
Abstract
Little is known of the endemicity of flea-borne rickettsiae in Kazakhstan. Thus, a survey for rickettsiae within great gerbil fleas was conducted in Almaty oblast. High prevalence of Rickettsia asembonensis was detected among Xenopsylla gerbilli, demonstrating that flea-borne rickettsiae are endemic to southeastern Kazakhstan. Interestingly, no Rickettsia typhi were detected in these same fleas.
Introduction
F
Flea-borne diseases in Kazakhstan have been a significant health risk to inhabitants and visitors for ages, particularly plague (Davis et al. 2004, Atshabar et al. 2014). The major natural vector hosts in Kazakhstan for Yersinia pestis (plague) are the fleas that parasitize the great gerbil (Rhombomys opimus) (Begon et al. 2006, Wilschut et al. 2015). However, flea-borne rickettsial disease threats are little known in Kazakhstan; we, therefore, initiated a study to detect and identify rickettsiae within fleas collected from great gerbils to ascertain the presence and prevalence of flea-borne rickettsiae in Almaty oblast, in southeastern Kazakhstan.
Materials and Methods
Flea collection and identification
Members of the Taldykorgan anti-plague station collected 2,963 fleas in Almaty oblast (province) during 2014 from 5 rayons (districts). Individual fleas were collected from Balkhashskiy (n = 359), Karatalskiy (n = 313), Aksuskiy (n = 1,896), Iliyskiy (n = 105), and Uygurskiy (n = 290) rayons. Fleas collected from great gerbil burrows (238 fleas in 18 pools) were only obtained from Aksuskiy. Fleas were identified to species level by using a morphologic key (Ioffe et al. 1965). The 2,963 fleas of the same species were pooled together by individual host or host burrow (n = 247; mean: 12; range: 1–99 fleas per pool) and stored at or below −20°C. Distribution by rayon of the 247 pool samples containing the 2,963 fleas was as follows: Balkhashskiy (n = 55), Karatalskiy (n = 33), Aksuskiy (n = 82), Iliyskiy (n = 28), and Uygurskiy (n = 49). Collection data were recorded electronically and included: locations from where the fleas were collected (GPS coordinates), flea species, number of pools, number of fleas, and host species or host burrow. The fleas were subsequently delivered to the Kazakhstan Scientific Center for Quarantine and Zoonotic Disease (KSCQZD), Almaty, for molecular analysis. Mapping of the R. felis and RFLOs positive fleas was performed by utilizing ArcGIS (Esri, Redlands, CA).
Rickettsia detection and identification
DNA was extracted from pooled flea triturates by PrepMen Ultra Sample Preparation Reagent (Applied Biosystems, Foster City, CA) according to the manufacturer's instructions and stored at −20°C. Rickettsia genus-, R. felis genogroup-, and species-specific quantitative real-time PCR (qPCR) assays were used to detect and identify various flea-borne rickettsiae as previously described (Luce-Fedrow et al. 2015b). Briefly, initial screen testing of the 247 pooled flea DNA preparations was performed with the genus-specific Rick17b qPCR assay. The Rick17b qPCR assay-positive samples were subsequently tested by the R. typhi species-specific Rtyph and the R. felis genogroup-specific RfelG qPCR assays. Positive DNA samples for R. felis genogroup were then tested with the R. felis and R. asembonensis species-specific qPCR assays RfelB and Rasemb, respectively. During the writing of this article, data have been presented, which indicate that the RfelB qPCR assay detects Ca. R. senegalensis, in addition to R. felis (Maina et al. 2016a).
Statistics
The estimated prevalence (ep) of rickettsia-infected fleas by testing pooled samples was determined by generalized linear modeling, calculated by using maximum-likelihood estimates (EpiTools;
Results
Of the 2,963 fleas surveyed, all were collected from great gerbils (R. opimus) or their burrows in five rayons (districts) within Almaty oblast (province) in southeastern Kazakhstan. The flea species collected were identified to be: Coptopsylla lamellifer (374 fleas; 40 pools), Echidnophaga oschanini (3 fleas; 1 pool), Nosopsyllus laeviceps (151 fleas; 14 pools), Nosopsyllus tersus (5 fleas; 1 pool), Nosopsyllus turkmenikus (10 fleas; 1 pool), Paradoxophsyllus teretifrons (73 fleas; 2 pools), Xenopsylla conformis (17 fleas; 1 pool), Xenopsylla gerbilli (532 fleas; 87 pools), Xenopsylla hirtipes (354 fleas; 27 pools), and Xenopsylla skrjabini (1,444 fleas; 73 pools). Their distribution within the five rayons is shown in Table 1.
