
Abstract
Bartonellae are blood-borne and vector-transmitted pathogens, some are zoonotic, which have been reported in several Mediterranean countries. Transmission from dogs to humans is suspected, but has not been clearly demonstrated. Our objectives were to determine the seroprevalence of Bartonella henselae, Bartonella vinsonii subsp. berkhoffii, Bartonella clarridgeiae, and Bartonella bovis (as a proxy for Candidatus Bartonella merieuxii) in stray dogs from Tunisia, identify the Bartonella species infecting the dogs and evaluate potential risk factors for canine infection. Blood samples were collected between January and November 2013 from 149 dogs in 10 Tunisian governorates covering several climatic zones. Dog-specific and geographic variables were analyzed as potential risk factors for Bartonella spp. seropositivity and PCR-positivity. DNA was extracted from the blood of all dogs and tested by PCR for Bartonella, targeting the ftsZ and rpoB genes. Partial sequencing was performed on PCR-positive dogs. Twenty-nine dogs (19.5%, 95% confidence interval: 14–27.4) were seropositive for one or more Bartonella species, including 17 (11.4%) for B. vinsonii subsp. berkhoffii, 14 (9.4%) for B. henselae, 13 (8.4%) for B. clarridgeiae, and 7 (4.7%) for B. bovis. Statistical analysis revealed a few potential risk factors, mainly dog's age and breed, latitude and average winter temperature. Twenty-two (14.8%) dogs, including 8 of the 29 seropositive dogs, were PCR-positive for Bartonella based on the ftsZ gene, with 18 (81.8%) of these 22 dogs also positive for the rpoB gene. Partial sequencing showed that all PCR-positive dogs were infected with Candidatus B. merieuxii. Dogs from arid regions and regions with cold average winter temperatures were less likely to be PCR-positive than dogs from other climatic zones. The widespread presence of Bartonella spp. infection in Tunisian dogs suggests a role for stray dogs as potential reservoirs of Bartonella species in Tunisia.
Background
B
Limited studies have been conducted on canine bartonellosis in North Africa. A serologic study in Morocco found that stray dogs were more likely than urban pet dogs to be seropositive for B. vinsonii subsp. berkhoffii (Henn et al. 2006). In Algeria, B. vinsonii subsp. berkhoffii, B. clarridgeiae, B. henselae, and B. rochalimae have been detected by molecular testing in Bartonella-infected dogs (Kernif et al. 2010, Azzag et al. 2015). A limited study on Bartonella seroprevalence in domestic dogs in northern Tunisia reported an overall seroprevalence of 23.5% (Makhlouf 2008).
Tunisia is a geographically diverse country divided into 24 governorates. Recent studies have shown that age and climate are potential risk factors for vector-borne infections in dogs (Gubler et al. 2001, Gage et al. 2008). Knowing the seroprevalence of Bartonella spp. in dogs, the Bartonella species infecting these dogs and the risk factors for canine infection would help us to better understand potential transmission mechanisms for zoonotic bartonellosis in humans and the potential health impact on these dogs.
The goals of this study were to determine the seroprevalence of four Bartonella species or subspecies in stray dogs in Tunisia and the Bartonella species infecting these dogs, based on PCR analysis of ftsZ and rpoB genes. We also aimed at analyzing several dog-specific and climatic/geographic variables as potential risk factors for Bartonella seropositivity, and risk factors for PCR-positive status. The results of this study will expand our understanding of dogs as reservoirs and potential vectors of zoonotic Bartonella species in North Africa.
Materials and Methods
Study design, animals, and sample collection
Stray dogs captured as part of the National Program for Rabies Control (NPRC) in Tunisia were enrolled in the study. Blood samples were collected between January and November 2013 in collaboration with veterinarians from the Institut National de Recherche Vétérinaire de Tunis (IRVT). On average, 6000 stray dogs are vaccinated against rabies every year under the NPRC in 21 of the 24 Tunisian governorates. For this study, 149 stray dogs (a convenience sample consisting of 20% of dogs sampled by NPRC veterinarians) from 10 Tunisian governorates (Bizerte, Nabeul, Zaghouan, Sousse, Monastir, Kairouan, Kasserine, Sfax, Beja, Jendouba), covering four different climatic zones were selected (Fig. 1). These governorates were chosen because they have different geographic characteristics that affect their climate. Climate in these regions was classified as humid, sub-humid, semi-arid, or arid, based on average yearly rainfall (Fig. 1). Average winter temperatures for each region, as recorded by the Tunisian National Weather Office, were categorized as cold (<10°C), moderate (10–20°C), or warm (≥21°C).
