
Abstract
Background:
Cercarial dermatitis or swimmer's itch is a skin condition in humans due to the larval forms of bird schistosomes of some species of the genus Trichobilharzia. The life cycle of these schistosomes requires freshwater snails (intermediate host) and waterfowl (definitive host). Repeated exposures to cercariae can lead to skin sensitization with the induction of pruritic skin lesions.
Methods:
We describe, in this study, two outbreaks of human cercarial dermatitis at the Eau d'Heure Lakes, Belgium. In July and August 2012, a total of, respectively, 78 and 10 people reported a sudden skin rash accompanied by pruritus following recreational activities in the Plate Taille Lake. However, no ocellate furcocercariae were detected following light exposure of the snails collected between September 2012 and September 2013 (n = 402). No outbreaks were recorded in 2013 and 2014. In August 2015, about 30 new cases were recorded. Snails were collected (n = 270) in different locations around the lake. PCR was used to identify accurately the intermediate hosts and the parasite species involved.
Results:
After light exposure, seven Radix spp. (2.6%) shed ocellate furcocercariae. Molecular identification based on the rDNA internal transcribed spacer 2 sequence ascribed the infected snails to R. balthica ( = R. peregra = R. ovata) (6/7) and R. auricularia (1/7). Based on the amplification of the D2 domain of the 28S rDNA, the cercariae were shown to belong to two different haplotypes of Trichobilharzia franki.
Conclusions:
This is the first record in Belgium of T. franki and associated skin condition.
Introduction
T
The penetration of the human skin by the parasitic larvae, an ocellate furcocercariae, is responsible for an allergic response and can lead to a severe skin condition. Indeed, in humans engaged in recreational or professional water activities, repeated exposures to cercariae (a free-living trematode stage) can lead to skin sensitization with the induction of pruritic skin lesions, reddening of the skin, and macular eruptions; in some individuals, fever, edema, and local lymph node swelling may occur (Kolarova et al. 2013). Nevertheless, the cercariae are not able to complete their life cycle within these accidental hosts, and most die soon after penetration (Horak and Kolarova 2005, Horak et al. 2008). Thus, swimmer's itch is considered essentially not harmful, but very unpleasant.
This seasonal pathology is regarded as an emerging or reemerging disease worldwide because it is increasingly reported (Horak and Kolarova 2011). Outbreaks can lead to the closing of swimming areas and consequently to major financial losses for touristic operators. Several foci of HCD are well known in Europe, for example in Germany (Selbach et al. 2015), France (Jouet et al. 2008), the Netherlands (Schets et al. 2008), and Poland (Zbikowska 2004). It is worth mentioning that in Belgium, to the authors' knowledge, no cases were ever recorded in scientific literature. The aim of this study is to describe two presumptive outbreaks of HCD in Belgium and identify the responsible intermediate host and causative agents in a double approach as it was previously done in France (Ferte et al. 2005).
Materials and Methods
Sampling location
The Eau d'Heure Lakes are composed of five different artificial water bodies (Plate Taille, Falemprise, Eau d'Heure, Féronval, and Ry Jaune) resulting from the building of two dams in the 70s (Fig. 1). The area covers 1.800 hectare (1/3 water surface, 1/3 woodland, and 1/3 meadows). The Plate Taille Lake is closed by a dam used for power supply and consequently the depth of the lake can vary considerably, which is not the case at Falemprise, Féronval, and Ry Jaune. In the early 2000s, a recreational area comprising a series of cottages and touristic facilities was built up at the Plate Taille. It includes a small shallow swimming area (50°11′21.01″N; 4°21′15.30″E, 250 m a.s.l.) available for the tourists (maximum depth of 2 m, ±2.500 m2).