Name of Rayon (district) of Almaty oblast (province) in southeastern Kazakhstan.
Number of flea pools (number of individual fleas).
Extracted DNA from the flea pools assessed by genus-, R. felis genogroup-, and species-specific qPCR assays identified various Rickettsia species DNA (Table 2). Of the 247 flea pools, 88 (35.6%) were positive for Rickettsia spp., with an ep of 3.4 (95% CI of 2.7–4.2) per 100 fleas. Of these Rickettsia-positive samples, 48 (54.6%) were positive for R. felis genogroup and R. typhi DNA was not detected. From the 48 R. felis genogroup-positive samples, 32 (66.7%) were positive for R. asembonensis, 3 (6.3%) were positive for R. felis/Ca. R. senegalensis, 5 (10.4%) were positive for both R. asembonensis and R. felis/Ca. R. senegalensis, and 8 (16.7%) were identified as neither R. asembonensis nor R. felis/Ca. R. senegalensis. The collection sites with GPS coordinates for the R. felis and RFLOs positive fleas are listed in Table 3 and presented in Figure 1.
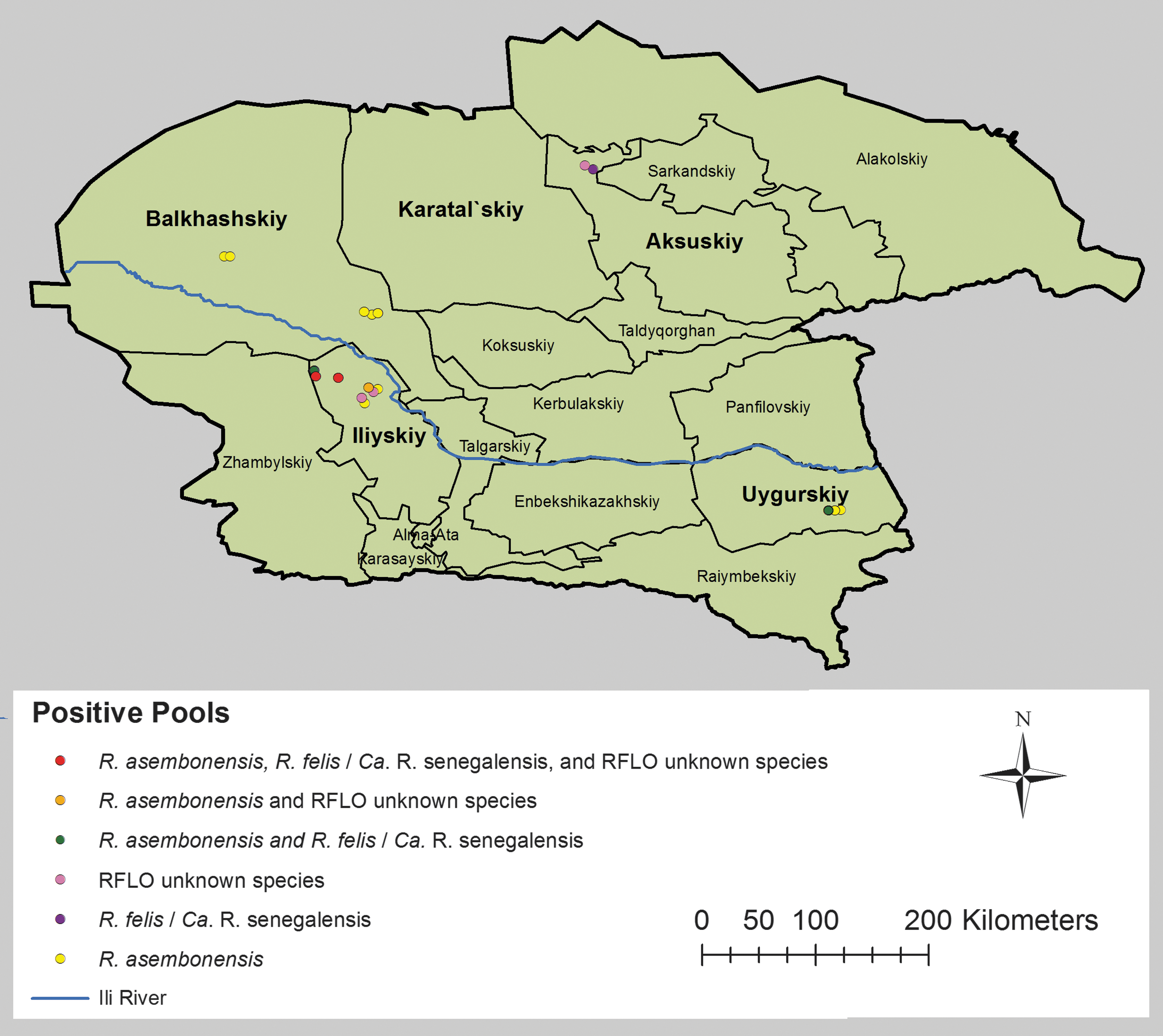
Rickettsia felis and RFLOs among fleas captured from great gerbils in Almaty oblast, southeastern Kazakhstan. Each dot represents a location where one or more flea(s) was/were collected. Red dots: sites of Rickettsia asembonensis, R. felis/Ca. R. senegalensis, and RFLO-unknown species positive flea pools. Orange dots: sites of R. asembonensis and RFLO-unknown species positive flea pools. Green dots: sites of R. asembonensis and R. felis/Ca. R. senegalensis positive flea pools. Pink dots: RFLO-unknown species. Purple dots: R. felis/Ca. R. senegalensis positive flea pools. Yellow dots: sites of R. asembonensis positive flea pools. RFLOs, Rickettsia felis-like organisms.
Rayon, an administrative district within an oblast (province).
Rickettsia typhi was not detected in any flea pool samples by Rtyph qPCR assay.
ep, 95% CI per 100 fleas.
Includes four pooled samples positive for both R. asembonensis and R. felis/Ca. R. senegalensis.
Includes one pooled C. lamellifer sample positive for both R. asembonensis and R. felis/Ca. R. senegalensis.