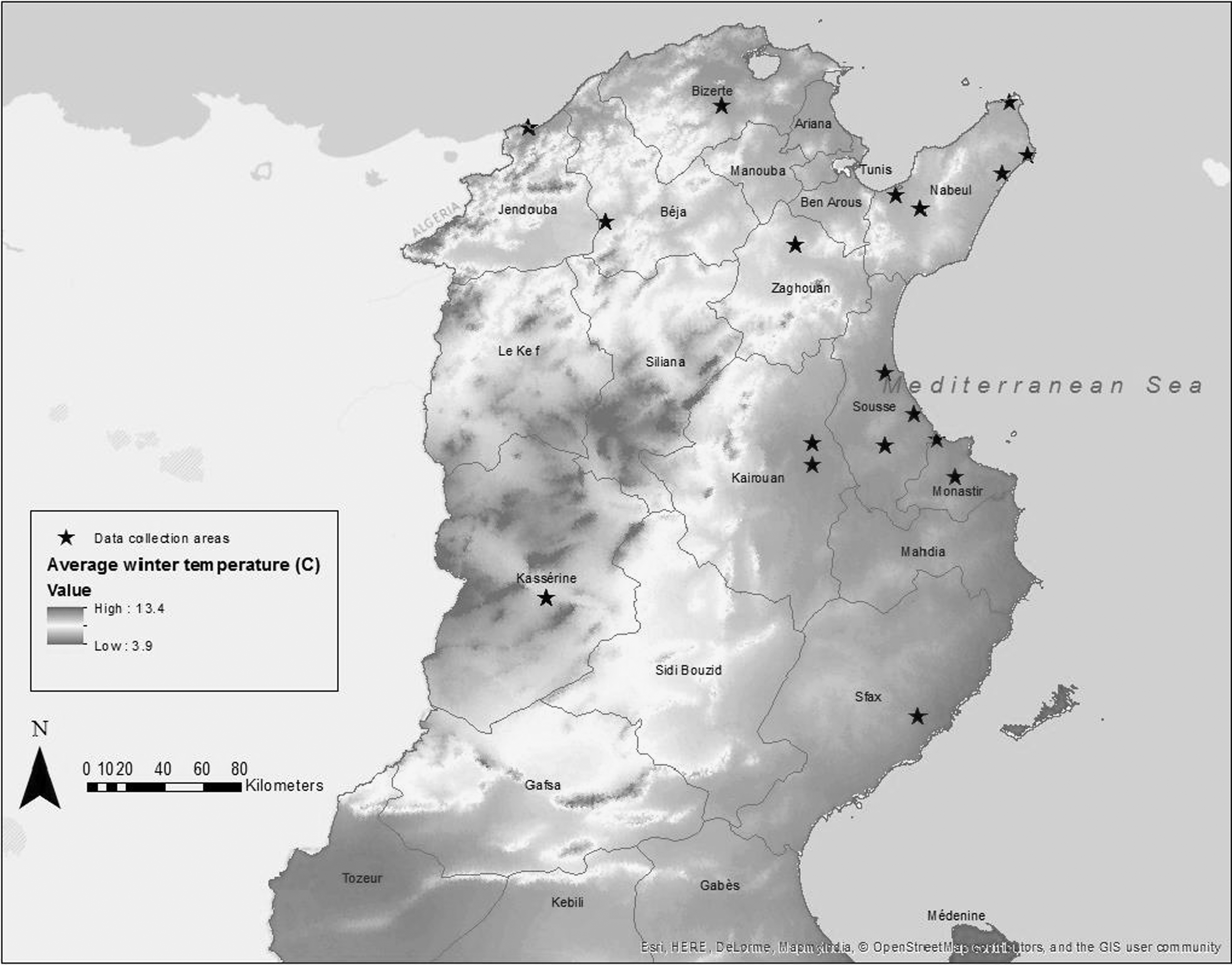
Political and administrative regions (governorates) of Tunisia. Stars indicate locations of dog's blood sample collection. Average winter temperatures (in °C) in Tunisia. (Map designed with ESRI-ArcGIS Desktop: Release 10.3, Redlands, CA: Environmental Systems Research Institute)
Estimated age in months, based on tooth wear, sex, breed, presence of external parasites (fleas and/or ticks), and location of sample collection were recorded and analyzed as potential risk factors. A visual inspection for ectoparasites (up to 2 min/dog) was performed by the IRVT veterinarians. Five to 10 mL of whole blood was collected from the jugular or saphenous vein. Approximately 40 μL of blood was placed onto a Nobuto filter paper strip (AdvantecH Type I Blood Sampling Paper; Toyo Roshi Taishu, Ltd., Tokyo, Japan) via syringe and air-dried, as described previously (Chomel et al. 2004), because of importation constraints. Strips were placed into an envelope and stored at room temperature (20–25°C) for 2 months until shipped to the School of Veterinary Medicine, University of California, Davis (UCD) for serologic and molecular analysis. The remaining blood was transferred to an EDTA tube and stored at −20°C at the IRVT for backup.
Serology
Using sterile scissors, one-third of each Nobuto strip was cut off and placed in a tube containing 400 μL of phosphate-buffered saline and 1 μL of thimerisol and stored overnight at 4°C. An indirect immunofluorescence assay was used to detect antibodies against B. henselae H1 (ATCC 49882) and U4 (UC Davis), B. clarridgeiae (ATCC 51734), B. vinsonii subsp. berkhoffii genotype I (ATCC 51672), and Bartonella bovis (strain 91-4T), as a proxy for Candidatus B. merieuxii, as previously described (Bermond et al. 2002, Brenner et al. 2013). For initial screening, extracted blood from Nobutos was diluted with milk buffer at 1:8 (corresponding to a 1:64 serum dilution), as previously reported (Yamamoto et al. 1998) and 20 μL