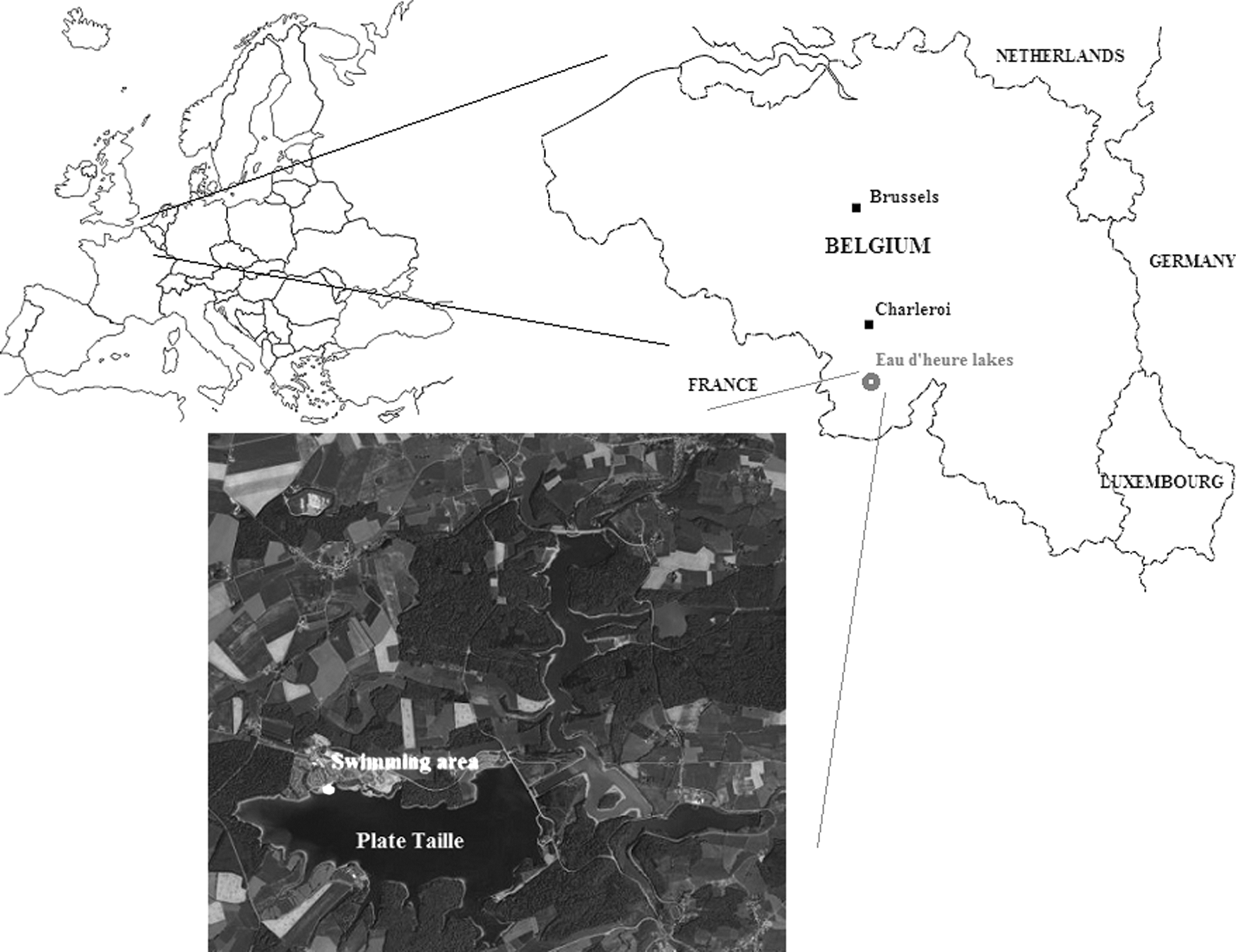
Geographic location of the Eau d'Heure Lakes.
Bathing season is from June 15 to September 15. The lakes are visited by many different species of migrating birds in winter, including different species of Anatidae (Anas platyrhynchos, Cygnus cygnus, Aythya nyroca, Mergellus albellus, Anser fabalis, Anser anser, Anas strepera, Anas acuta, Somateria mollissima, Melanitta nigra, Melanitta fusca, Mergus serrator, Mergus merganser). Mallards (Anas platyrhynchos) are particularly abundant and formed more or less permanent resident populations.
2012 outbreaks
On July 27th, 2012 the weather was hot and sunny and a lot of people occupied the swimming area at Plate Taille. A total of 78 people reported a sudden skin rash accompanied by itching. A few people were transported toward the nearest hospitals. Initially, cyanobacteria (algal bloom) were suspected as etiological agents and the swimming area was closed. However, the different samples were found negative for cyanobacteria toxins and the swimming area was reopened on August 2nd. On August 19th, there was a second outbreak involving 10 people. No cases were reported at Falemprise, the second swimming area. On September 21st, our team visited the swimming areas. Snails and aquatic macrophytes were regularly collected in both recreational areas in September and October 2012 and during the whole bathing season in 2013 at the Plate Taille. No other HCD cases were recorded in 2013 and 2014.
2015 outbreak
On August 30th, 2015, the weather was also hot and sunny and 30 new HCD cases (Fig. 2) were recorded in Plate Taille in an area where swimming was forbidden (over 2 km away from the swimming area). Following this outbreak, several hundreds of snails were collected in September, by hand, from different locations around the lake.