CI, confidence interval; ep, estimated prevalence; qPCR, quantitative real-time PCR; RFLOs, Rickettsia felis-like organisms.
The R. asembonensis positive flea pool samples, which include those pooled samples that were positive for both R. asembonensis and R. felis/Ca. R. senegalensis (n = 37 pools), were identified from 3 of the 5 rayons surveyed: Balkhashskiy (13 positive pools), Iliyskiy (9 positive pools), and Uygurskiy (15 positive pools). The Balkhashskiy positive samples comprised C. lamellifer (2 pools), X. gerbilli (10 pools), and X. hirtipes (1 pool) fleas. Flea samples from Uygurskiy that were positive for R. asembonensis consisted of C. lamellifer (3 pools), N. laeviceps (1 pool), and X. gerbilli (10 pools), and 1 pooled C. lamellifer sample positive for both R. asembonensis and R. felis/Ca. R. senegalensis. Five and four pooled X. gerbilli samples from Iliyskiy were positive for R. asembonensis and for both R. asembonensis and R. felis/Ca. R. senegalensis, respectively.
X. gerbilli was the flea species most commonly found in this survey to harbor R. asembonensis: 29 of 87 pools (33.3%) positive with an ep of 6.2 with 95% CI of 4.3–8.7 (p < 0.05.). C. lamellifer was the next most common flea species with 6 of 40 pools (15%) positive with an ep of 1.7 (95% CI of 0.7–3.3). The only other flea species positive for R. asembonensis DNA were single pools of X. hirtipes (1 of 27 pools [3.7%] positive with an ep of 0.3 with 95% CI of 0–1.2) and N. laeviceps (1 of 14 pools [7.1%] positive with an ep of 0.7 with 95% CI of 0–2.9).
R. felis/Ca. R. senegalensis positive flea samples (n = 8 pools) were identified from three of the five rayons surveyed: Aksuskiy (n = 1), Uygurskiy (n = 1), and Iliyskiy (n = 6). One X. skrjabini flea sample from Aksuskiy and one C. lamellifer flea pool sample from Uygur were positive for R. felis/Ca. R. senegalensis. Iliyskiy positive samples comprised X. gerbilli fleas (two pools positive for R. felis/Ca. R. senegalensis and four pools positive for both R. asembonensis and R. felis/Ca. R. senegalensis).