of the dilution was added to test wells. Positive and negative controls were included on each slide. Three conjugate dilutions (Cappel fluorescein-conjugate goat anti-dog IgG fraction; MP Biomedicals, Aurora, OH) were used for the four antigens: 1:1400 (B. vinsonii subsp. berkhoffii and B. bovis), 1:2800 (B. henselae H1 and U4) and 1:3600 (B. clarridgeiae), as previously described (Brenner et al. 2013). For each well, 20 μL of conjugate was added to the respective slides. The cutoff for a positive test result was fixed at grade ≥2 on a scale of 4 at 1:64 dilution. Any sample positive at 1:64 dilution was tested to final titer point up to 1:1024 (Table 1). Three independent readers graded the intensity of the fluorescence and the mean of the three values was determined.
DNA extraction
Using sterile scissors, another one-third of each Nobuto strip was cut off and placed in a tube containing 180 μL ATL buffer (QIAGEN, Valencia, CA). The vial was incubated at 90°C for 15 min; 20 μL of proteinase K solution (QIAGEN) was then added and the vial was incubated at 56°C for 1 h. Extraction was continued using the manufacturer's (QIAGEN) recommended procedure. Final elution volume was 100 μL in AE buffer heated to 70°C (QIAGEN).
PCR testing
PCR reactions were carried out in a total volume of 50 μL, containing 0.5 μM primers, 200 μM of dNTP, 1.25 U of AmpliTaq Gold DNA Polymerase (Applied Biosystems, Foster City, CA), and 5 μL of template DNA or water (negative control). DNA from B. henselae strain U4 was used as a positive Bartonella control. For the PCR of the ftsZ gene fragment primers prAPT0257, forward GCCTTCAAGGAGTTGATTTTGTTGTTGCCAAT and prAPT0258, reverse ACGACCCATTTCATGCATAACAGAAC were used (Veikkolainen et al. 2014). PCR run conditions were initial denaturation at 95°C for 5 min, followed by denaturation at 95°C for 30 s, annealing at 55°C for 40 s, and extension at 72°C for 1 min. Amplification was completed by 40 additional cycles and final extension at 72°C for 10 min.