Pruritic macular eruptions in a young man during 2015 outbreak (La Nouvelle Gazette©2015).
Stimulation of furcocercarial shedding by light exposure
Several hundreds of snails (mainly of the genera Lymnaea and Radix) were collected during both outbreaks, transported to the laboratory, and cercarial shedding was stimulated through light exposure as previously described (Ferte et al. 2005). Briefly, pools of 10 snails were deposited in 9 cm diameter Petri dishes and cercarial emergence was stimulated by lighting for 1 h in the laboratory. Snails from positive batches were individually isolated for a second assay. Screening of cercariae was performed using morphological criteria (Combes et al. 1980). Furcocercariae with pigmented eye spots were preserved in 95% ethanol and frozen (−20°C) until DNA extraction. Positive snail hosts were frozen directly at −20°C for storage. Only the samples from which both cercariae and corresponding snails were sequenced were used for molecular analyses.
Molecular analysis of cercariae and positive snails
Sequencing of the D2 domain of the 28S ribosomal DNA (rDNA) was used for the identification of furcocercariae from naturally infected snails. Internal transcribed spacer 2 (ITS-2) was used at the species level for snail identification. DNA extraction was performed using the DNeasy® Blood & Tissue Kit (Qiagen, Hilden, Germany) following the manufacturer's instructions. During the first step, five furcocercariae or a small part of the foot of each positive snail was used. DNA was eluted in 50 μL of water. PCR was performed in a 50-μL volume using 5 μL of DNA and 50 pmol of each of the primers. The sequences were amplified using a commercial kit Taq PCR Master Mix (Qiagen, Hilden, Germany) in a Peltier Thermal Cycler (MJ Research). A protocol previously described (Jouet et al. 2008, 2009) was followed. The rDNA of furcocercariae was amplified using the primers previously described (Mollaret et al. 1997): C2′B (forward) (5′-GAA AAGTACTTTGRARAGAGA-3′) and D2 (reverse) (5′-TCC GTGTTTCAAGACGGG-3′). For the amplification of the positive snails, a primer pair, previously described (Jouet et al. 2008), was used: DIX1(forward) (5′-CGCGCTCTG GWCCKTCGCGGC-3′) and DIX2 (reverse) (5′-ATYTYG TYCGATTTGAGGTTG-3′) to amplify the ITS-2 of Radix spp.
For cercariae, initial denaturation at 94°C for 3 min was followed by 40 cycles of denaturation at 94°C for 45 s, annealing at 50°C for 45 s, and extension at 68°C for 2 min with a final elongation time of 10 min at 68°C. For snails, initial denaturation at 94°C for 3 min was followed by 35 cycles of denaturation at 94°C for 45 s, annealing at 57°C for 30 s, and extension at 68°C for 1 min, with a final elongation time of 10 min at 68°C. PCR products were purified using the commercial kit QIAquick® PCR Purification kit (Qiagen, Hilden, Germany) following manufacturer's recommendation.
Cycle sequencing reactions were performed (in duplicate and in both direction) by BigDye terminator v3.1 (3730 DNA analyzer; Applied Biosystems) by GIGA Genomics Facility (Liège University, Belgium). Consensus sequences were made according to the results of sequencing of the PCR products and were aligned using BioEdit 7.1.10 (Hall 1999) and analyzed using BLASTN 2.2.26 searches in GenBank (
Results
Outbreaks 2012
A total of 402 snails were collected from September 2012 to September 2013 (96% belonging to Radix spp. and 4% to L. stagnalis). Emergence of different types of cercariae (gymnocephalous cercariae, xiphidiocercariae, longifurcate furcocercariae, and echinostome cercariae) belonging to several trematode families was recorded, but no ocellate furcocercariae were identified.
Outbreak 2015
Two hundred seventy lymnaeids snails were collected in September (99% belonging to Radix spp. and 1% to L. stagnalis). A massive emergence of cercariae belonging to several trematode families was recorded at the Plate Taille and ocellate furcocercariae were also identified (only in Radix spp. snails). The eye spots were located on the dorsal part of the body, between the spherical acetabulum and the anterior organ. The cercariae were apharyngeate, with a subterminal mouth and a bifurcate intestine (Fig. 3). Seven snails (2.6%) shed ocellate furcocercariae.