Eight RFLO samples that were not identified as either R. asembonensis or R. felis/Ca. R. senegalensis were detected in pool samples of X. hirtipes from Aksuskiy (n = 1), and X. gerbilli (n = 2), C. lamellifer (n = 1), and X. gerbilli (n = 4) from Iliyskiy. Sequence analysis was not performed to identify these RFLO agents.
Balkhashskiy and Iliyskiy had more rickettsia-positive samples than the other three rayons (p < 0.05); more R. asembonensis positive samples were from Balkhashskiy, Iliyskiy and Uygurskiy (p < 0.05), and R. felis/Ca. R. senegalensis; and other RFLO were found mostly from Iliyskiy (p < 0.05).
Fleas from burrows (n = 238) were only collected from one site, Aksuskiy rayon. The fleas collected included: C. lamellifer (1 pool; 5 fleas), X. conformis (1 pool; 17 fleas), X. hirtipes (2 pools; 35 fleas), and X. skrjabini (14 pools; 181 fleas). All of these 18 pooled flea samples were negative for rickettsial DNA. This finding maybe due to the site selected for assessing fleas captured from R. opimus burrows, Aksuskiy, which had a very low prevalence of rickettsia-infected fleas. Of 82 pools assessed from great gerbils from Aksuskiy, only 9 pools were Rickettsia positive, of which 2 were R. felis genogroup and R. felis positive (X. hirtipes [19 fleas/pool] and X. skrjabini [58 fleas/pool]). Moreover, the most commonly Rickettsia-infected flea collected during this survey, X. gerbelli, was not identified in the fleas obtained from Aksuskiy. Thus, there may have been a selection bias for assessing the utility of testing fleas from burrows for evidence of rickettsiae.
Discussion
R. felis, R. asembonensis, Ca. R. senegalensis, and other RFLOs belong to the antigenic spotted fever group rickettsiae (SFGR) and the genetic transitional group rickettsiae (TRGR; Gillespie et al. 2007). To date, only R. felis has been associated with flea-borne spotted fever, a febrile disease described around the world (Parola 2011); whereas the majority of RFLOs have only been detected in various arthropods (Odhiambo et al. 2014). However, three recent reports suggest that RFLOs can cause infections in vertebrates, since they have been detected in the blood of a dog (Kolo et al. 2016), monkeys (Tae et al. 2015), and a febrile patient from Senegal (Socolovschi et al. 2012). Rickettsial DNA was found in the blood nucleic acid preparations of a healthy dog in South Africa and in 12 out of 50 cynomolgus monkeys tested in Malaysia, which was the most similar to RF2125 and R. asembonensis, suggesting that this agent(s) can cause infection in mammals (Tae et al. 2015, Kolo et al. 2016). This agent has yet to be reported to cause infection in humans. However, there is an RFLO that was detected in the blood of a febrile patient from Senegal that was genetically related to Rickettsia sp. RF31 (Socolovschi et al. 2012). Thus, the pathogenicity of the RFLOs is still unresolved, though these agents should be of concern, as they are commonly found in fleas that bite humans.
The most extensively characterized RFLO by molecular methods is R. asembonensis (Maina et al. 2016b, Rzotkiewicz et al. 2015). This agent, initially identified in Asembo, Kenya (Jiang et al. 2013), has been reported elsewhere in Africa (Parola 2011), South America (Ecuador: Oteo et al. 2014, Columbia: Faccini-Martinez et al. 2016, and Peru: Kocher et al. 2016), North America (California: Maina et al. 2016a), Thailand (Khon Kaen: Odhiambo et al. 2014), and Israel (Rzotkiewicz et al. 2015). Related RFLOs that lack complete molecular characterization but appear to be, or are closely related to, R. asembonensis include: RF2125 from Thailand (Parola et al. 2003), Cf1 and Cf5 from United States, (Reeves et al. 2005), SE313 from Egypt (Loftis et al. 2006), Hu9-2007 from Hungary (Hornok et al. 2010), and Uruguay (Venzal et al. 2006). Thus, R. asembonensis or agents closely related to it have a large geographical footprint and have been associated with various flea species (Archaeopsylla erinacei, Ctenocephalides felis, Ctenocephalides canis, Echidnophaga gallinacea, Pulex irritans, Synosternus cleopatrae, Synosternus pallidus, Xenopsylla cheopis, Xenopsylla ramesis) (Gilles et al. 2009, Roucher et al. 2012, Jiang et al. 2013, Rzotkiewicz et al. 2015), tick species (Rhipicephalis sanguineus) (Kocher et al. 2016), mosquitoes (Socolovschi et al. 2012), and the tropical rat mite species: Ornithonyssus bacoti (Reeves et al. 2007). Thus, this agent is most likely a very primitive rickettsia that has not changed much during its adaptation to survival among various arthropod hosts (Rzotkiewicz et al. 2015).