For the PCR of the rpoB gene fragment, primers prAPT0244, forward GATGTGCATCCTACGCATTATGG and prAPT0245, reverse AATGGTGCCTCAGCACG TATAAG were used (Oksi et al. 2013). PCR run conditions were an initial denaturation at 95°C for 5 min, followed by denaturation at 95°C for 30 s, annealing at 58°C for 1 min, and extension at 72°C for 1 min. Amplification was completed by 45 additional cycles and final extension at 72°C for 10 min. PCR gene fragments were separated in 2% agarose gel and visualized by staining with GelRed (Phenix Research Products, Candler, NC) and a BioRad Gel Doc EZ Imager (Hercules, CA).
Sequencing and phylogenetic analysis
Amplified PCR products were cleaned using a Qiagen QIAquick PCR Purification Kit according to the manufacturer's instructions. Purified PCR products (suspect bands) were sequenced at the College of Biological Sciences DNA Sequencing Facility at the University of California, Davis. Phylogenetic trees were constructed from ftsZ and rpoB sequences using the MrBayes plugin in Geneious version 8.1.7 (
Statistical analysis
Seroprevalence was determined for each of the four Bartonella species by dividing the number of seropositive dogs by the total number of dogs sampled. Approximated confidence intervals (CIs) for the seroprevalence were calculated using the package “binom.” All statistical analysis for risk factors associated with seropositivity was performed using R software version 0.98.501 (
Results
Of the 149 dogs, 91 (61.1%) were male, 58 (38.9%) were female, 126 (84.6%) were from mixed breed and 23 (15.4%) were pure bred, including German Shorthair Pointer (4), English Pointer (4), Labrador retriever (3), German Shepherd (3), Pit bull Terrier (3), American Staffordshire Terrier (2), Cocker Spaniel (2), Rottweiler (1), and Poodle (1). Age ranged from 4 months to 12 years, with 26 (17.7%) dogs <1 year, 69 (46.9%) 1 to 2 years, 34 (23%) 3 to 4 years and 18 (12.2%) ≥5 years. Age was not recorded for two dogs, including one seropositive dog. Fleas and ticks were observed on 16 (10.7%) and 38 (25.6%) dogs, respectively. The distribution of dogs in the 10 Tunisian governorates was as follows: Nabeul (n = 29, 19.5%), Sousse (n = 20, 13.4%), Monastir (n = 17, 11.4%), Kasserine (n = 15, 10.1%), Zaghouan, Beja and Jendouba (n = 13, 8.7% each), Kairouan (n = 11, 7.4%) and Sfax and Bizerte (n = 9, 6% each). Based on these locations, dogs were from four climatic zones: humid (n = 13, 8.7%), semi-humid (n = 38, 25.5%), semi-arid (n = 63, 42.3%), and arid (n = 35, 23.5%) where average winter temperature was warm (n = 24, 16.1%), mild (n = 110, 73.8%), or cold (n = 15, 10.1%). Because of the limited time to adequately inspect dogs for presence of ectoparasites, it precluded any proper evaluation of ectoparasite infestation and Bartonella status.
Bartonella seroprevalence
Twenty-nine dogs (19.5%, 95% CI: 14.0–27.4) were seropositive for one or more Bartonella species, including 17 (11.4%, 95% CI: 8.3–19.9) seropositive for B. vinsonii subsp. berkhoffii, 14 (9.4%, 95% CI: 5.7–16) for B. henselae, 13 (8.4%, 95% CI: 3.7–12.8) for B. clarridgeiae and 7 (4.7%, 95% CI: 3.2–12) for B. bovis. Antibody titers ranged from 1:64 to 1:1024 (Table 1). Of the 29 seropositive dogs, 12 (41.4%) were positive for more than one antigen. Three (25%) of these last 12 were seropositive for all four antigens. All dogs seropositive for B. bovis were also seropositive for at least one other antigen (Table 1). Seroprevalence increased with age: 7.7% (n = 2) for dogs <1 year, 8.6% (n = 6) for dogs 1–2 years, 41.2% (n = 14) for dogs 3–4 years, and 35.3% (n = 6) for dogs ≥5 years. A Bartonella seroprevalence gradient was observed by climatic zones, ranging from 5.6% in the arid zone to 54% in the humid zone (Table 2).
