
Abstract
Tula virus (TULV) is a zoonotic hantavirus. Knowledge about TULV in the Netherlands is very scarce. Therefore in 2014, 49 common voles (Microtus arvalis) from a region in the south of the Netherlands, and in 2015, 241 common voles from regions in the north of the Netherlands were tested with the TULV quantitative RT-PCR. In the southern region, prevalence of TULV was 41% (20/49). In the northern regions, prevalence ranged from 12% (4/34) to 45% (17/38). Phylogenetic analysis of the obtained sequences showed that the regions fall within different clusters. Voles from the south were also tested on-site for the presence of hantavirus antibodies, but serology results were poorly associated with qRT-PCR results. These findings suggest that TULV may be more widespread than previously thought. No human TULV cases have been reported thus far in the Netherlands, but differentiation between infection by TULV or the closely related Puumala virus is not made in humans in the Netherlands, thus cases may be misdiagnosed.
Introduction
H
The various hantaviruses in Eurasia are strongly linked to a predominant species of rodents and insectivores as their natural reservoir (Plyusnin and Morzunov 2001, Zhang 2014). Tula hantavirus (TULV) has the European common vole (Microtus spp.) as the main reservoir, but TULV has also been found in several other rodent species of the subfamily Arvicolinae (Schlegel et al. 2012, Schmidt et al. 2016). The species TULV was first described in Microtus arvalis and Microtus levis, caught in 1987 in Tula in Russia (Plyusnin et al. 1994). It has been found in several European regions, ranging from Central Europe (Plyusnin et al. 1995, Korva et al. 2009, Schlegel et al. 2012) to Belgium (Heyman et al. 2002). Although zoonotic transmission of TULV has been demonstrated, only few clinical patients have been described in literature. In 2002 an apparent case of TULV infection was reported (Schultze et al. 2002, 2003, Clement et al. 2003), in 2003 a case was reported in Germany (Klempa et al. 2003), in 2012 TULV was described in an immunocompromised patient (Zelena et al. 2013), and recently, TULV infection was described in France (Reynes et al. 2015). In The Netherlands, hantavirus diagnostics is primarily based on serologic testing. Due to the difficulty to differentiate between Puumala hantavirus (PUUV) and TULV infections based on serology, it may be that TULV infection is underdiagnosed.
TULV in the Netherlands was first reported during a study into the circulation and population structure of hantavirus in wild rodents (Reusken et al. 2008). One common vole from Limburg (June 2007) and two common voles from Twente (July 2007) tested positive. No follow-up was performed on these findings. Apart from this one description of TULV in common voles in Limburg and Twente, no information is available about the prevalence of TULV in these areas, or the occurrence of TULV in other areas in the Netherlands.
In 2014, a survey on rodent-borne pathogens was performed in the south of the Netherlands (province of Limburg), close to the location where TULV was previously found. Therefore, TULV was included in the study with the aim to assess the prevalence of TULV in the rodent population. Building on to the results of Limburg, TULV was also included in 2015 in a study on rodent-borne pathogens in the north of the Netherlands (province of Friesland) during an outbreak of common voles.
Material and Methods
Rodents originating from Limburg
In the autumn of 2014, common voles were captured by the Dutch Mammal Society with live traps in the area of Sibbe and Vilt in South-Limburg (Fig. 1). Trapping was done with 60 traps in the habitat of the common vole using Longworth traps (Bolton, Inc., United Kingdom). Prebaiting was used to enhance the trapping index. The trapped rodents were anesthetized with isoflurane (IsoFlo, Abott labs Ltd, England) and serum was collected, after which they were euthanized and dissected. Lungs were collected for TULV research, and were stored in RNAlater (Ambion, Inc., Austin, TX) according to the manufacturer's instructions and at second instance transferred to −80°C. All animals were handled in compliance with Dutch laws on animal handling and welfare (Animals Ethics Committee permission 201000139).
