 1,100,000 MCMC and 100,000 burn-in length. Color code: red: this study, blue: data from Rubio et al. (2014), black: referenced Bartonella species. Color images available online at
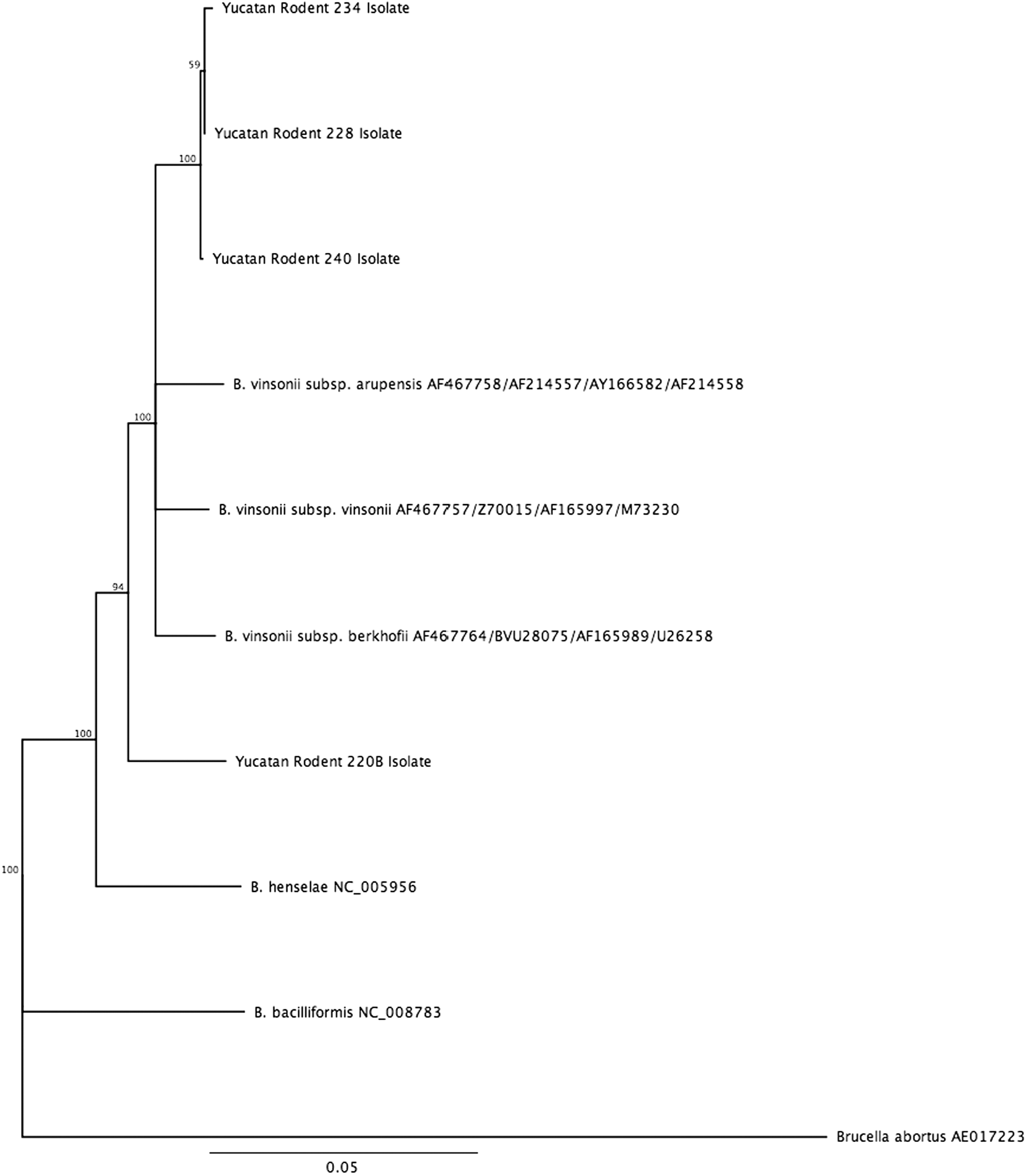
Phylogenetic tree of four rodent isolates from Yucatan based on concatenated sequences of four housekeeping genes (ftsZ, gltA, rpoB, and 16S rRNA). The phylogenetic tree was constructed from partial sequences using neighbor-joining method with 100,000 bootstrapped replicates using the Tamura distance model in Geneious version 8.1.8.