Molecular analysis of strains
Twenty-two of the 149 dogs (14.8%), including 8 (27.6%) of the 29 seropositive dogs, and 14 (11.6%) of the 120 seronegative dogs were PCR-positive for Bartonella sp. based on the ftsZ gene with 18 (81.8%) of these 22 dogs also positive for the rpoB gene. Partial sequencing showed that all PCR-positive dogs were infected with Candidatus B. merieuxii. Three representative PCR-positive dogs (Dogs Nos. 18, 50 and 75) were included in the dendograms (Figs. 2 and 3). All partial sequences were deposited in GenBank (Table 3). Sequence analysis of the organism for both ftsZ and rpoB genes clustered with 100% homology (541/541 base pairs [bp] for ftsZ and 422/422 bp for rpoB) with a dog (Dog F059) and jackal strains (Jackal F040) from Iraq (Chomel et al. 2012) and a dog (HMD No. 14) from Greece (Mylonakis et al. 2014). It also had a 100% sequence similarity with the previously described HMD strain for both genes (Diniz et al. 2009). The next closest Bartonella species were Bartonella dromedarii (97.6% homology based on 549 bp) reported from camels from Israel (Rasis et al. 2014) and other ruminant Bartonella species (Bartonella capreoli with 95.93% homology for 550 bp; Bartonella chomelii with 95.75% homology based on 549 bp; and B. bovis and Bartonella schoenbuchensis with 95.56% homology based on 549 bp).
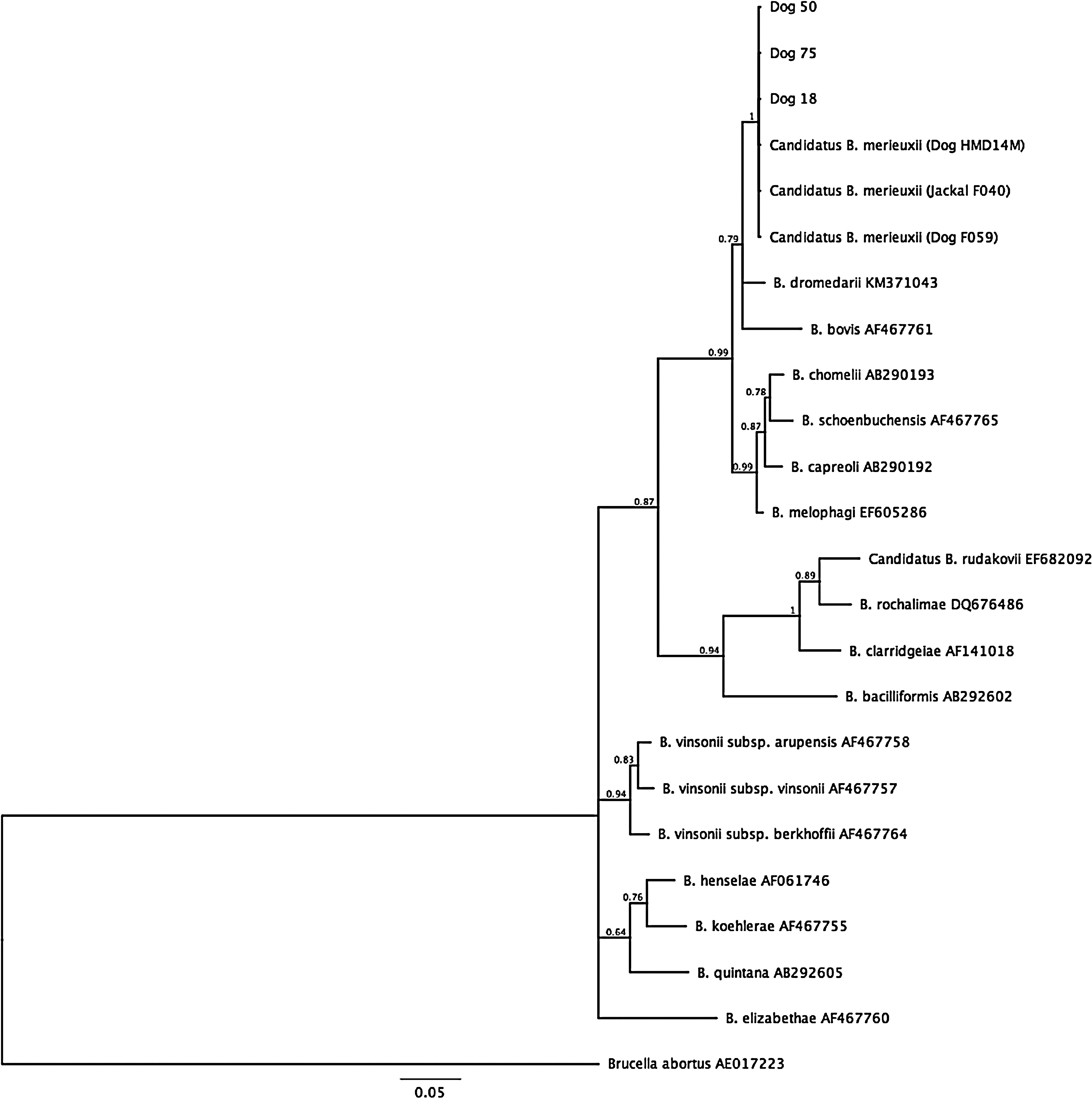
Dendogram of Bartonella strains identified by partial sequencing of the ftsZ gene. The phylogenetic tree was constructed from ftsZ sequences using the MrBayes plugin in Geneious version 8.1.7 with a 1,100,000 MCMC and 100,000 burn-in length.

Dendogram of Bartonella strains identified by concatenating partial sequencing of the rpoB and ftsZ genes. The phylogenetic tree was constructed from rpoB and ftsZ sequences using the MrBayes plugin in Geneious version 8.1.7 with a 1,100,000 MCMC and 100,000 burn-in length.
Risk factor analysis