Similar to R. asembonensis, Ca. R. senegalensis was initially identified in Africa (Senegal: Mediannikov et al. 2014) and has now been identified in Orange County, California (Maina et al. 2016a). The agent was also probably first identified in Thailand and reported as Rickettsia sp. RF31 (Parola et al. 2003) and later as Cf15 (GenBank acc. no. DQ379482), HL15c in Malaysia cat fleas (Tae et al. 2014), and, recently, in mosquitoes from Code d Ivoire and Gabon (Socolovschi et al. 2012). Thus, Ca. R. senegalensis has a distribution that is worldwide as well.
Other RFLOs not so well characterized but that appear to be genetically related to R. felis, R. asembonensis, and Ca. R. senegalensis include: SGL01 from tsetse flies (Glossina morsitans) (Medianniakov et al. 2012), Hf56 from hedgehog fleas (A. erinacei) (Gilles et al. 2009), and other arthropods, including mosquitoes (Socolovschi et al. 2012). The genetic relationship of these RFLOs is shown in the phylogram based on gltA sequences (Fig. 2). Although unable to directly show gltA sequence data from our study, the genetic relationship of the rickettsiae detected by the genus-, group-, and species-specific qPCR assays suggests that the RFLO from Almaty oblast, Kazakhstan, would group similarly to those shown in Figure 2.
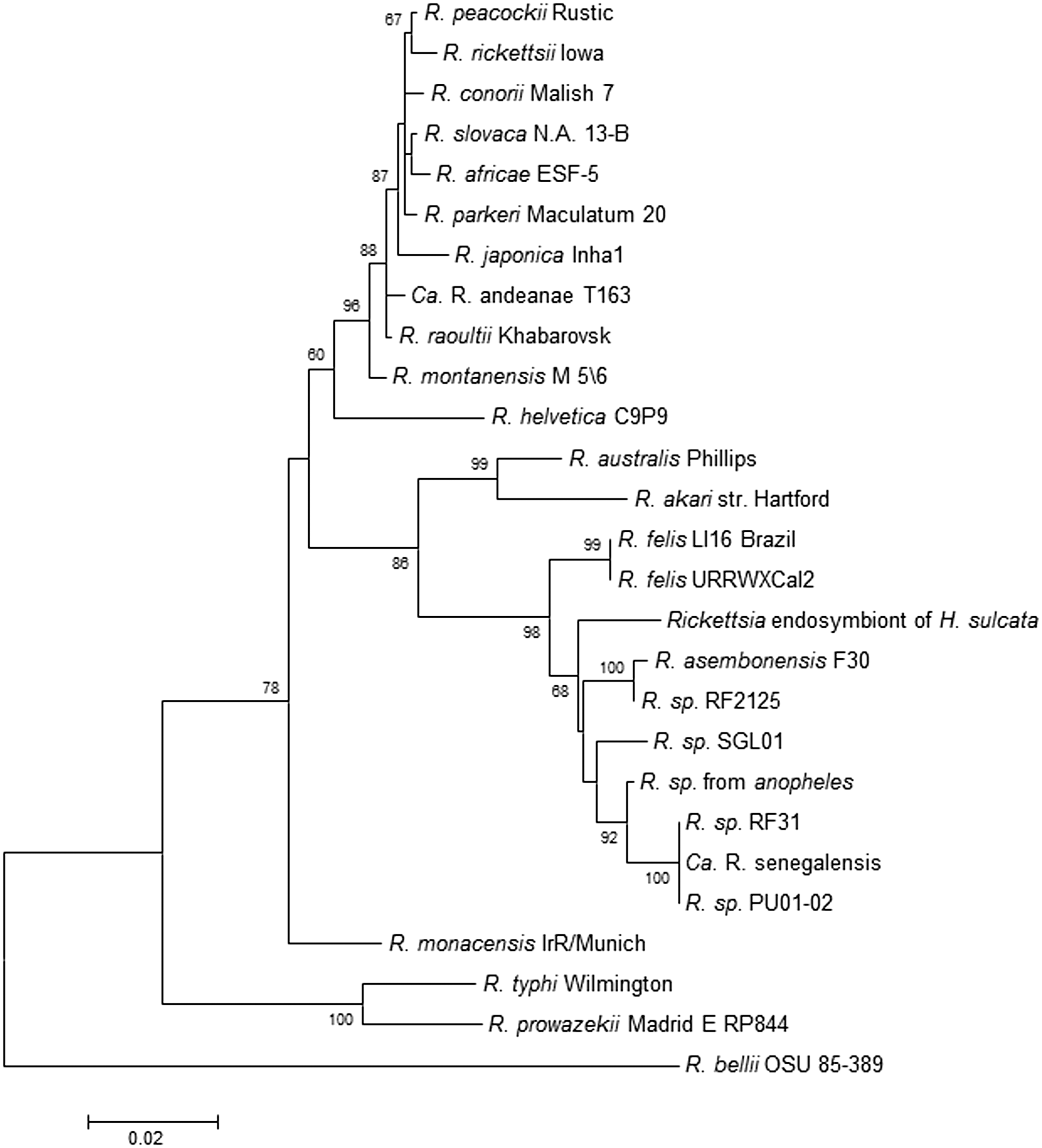
Rickettsia phylogenetic tree based on gltA sequences. The phylogenetic tree depicts the inferred evolutionary relationships of members of the genus Rickettsia including R. felis and RFLOs available in GenBank. There are no sequence data available from this study to include in this phylogenetic tree.
The test results described here demonstrated that there is a high prevalence of RFLO in fleas obtained from the great gerbil in Almaty oblast, southeastern Kazakhstan. These fleas (X. gerbilli, C. lamellifer, X. hirtipes, and N. laeviceps) have not been reported earlier to contain rickettsiae. Especially interesting is the high prevalence of rickettsiae in the X. gerbilli, a species of fleas known to harbor/vector Y. pestis, the causative agent of plague in Central Asia (Stenseth et al. 2006, Wilschut et al. 