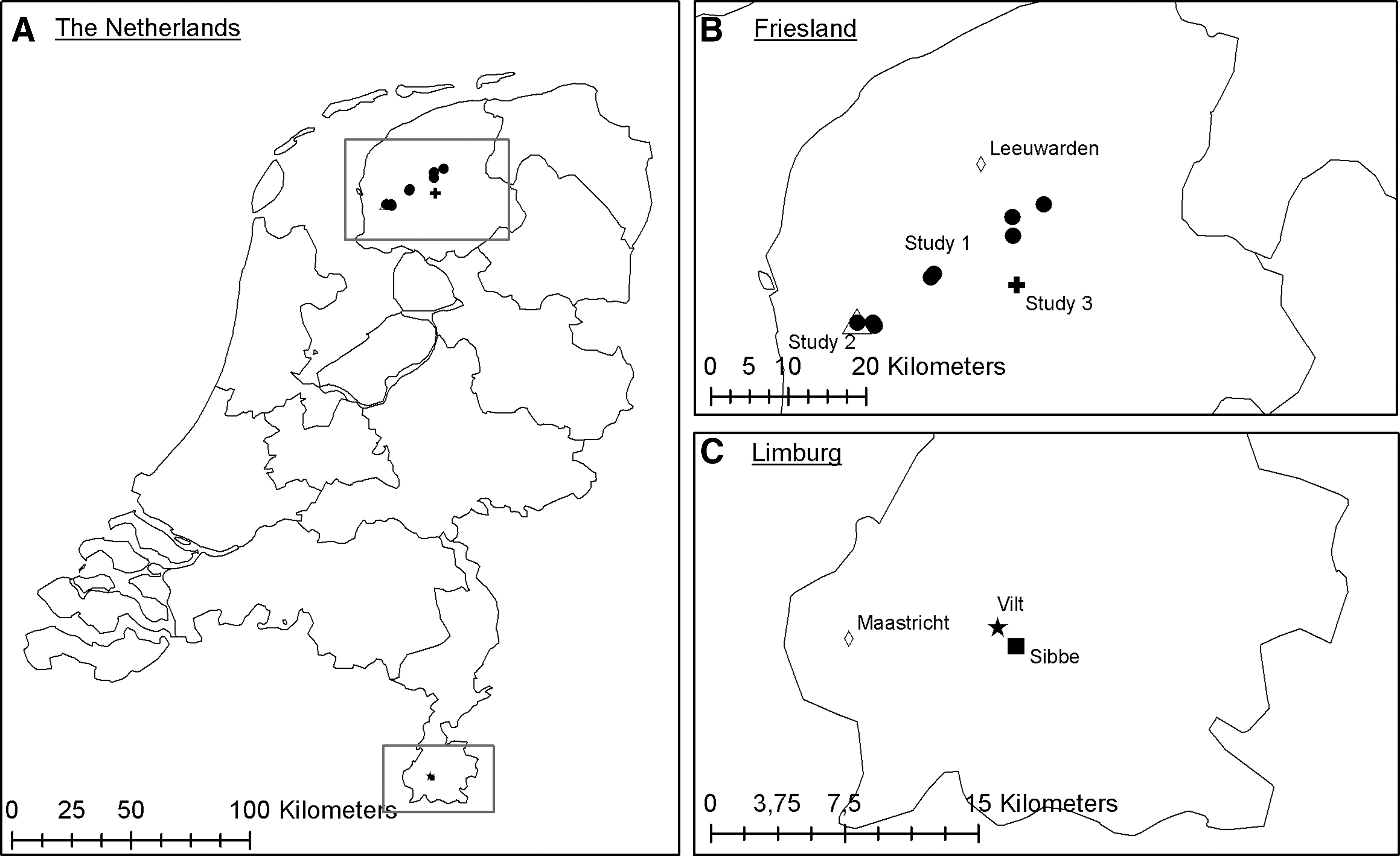
Overview of capture locations of the common voles.
All common voles were tested on-site for the presence of hantavirus antibodies. The screening was done using a PUUV-IgG immunochromatography test (Reascan™ Ab-Dect PUUMALA IgG test (Oy Reagena Ltd., Toivala, Finland)) according to the manufacturer's instructions, using 10 μL whole blood. This test is developed to detect PUUV, but also cross-reacts with TULV (Reusken et al. 2008). In addition, from all voles, total RNA was isolated from lung tissue and used to confirm the presence of TULV RNA by qRT-PCR.