Bivariate analysis for dog seropositivity to any of the four antigens tested led to the selection of the following variables as potential risk factors: age (p < 0.001), breed (p = 0.004), weather (p = 0.003), average winter temperature (p = 0.1), presence of external parasites (p = 0.005), latitude (p = 0.01), and longitude (p = 0.02). The best fitting model selected by backward elimination analysis included age (adults vs. young dogs, OR = 5.31, 95% CI: 2.07–13.61) and latitude (OR = 3.64, 95% CI: 1.43–9.26) for increased risk and pure breed (OR = 0.17, 95% CI: 0.02–1.41) and warm average winter temperature, warm vs. moderate (OR = 0.18, 95% CI: 0.03–0.91) as protective factors.
Dogs from the arid zone were significantly less likely to be PCR-positive than dogs from other climatic regions (p = 0.027, Fisher exact test), as prevalence of PCR-positive dogs ranged from 2.8% for dogs from the arid zone to 23.1% for dogs from the humid zone (Table 2). PCR-positive dogs were from regions with mild (81.8%, 18/22) or warm (16.7%, 4/22) average winter temperature. None of the dogs from areas where winters are cold were Bartonella PCR-positive. Prevalence of PCR-positivity was quite similar between males (15.4%, 14/91) and females (13.8%, 8/58). Fourteen (63.6%) of the 22 PCR-positive dogs were seronegative for the four Bartonella antigens used for serologic testing and two PCR-positive dogs were seropositive for all four antigens.
Discussion
We demonstrated that antibodies to one or more Bartonella spp., including two zoonotic pathogens, and presence of Bartonella DNA were common in dogs in specific climatic and geographic regions in Tunisia. Dog's age and breed, average winter temperature, and latitude were risk factors for Bartonella seropositivity. Several variables known to be risk factors for Bartonella seropositivity in previous studies, dogs being stray (usually more likely to be from mixed breeds) (Henn et al. 2006, Azzag et al. 2015) and dogs being of older age (Henn et al. 2007) were confirmed in this study. However, risk factors such as presence of external parasites (Henn et al. 2005), were not confirmed in the multivariate analysis, likely due to our small sample size.