2013). Also, there may be other RFLO associated with these fleas that were not described due to the limitation of species-specific assays in this study. Multilocus sequence typing would have helped to characterize some of the Rick17b, RfelG, and RfelB positive samples (Luce-Fedrow et al. 2015b) but that capability was unavailable at the time of this study.
R. felis was found rarely in the fleas and locations assessed and for R. typhi, not at all. Thus, the risk of flea-borne spotted fever from great gerbil fleas in the five rayons surveyed is low. Further investigation of flea-associated rickettsiae is needed throughout Kazakhstan and among other flea species to adequately assess the risk of flea-borne rickettsioses.
Conclusion
Fleas captured from R. opimus were found to be infected with rickettsiae. The most commonly found rickettsia-infected flea species was X. gerbilli, and the most commonly found rickettsia was R. asembonensis. Additional surveys for flea-borne rickettsiae are needed to investigate more completely the presence, prevalence, and distribution of these agents in other regions of Kazakhstan. In addition, it would be important to assess more completely the relationship between RFLO and the human pathogens R. felis and R. typhi as well as Y. pestis among the great gerbil fleas.
Footnotes
Acknowledgments
The views expressed here are those of the authors and do not necessarily represent the official policy or position of the Department of the Navy, the Department of Defense, the U.S. Government, or the Republic of Kazakhstan. This project was funded by the Defense Threat Reduction Agency, work unit number A1266. A.L.R. and C.M.F. are employees of the U.S. Government, and their work was prepared as part of their official duties. Title 17 U.S.C. §105 provides that “Copyright protection under this title is not available for any work of the United States Government.” Title 17 U.S.C. §101 defines a U.S. Government work as a work prepared by a military service member or employee of the U.S. Government as part of that person's official duties.
Author Disclosure Statement
No competing financial interests exist.