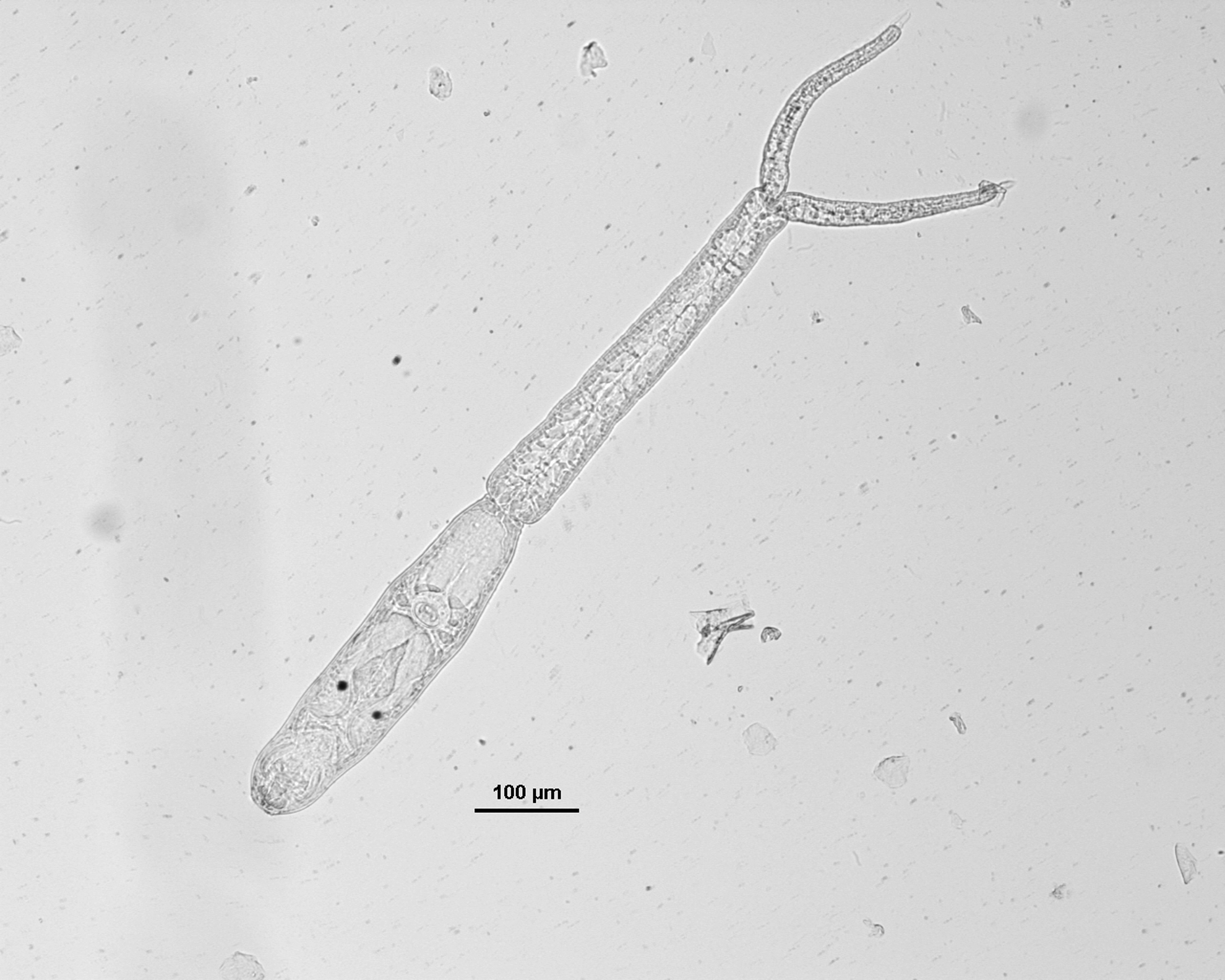
Ocellate furcocercariae.
Molecular analysis of positive snails and cercariae
Molecular analyses from 2015 outbreaks led to the amplification of DNA from shedding snails and DNA from ocellate furcocercariae. Two different ITS-2 sequences were obtained for the seven positive snails. Six were identical (335 bp) (Genbank: KX034091) and were found to be 100% identical to R. balthica (GenBank: HE573099). The last sequence (339 bp) (GenBank: KX034090) was found to be 100% identical to R. auricularia (GenBank: KT337585)]. Concerning DNA from the parasites, two different D2 sequences were obtained for the seven batches of ocellate furcocercariae. The six R. balthica snails shed T. franki cercariae and their D2 sequences (552 bp) (GenBank: KX034089) were found to be 100% identical to T. franki haplotype peregra (GenBank: EU413962). R. auricularia shed T. franki cercariae and its D2 sequence (552 bp) (GenBank: KX034088) was found to be 100% identical to T. franki haplotype auricularia (GenBank: KJ775866).