Based on PCR analysis of ftsZ and rpoB genes, Candidatus B. merieuxii was detected in 15% of the dogs, including 11% of the seronegative ones. Our results confirm the presence of Bartonella sp. in Tunisia, identifying some possible risk factors for exposure and infection, and expand the known distribution of Candidatus B. merieuxii. Importantly, these findings support further investigation of the potential role of dogs as reservoirs for zoonotic transmission of Bartonella sp. in Tunisia.
The overall prevalence (20.1%) of Bartonella antibodies in stray dogs was similar to the 23.5% reported in a more limited study of owned dogs from northern Tunisia (Makhlouf, 2008), but was lower than that observed in dogs from Morocco (38%) and Algeria (27–32.4%) (Henn et al. 2006, Azzag et al. 2015). The highest seroprevalence (11.4%) and antibody titers were observed for B. vinsonii subsp. berkhoffii, suggesting that this Bartonella subspecies is more likely present in Tunisian dogs. Therefore, the lack of detection of PCR-positive dogs for this Bartonella subspecies is puzzling and requires further investigation on a larger sample of dog, as B. vinsonii subsp. berkhoffii was detected in dogs from Algiers, Algeria (Kernif et al. 2010, Azzag et al. 2015). However, this prevalence is in agreement with the seroprevalence of B. vinsonii subsp. berkhoffii in rural dogs in Turkey (12% of 333) (Celebi et al. 2010b) and in dogs suspected of having tick-borne disease in Israel (10% of 40 dogs) (Baneth et al. 1998). In southern Italy, 28% of 205 dogs tested were seropositive for B. henselae in Sassari, Sardinia (Pinnaparpaglia et al. 2007).
Several dogs were positive for more than one Bartonella antigen, including three that were seropositive for all four antigens tested. Because this was a cross-sectional study, it was difficult to determine whether this was the result of cross-reactivity, co-infection, or sequential infection by the various Bartonella species. Cross-reactivity between various Bartonella antigens has been reported in animals presenting with high Bartonella titers, especially in cases of endocarditis (Chomel et al. 2001), but appears unlikely in this study since almost 60% of seropositive dogs were positive for only one antigen. In addition, experimental infection of dogs with various strains of Bartonella led to seroconversion only to the specific inoculum (Balakrishnan et al. 2013). Our target population was free-roaming dogs from urban and rural areas of Tunisia, which may explain the high prevalence of seropositivity for multiple organisms at the time of blood collection. Stray dogs are more likely to be exposed to ectoparasites, and nearly one-third of dogs in our study had ticks or fleas.
Age was a primary risk factor for seropositivity in the Tunisian dogs. Older dogs were more likely to be seropositive than younger dogs, which is consistent with longer exposure of stray dogs to organisms in the environment over time. In free-roaming dogs from Northern California, antibody prevalence for Bartonella also increased with age (Henn et al. 2007). A similar trend has been described for other vector-borne diseases (ehrlichiosis and rickettsiosis) in dogs (Cocco et al. 2003). Sex has not been previously identified as a primary risk factor for Bartonella in animal reservoirs (Müller et al. 2004, Solano-Gallego et al. 2006, Foley et al. 2007).
Latitude was indicative of a higher risk of seroprevalence in dogs living in the northern part of the country, which is also the most humid region. Such data are in concordance with other prevalence studies in domestic dogs (Foley et al. 2007) and coyotes (Beldomenico et al. 2005) in California. In coyotes, it was shown that Bartonella seropositivity was associated with higher precipitation (p = 0.003) and proximity to the coast (p = 0.007). Climatic factors (warm and humid) have been reported as a risk factor for B. henselae in cats in North America (Jameson et al. 1995).