Rodents originating from Friesland
In 2014 and 2015, with a peak in the winter of 2014–2015, common vole populations reached high densities in the province of Friesland, leading to population control measures in certain areas to avoid damage to the land (Wymenga et al. 2015). Common voles were collected in this period from different areas in Friesland (Fig. 1). Study 1 consisted of common voles from nine controlled areas, captured from February to April 2015 for ecological research using snap traps as described in Hörnfeldt (1978) and Beemster and Dijkstra (1991). They were stored at −20°C, and were subsequently sent to the RIVM where they were dissected. Study 2 (Feytebuorren) consisted of common voles that were caught during field work in April 2015 for pathogen research. Vole densities were high enough to allow manual capture, and voles exiting their burrows were captured with buckets. Subsequently, they were anesthetized with isoflurane (IsoFlo, Abott labs Ltd, England), euthanized, and dissected. All animals were handled in compliance with Dutch laws on animal handling and welfare (Animals Ethics Committee permission 201000139). Study 3 (Nes) was performed in August 2015 as part of the rodent population control in the area, using the method described for study 2. The voles were stored at −20°C, after which they were sent to RIVM were they were dissected.
In all three studies, lungs were collected and stored in RNAlater (Ambion, Inc., Austin, TX) according to the manufacturer's instructions and at second instance transferred to −80°C. From all common voles total RNA was isolated from lung tissue and used to detect TULV RNA using a qRT-PCR. Because serology results from common voles in Limburg were poorly associated with qRT-PCR results, no serologic test was performed on-site in Friesland.
RNA isolation and TULV qRT-PCR
Lung tissue samples obtained from voles in Limburg were disrupted using liquid N2 and pestles, and RNA was extracted using RNeasy isolation minikit (Qiagen®), including a Qiashredder step for homogenization and DNase I treatment. Negative control samples were included as every fifth sample. Lung tissue samples from voles from Friesland were homogenized using MagNA Lyser Green Bead tube (Roche Diagnostics, GmbH, Mannheim, Germany) with 500 μL lysis buffer (MagNa Pure 96 Total Nucleic Acid Isolation kit) on a FastPrep FP120 homogenizer (Thermo Fisher, Waltham, MA) two times (40 s at max speed). Total nucleic acid was isolated using a MagNa Pure 96 Total Nucleic Acid Isolation kit on a MagNA pure 96 (Roche Diagnostics, GmbH, Mannheim, Germany). Negative control samples were included as every 10th sample. Comparison of these two methods showed higher Cq values for the MagNa method (on average four cycles) (results not shown), but due to the high viral RNA concentrations this was considered not to affect the results. Quality control of the RNA isolation and inhibition control was performed with β-actin RT-PCR using primers Actin-Fw and Actin-Rv as described earlier (Pinelli et al. 2007).
RNA from lung tissue was tested by the qRT-PCR according to the method described by Kramski et al. (2007), using a TULV-specific probe. To obtain sequence data for phylogenetic analysis, RT-PCR for the PUUV/TULV partial S-segment was performed according to the protocols described by Essbauer et al. (2006). All fragments were purified with ExoSAP-IT PCR clean-up (Isogen Life Science, the Netherlands) and sent to Baseclear B.V. (Leiden, the Netherlands) for sanger sequencing.
The likelihood of being TULV RNA positive was analyzed using a generalized linear mixed model (GLMM), with sex (M/F), location (Limburg/Friesland), and weight as explanatory variables. As random effect, we allowed the intercept to vary over the study areas, which represents possible variation in prevalence over the areas or variation due to different trapping schemes. Data analysis was performed using R, version 3.2.0.