Discussion
The present study reports the identification of B. vinsonii subsp. vinsonii from Yucatan deer mice and the isolation of a new subspecies of B. vinsonii, B. vinsonii subsp. yucatanensis. To our knowledge, this is the first report of the presence of a Bartonella species in this rodent species and in the Yucatan peninsula, Mexico. Despite the small sample size of the present study, the overall prevalence of Bartonella bacteria in deer Yucatan mice was very high (58%), but similar to a recent report from northwestern Mexico with an overall prevalence of 50% in other Peromyscus species (Rubio et al. 2014) and also similarly from other regions of North America (Kosoy et al. 1997, Jardine et al. 2005, Bai et al. 2008). Prevalence varied significantly among rodent host species, which is similar to the pattern observed in other geographic areas; however, numbers of animals tested were quite small for the three other rodent species under investigation. Most of the rodents were trapped in San José Pituch and they were P. yucatanicus, which appears to be the natural reservoir of B. vinsonii subsp. vinsonii in Yucatan. Because of the small number of other rodent species captured, it is not possible to establish their role as Bartonella sp. reservoir. Most of the Bartonella strains isolated from the Yucatan rodents were close to strains isolated from some rodent species in northern Mexico, mainly Neotoma albigula and Onychomys arenicola (Rubio et al. 2014), and also from rodents in western USA (Bai et al. 2008, 2011) and suggestive of adaptation of these Bartonella species to their specific hosts. Therefore, B. vinsonii subsp. vinsonii appears to be widely distributed in various rodent species in several regions of Mexico. Several rodent-borne Bartonella species detected in northern Mexico were not detected in Yucatan, likely because of our limited sample size and by the fact that most of our rodents belonged to one species not reported in the northern Mexico study. The fact that we had to share samples for different studies did not allow testing systematically each rodent for the presence of Bartonella in whole blood, blood clots, and heart tissues.
Most rodent-borne Bartonella spp. are flea-borne (Reeves et al. 2007), and likely the rodents captured in our study must have been flea and tick infested. For instance, 40.4% of the fleas collected from rodents captured in northeastern Mexico were Bartonella spp. PCR positive (Fernández-González et al. 2015). Phylogenetic analyses of gltA gene sequences revealed 13 genetic variants clustering in 5 phylogroups (I–V), 2 of which were matched with known pathogenic Bartonella species (B. vinsonii subsp. arupensis and B. washoensis) and 2 that were not related with any previously described species or subspecies of Bartonella.
Culture from blood or blood clots was more sensitive than DNA extraction from heart samples as five heart samples were PCR negative, but culture and PCR positive for blood, indicating that most of the rodents tested were bacteremic at the time of their capture and bacteria had not yet specifically localized in organs such as hearts, as reported for Bartonella in other reservoir hosts such as coyotes (Kehoe et al. 2014). However, this observation will need to be strengthened by further studies as we had to share samples for different studies, which did not allow testing systematically each rodent for the presence of Bartonella in whole blood, blood clots, and heart tissues.
In conclusion, we report the infection of Yucatan deer mice with Bartonella vinsonii subsp. vinsonii and a new subspecies in the B. vinsonii genogroup, B. vinsonii subsp. yucatanensis. It brings a better understanding of the distribution of this rodent-borne bacterium among rodents in North and Central America.
Footnotes
Acknowledgment
This project was partially funded by a 2013 collaborative research grant from U. C. Mexus-CONACYT.
Author Disclosure Statement
No competing financial interests exist.