Discussion
The morphological identification of lymnaeids species in the genus Radix is very difficult because of the continuous variability and plasticity of the different criteria depending on environmental conditions; consequently, molecular tools are extensively used to overcome these difficulties (Bargues et al. 2001, Pfenninger et al. 2006). Thus, preadult of R. auricularia could be confused with R. balthica or R. labiata, for example. In our study, the snails shedding ocellate furcocercariae belong to two species: R. balthica ( = R. peregra = R. ovata) (Bargues and Mas-Coma 2005) and R. auricularia. No ocellate furcocercariae were shed by the collected L. stagnalis.
The morphology of cercariae of T. franki is very similar to T. regenti (Lawton et al. 2014) and morphometric parameters are unreliable for species identification of cercariae of Trichobilharzia species (Horak et al. 2002). Nevertheless, some morphological differences were previously noted (Jouet et al. 2010) between T. franki haplotype peregra and T. franki haplotype auricularia: the total size and extent of the head organ, tail, and furca may be an indication, since these measures differ between the two. Unfortunately, it was not possible to objectivize these differences in this study. A molecular approach was therefore considered essential for accurate identification.
Interestingly, this approach permitted to differentiate two different haplotypes of T. franki as previously recorded in France (Jouet et al. 2010). This difference is based on three nucleotide variations and the fact that the D2 domain must be conserved and specific tend to ascribe this as an interspecific rather than an intraspecific variation. Furthermore, this difference was also detected within other domains (e.g., 5.8S rDNA or COX1 of mDNA) (Jouet et al. 2010). The close relationship between the species sampled in this study and populations found throughout France is probably linked to the migratory pattern of their definitive Anseriforme hosts (duck and swans) (Lawton et al. 2014).
This is the first documented report of T. franki in Belgium. Two reasons may explain this epidemiological gap. First, it was difficult to detect the presence of ocellate furcocercariae during this study. This was probably due to the fact that prevalences in the snails are generally low (between 0.05% and 5.0%) (Soldanova et al. 2013). In 2015, sampling was performed in different places around the lake, including the bathing areas, and this probably increased the probability to detect the responsible parasite and its snail intermediate hosts. Indeed, the positive samples in this study were collected from a known nesting and wintering area of Anas platyrhynchos, a well-known definitive host of T. franki (Horak and Kolarova 2011). Moreover, wind or activity of Plate Taille dam might create water currents responsible for the dispersion of cercariae as it was shown elsewhere (Leighton et al. 2000).
Presumptive HCD were recorded in summer 2011 and 2012 in the Boekenbergpark, Belgium (51°11′54.28″ N; 4°27′48.77″ E, 10 m a. s. l.), by a private company (Spygen®) (T. de Winter, personal communication). In September and October 2012, two presumptive HCD cases occurred in the lake of Blaarmeersen, Belgium (51°02′38.38″ N; 3°41′07.05″ E, 4 m a. s. l.), but no ocellate furcocercariae were recorded following light exposure (J. Charlier, personal communication).
The number of disease outbreaks in any summer is strongly linked to the number of days with a maximum temperature equal or above 25°C (Schets and de Roda Husman 2014). Global warming (with mild winters and hot summers) could increase the number of cases because all fluke infections in snails are climate/temperature sensitive with higher ambient temperatures tending to promote higher cercarial emission, but also higher snail numbers increasing the potential number of intermediate hosts and thus the risk of infection (Mas-Coma et al. 2009).
High eutrophication of surface water bodies linked to human activities was also considered a cause of increase of HCD (de Gentile et al. 1996, Schets et al. 2008, Soldanova et al. 2013). However, the parasite presence seems not linked to water quality as it was stated in Annecy Lake, which is an important drinking water reservoir, and Plate Taille Lake, classified as oligomesotrophic. Climate changes can also affect the distribution of definitive hosts, by altering migratory routes of waterfowl (Horak and Kolarova 2011).
It is worth mentioning that, all HCD are not always properly diagnosed by the medical professionals (symptoms can be confused with other allergic reactions or insect bites) and/or widely publicized in the local and national press. The creation of a center for the report of presumptive HCD cases could be useful to assess the real impact of the disease and set up preventive measures.
Footnotes
Acknowledgments
The authors would like to thank Elodie Renard and Caroline Vanvinckenroye for the help during sampling, Philippe Deflorenne for information concerning birds, and the interdisciplinary Group of Applied Genoproteomics (GIGA) Genomic Facility (
Author Disclosure statement
No competing financial interests exist.