Estimation of phylogeny
Nucleotide sequences were compared with sequences of the TULV partial S-segment from GenBank, after subtraction of the primer sequences. Sequences were aligned using MAFFT, version 7.271 (Katoh and Standley 2013). The option localpair was on, and the option maxiterate was set to 1000. A phylogenetic tree was estimated using raxml, version 8.1.17 (Stamatakis 2014). A general time reversible model of DNA evolution and CAT approximation for a gamma model of rate heterogeneity (-m GTRCAT) were selected. Options for rapid bootstrap analysis and maximum likelihood tree search (-f a) were set to compute confidence values for each internal node.
Results
To determine the prevalence and geographic distribution of TULV in common voles in the Netherlands, rodents were captured and tested for the presence of TULV-specific antibodies and/or viral RNA. In Limburg, 49 common voles were captured: 36 near the town Sibbe, four near the town Vilt, and nine from either Sibbe or Vilt. Three common voles showed a hantavirus-specific IgG response, two voles had borderline results. When tested with qRT-PCR, 20 out of the 49 (40.8%) common voles tested positive, of which 14/36 from Sibbe, 4/4 from Vilt, and 2/9 from Sibbe or Vilt (Tables 1 and 2). In Vilt, one field vole (Microtus agrestis) was also captured and was found positive for TULV in qRT-PCR. The presence of TULV RNA was used for subsequent analyses.
Including the two voles with doubtful responses.
Since we did not find a correlation between seroprevalence, with the serological method used in this study, and prevalence of TULV RNA in voles in Limburg, we decided to only screen by qRT-PCR in follow-up studies. In Friesland, common voles originated from three different captures. Study 1 consisted of 181 common voles coming from nine locations, all within the province of Friesland (Fig. 1). Study 2 and 3 both had 38 voles. RNA was successfully isolated and amplified of respectively 169, 38, and 34 voles of these studies. Of study 1, 70/169 (41%) tested positive, of study 2 17/38 (45%), and of study 3 4/34 (12%) (Table 2).
Though for some locations specific data were missing, for example, for common voles from study 1 and 3 no weight was available, neither was age class always recorded, the GLMM showed clearly that the only significant variable for TULV RNA positivity was weight (p < 0.001). It should be borne in mind that weight may be a proxy for other traits, for example, as voles grow heavier when they age, weight could very well indicate age.