Our identification of Candidatus B. merieuxii in Tunisian dogs expands the known geographic distribution of this Bartonella species in dogs beyond Iraq (Chomel et al. 2012), Greece, and southern Italy (Diniz et al. 2009, Mylonakis et al. 2014). None of the Bartonella PCR-positive dogs were infected with B. henselae, B. rochalimae, B. clarridgeiae, or B. vinsonii subsp. berkhoffii, as reported for dogs from northern Algeria (Kernif et al. 2010, Azzag et al. 2015). Through molecular analysis, we detected Candidatus B. merieuxii in 14.8% of the dogs, with significantly lower prevalence in dogs in the arid and dry climatic zone, likely associated with a lower level of exposure to ectoparasites, especially fleas, which are more likely to thrive in more humid environments. In Iran, flea populations decrease dramatically as rainfall decreased and temperature increased (Tavassoli et al. 2010). Increased PCR prevalence with age could have been related to an increased risk of exposure to the pathogen over time. The high percentage of PCR-positive dogs that were Bartonella seronegative may also be due to the choice of antigens from species usually common in dogs. As Candidatus B. merieuxii is closely related to Bartonella species isolated from ruminants, a lack of cross-reactivity with the antigens we used may be a plausible explanation for the high percentage of seronegative animals among the ones that were PCR-positive. Positivity to other antigens may also be due to earlier exposure of these dogs to different species of Bartonella, as our study was cross-sectional. The small sample size and limited information on presence of ectoparasites did not allow determining the likelihood of a specific vector; although there was a higher prevalence of PCR-positive dogs among those infested by fleas versus ticks. A recent study from Algeria, reported the detection of Bartonella from fleas collected on dogs and cats (Bessas et al. 2016). Almost 10% (21/225) of the tested fleas were PCR-positive with B. henselae detected in almost 5% (11/225) of the fleas, including 11 of 87 Ctenocephalides felis collected from cats and the 10 other PCR-positive fleas having sequences matching B. vinsonii subsp. berkhoffii in 6 of 62 Xenopsylla cheopis fleas collected on dogs and B. clarridgeiae in four of the 87 C. felis collected on cats.
A limitation of this study was the convenience sampling method, which may have introduced a selection bias, and the limited number of Bartonella-positive dogs. Similarly, lack of real estimates of the dog population by governorate and not including dogs from southern Tunisia may have biased the overall prevalence estimation for the country. Due to political instability, we were not able to sample dogs in some parts of Tunisia. Despite these limitations, our study supports the wide distribution of Bartonella among free-roaming Tunisian dogs.
It will be important to evaluate the respective role of stray and pet dogs in the transmission of Bartonella species to humans of these zoonotic bacteria. Beside the high prevalence of B. quintana in humans in Tunisia (Znazen et al. 2005) and Candidatus B. merieuxii in stray dogs, it will be important to determine the presence of B. henselae and other zoonotic Bartonella, such as B. vinsonii subsp. berkhoffii or B. rochalimae, as reported in Algeria (Kernif et al. 2010, Azzag et al. 2015).
Conclusions
Candidatus B. merieuxii was confirmed by PCR analysis in 15% of the dogs, expanding the known geographic distribution of this Bartonella species. Antibodies to various Bartonella species, including two with zoonotic potential were detected in stray dogs from diverse climatic and geographic regions of Tunisia. Latitude and average winter temperature were identified as risk factors for Bartonella seropositivity, as well as dog's age and breed. However, our results will need to be strengthened with a larger national study that includes a randomly selected population of dogs and ectoparasites from all regions of Tunisia, especially the southern deserts, to provide a better understanding of the distribution and prevalence of Bartonella infection across Tunisia and its zoonotic impact.
Footnotes
Acknowledgments
The authors thank the veterinarians and technicians from the National Program for Rabies Control in Tunisia, the Institut National de Recherche Vétérinaire de Tunis, and local veterinarians involved in blood sample collection.
Authors' Contributions
Samples were collected by T.B.H. Serology results were obtained by J.B. and D.F. Molecular testing was performed by J.B. and R.K. Sequence analysis was performed by R.K. and M.S. The project was designed and article written by the combined efforts of J.B., B.C., M.C., and T.F. Statistical analysis was performed by J.B. and T.B. Support and technology was provided by D.F. and M.S. All authors read and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.