Phylogeny
To determine the genetic variation and geographic clustering of TULV strains in the Netherlands, phylogenetic analysis using the amplified PCR products of the partial S-segment was performed on eight of the samples from Limburg and Friesland and on historical samples from Twente and Limburg, described in Reusken et al. (2008). The sequences from Limburg and Friesland are clearly separated (Fig. 2). The historic sequences of Limburg fall into the same section as the Limburg sequences from the current study. The data were too few to draw conclusions on the mutation speed. Within the regions of Limburg and Friesland, no clear separation is visible between the different capture locations (Limburg: Sibbe and Vilt; Friesland: A&W, Feytebuorren and Nes).
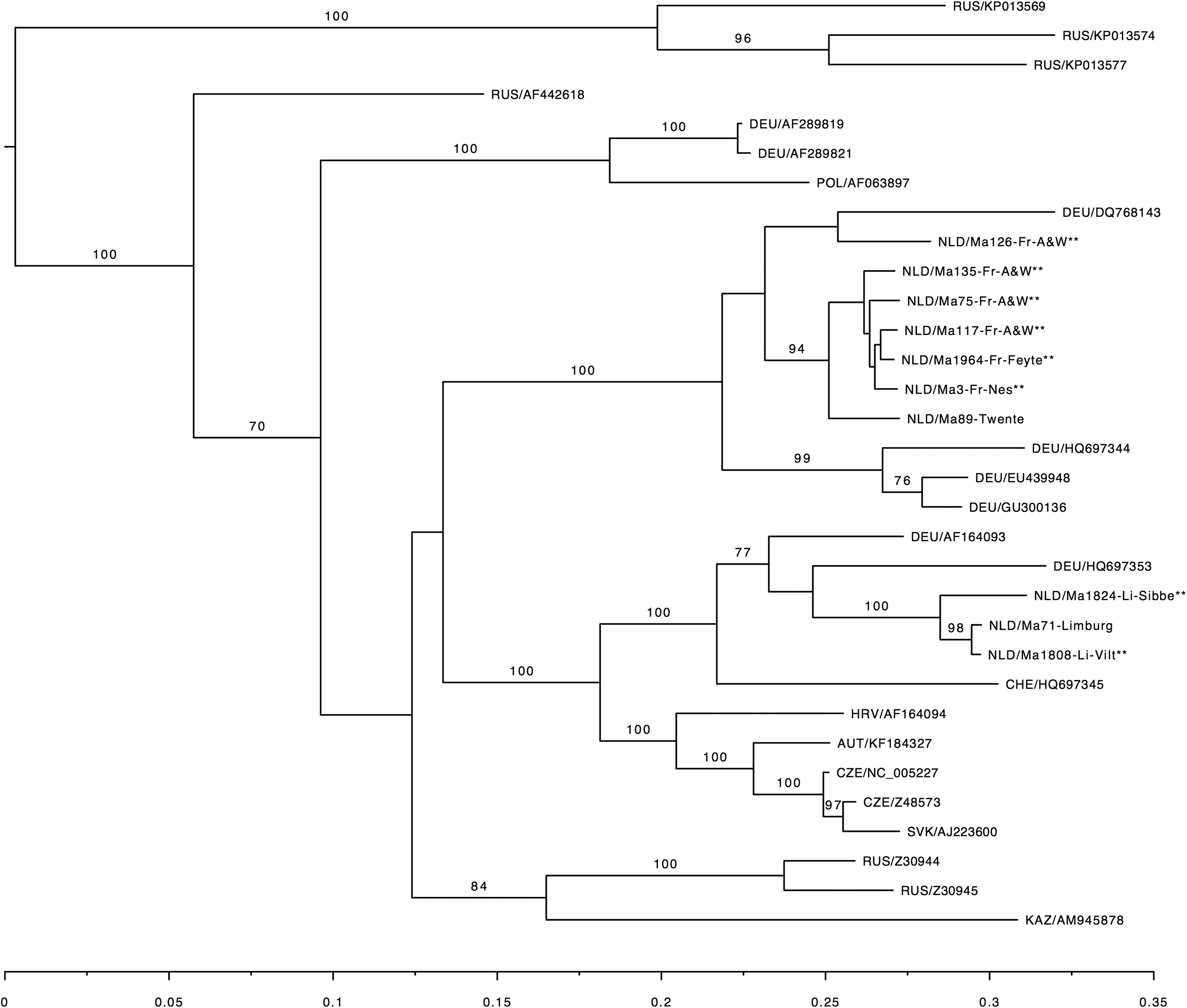
Maximum-likelihood phylogenetic tree of TULV sequences from the investigated common voles from the Netherlands compared to other European TULV sequences. The phylogenetic tree is based on the partial S-segment (nt 342–1102). Sequences were aligned using MAFFT and raxml was used to estimate the phylogenetic tree. Novel sequences are indicated with ** and are labeled with country code, host species, individual code, and study area. Published sequences from GenBank are labeled with country code and accession number. Bootstrap support is indicated for major branches.
Discussion
This study describes the detection of TULV in common voles at two different locations in the Netherlands. In 2014, a study was performed in South Limburg and in 2015, common voles from three different studies in Friesland were tested.
The prevalence of TULV RNA in common voles for the four locations in this study ranged from 11.8% to 44.7%. Other studies report TULV RNA prevalence in common voles of 0% to 37.5% in various regions in Germany, Luxembourg, and France (Schmidt et al. 2016) and 37% in Hungary (Jakab et al. 2008). The local prevalence of >40% in Limburg and two locations in Friesland is reason for further investigation into the apparent favorable conditions for TULV infection in common voles in these regions. The relatively high prevalence may also be due to the high vole densities that were present at the time of those studies, which may have increased contact rates and thus facilitated transmission of the virus through the population. Since the methods applied varied slightly between Limburg and Friesland, comparisons between the regions should be made with caution.
For the on-site detection of TULV infection in Limburg, the Reascan test was used. This assay is developed for the detection of PUUV, but cross-reactivity has been described for TULV (Reusken et al. 2008). However, our results show that 17 out of 20 qRT-PCR positive common voles were missed with the initial on-site screening. The low sensitivity implies that the cross-reactivity may be less than previously thought, which confirms other studies that show low cross-reactivity between TULV and PUUV antibodies in voles, in contrast to sera from humans (Plyusnin et al. 1994, Tegshduuren et al. 2010). The apparent lower sensitivity of TULV serology compared to RT-PCR was also recently described by Schmidt et al. (2016). The low sensitivity of the serologic screening may also arise from a delayed seroconversion, as was described for natural infection with PUUV (Voutilainen et al. 2015). However, the relative large number of voles it concerns makes it unlikely that this is the main reason for the low test sensitivity.
Heroldova et al. (2010) report that the prevalence of TULV antigen increased with vole population numbers. They also report no significant differences in prevalence between men and women, but heavier individuals were more often positive. There was a significant effect of season, as overwintering individuals were found more often positive in early spring. This corresponds to the finding of Deter et al. (2008) that reports that TULV-seropositive common voles were mainly older voles. In our study, also no significant differences were found between men and women, but weight had a significant influence on TULV positivity.
In a study on rodent host specificity of European hantaviruses, field voles could not readily be infected with TULV and no potential spillover infections were found in wildtrapped rodents in an area in which TULV was found (Klingström et al. 2002). During our field study in Limburg, also rodent species other than common voles were captured and tested for TULV. One field vole (M. agrestis) was captured and was found positive. In the phylogenetic analysis, the TULV-positive field vole fell in the same phylogenetic TULV group as the common voles from that region (results not shown). This confirms other findings, for example, from Schmidt-Chanasit et al. (2010), who detected TULV antibodies in both M. arvalis 19% (70/377) and M. agrestis 12% (6/50). In a Slovenian study, different TULV sequences were found in different rodent hosts, and from different country regions, with sequences clustering together based on geographical origin and not on a different rodent host (Korva et al. 2009).
The finding of high TULV prevalence at two locations in the Netherlands, suggests that TULV may be much more widespread and common than previously thought. Though specific antibodies against TULV have been identified in healthy humans (Vapalahti et al. 1996, Mertens et al. 2011), and a few human patients have been described, the pathogenic potential of TULV is less well known than of other hantaviruses such as PUUV and Seoul virus. No human cases of TULV infection have been reported yet in the Netherlands, but human PUUV cases occur several times in the year. Hantavirus diagnostics is based on serology in the Netherlands, and the differentiation between TULV and PUUV infection in humans is not made, thus it is unknown whether there are incorrectly diagnosed PUUV cases that are actually caused by TULV. Studies differentiating between PUUV and TULV are needed to determine the true prevalence of TULV in humans and to determine its pathogenicity.
Our results show that the TULV partial S-segment sequences cluster within two separate regions. Geographical clustering has been reported in various studies. The sequences found in Limburg cluster with the sequences from, for example, Southern Germany, Luxembourg and the Alsace region from France, described as cluster III by Schmidt et al. (2016). The sequences found in Friesland and the historical sequence from Twente cluster with German sequences, described as cluster 1a and 1b (Schmidt et al. 2016).
Conclusions
We have shown that TULV is present in two different regions in the Netherlands at a high prevalence. Subsequent studies should be performed to assess whether other areas in the Netherlands have an equal high prevalence, and longitudinal studies in Limburg and Friesland could provide information about fluctuations in the TULV prevalence. No human cases have been reported yet in the Netherlands, however, development of a serological assay that allows differentiation between TULV and PUUV-specific antibodies will be necessary to determine the prevalence in humans.
Footnotes
Acknowledgments
We would like to thank Dick Bekker, at that time working at de Dutch Mammal Society, for his contribution to the field work in Limburg and Nico Beemster from Altenburg and Wymenga ecological consultants for his contribution to the capture of common voles in Friesland. Postmortem research was performed on-site by Tanja Schouten and Angela Gommersbach. We thank Sanne van den End for her help with the postmortem research in Limburg.
Author Disclosure Statement
No competing financial interests exist.