
Abstract
Most emerging zoonotic pathogens originate from animals. They can directly infect humans through natural reservoirs or indirectly through intermediate hosts. As a bridge, an intermediate host plays different roles in the transmission of zoonotic pathogens. In this study, we present three types of pathogen transmission to evaluate the effect of intermediate hosts on emerging zoonotic diseases in human epidemics. These types are identified as follows: TYPE 1, pathogen transmission without an intermediate host for comparison; TYPE 2, pathogen transmission with an intermediate host as an amplifier; and TYPE 3, pathogen transmission with an intermediate host as a vessel for genetic variation. In addition, we established three mathematical models to elucidate the mechanisms underlying zoonotic disease transmission according to these three types. Stability analysis indicated that the existence of intermediate hosts increased the difficulty of controlling zoonotic diseases because of more difficult conditions to satisfy for the disease to die out. The human epidemic would die out under the following conditions: TYPE 1:
Introduction
Z
Natural reservoirs maintain the existence of pathogens, and intermediate hosts provide the link for pathogen transmission. For example, wild aquatic birds are the natural reservoirs of the avian influenza virus, and domestic birds and pigs are its intermediate hosts (Fang 1981, Saenz et al. 2006). Initially, the avian influenza virus mainly infected birds. However, the virus has developed over time a capability to infect mammals such as pigs and humans. Although avian influenza viruses originally came from birds, the biggest human epidemics caused by them were from pigs, such as the Spanish Flu, which became a pandemic in 1918. In the transmission of the avian influenza virus, pigs act as “mixing vessels” for avian and human strains, which can produce super strains from genetic recombination that can be more easily transmitted to humans (Saenz et al. 2006, Coburn et al. 2011).
Pathogen transmission among species should be given serious attention because of expanding population and the changing animal–human interface, which have broken species barriers between humans and animals (Fang 1981, Dietz et al. 1988, Lanciotti et al. 1999, Bengis et al. 2004, Feng et al. 2004, Bowman et al. 2005, Hayes et al. 2005, Kramer et al. 2008, Kilpatrick and Randolph 2012, Mackenzie et al. 2013, Atlas and Maloy 2014, Sing 2015). Wildlife, domestic animals, and humans in the ecosystem have constructed the transmission net for zoonotic pathogens. In this net, humans can be infected directly or indirectly. Although human contact with wildlife as natural reservoir occurs rarely, human contact with domestic animals as intermediate hosts often occurs in livestock-based agricultural production (Bengis et al. 2004).
The influence of intermediate hosts on pathogen transmission is important in epidemiology, particularly in the prevention and control of emerging zoonoses. However, experiments on intermediate hosts in epidemiology with controls are difficult to design; sometimes, data are incomplete or inaccurate. In the present study, we use mathematical models to describe pathogen transmission from natural reservoirs to intermediate hosts and then to humans. Mathematical models in epidemiology can elucidate the mechanisms underlying disease transmission and provide a way to more quickly and easily determine the effect of intermediate hosts on zoonotic pathogen transmission (Fred and Carlos 2012).
The construction of a real transmission net requires complexity; however, the different roles of animals in human epidemics is considerably easier to identify if the infectious animals possess the ability to infect themselves or infect the next species, which is suitable for compartmental models with the population divided into several compartments and with assumption concerning the nature and time rate of transfer from one compartment to another (Fred and Carlos 2012). In our models, we classified zoonoses into several categories by their different transmission characteristics; we also divided wildlife, domestic animals, and humans into susceptibles (S), infectives (I), and recovered individuals (R).
Methods
Zoonotic pathogen transmission is mainly classified into 3 types according to the effect of intermediate hosts, as shown in Figure 1.

Transmission from natural reservoirs to intermediate hosts and then to humans under TYPE 1, TYPE 2, and TYPE 3.
TYPE 1: No intermediate host
For HIV, human–human transmission has considerably harmed humans in the last several decades, although initially, AIDS was not considered a human infectious disease. Scientists have discovered by gene sequence analysis that chimpanzees are the natural reservoirs of HIV. Regardless, the exact chimpanzee–human transmission remains undetermined (Sharp and Hahn 1995, Zacharova et al. 1997, Gao et al. 1999). HIV transmission initially conformed to TYPE 1 with animal–human and human–human transmissions. However, after decades of development, AIDS has become more similar to human infectious diseases than zoonotic diseases because of the number of infected humans who fail to recover, such as in the case of tuberculosis. TYPE 1 also matches human–human and human–animal transmissions, such as in the case of tuberculosis in which humans act as reservoir to infect other animals, particularly domestic ones (Fritsche et al. 2004).
Under TYPE 1, pathogens can be directly transmitted from natural reservoirs to humans (Zhang et al. 2012). This type of zoonosis can be transmitted without an intermediate host (Fig. 2). TYPE 1 is the condition set for the control group to compare the different effects of intermediate hosts under different conditions.
Transmission cycle under TYPE 1.
For TYPE 1, we can establish a multi-SIR model (1) to reflect the transmission from natural reservoirs to humans.
Each species, including natural reservoirs (N) and humans (H), is assumed to be divided into 3 epidemiological compartments: susceptibles (Si ), infectives (Ii ), and recovered individuals (Ri ), where i = N or H. We thus establish an ordinary differential equation (ODE) model (1) to reflect TYPE 1 transmission. Susceptibles can become infected through intra- or interspecies transmission and then recover with temporary immunity. To simplify the models, transmission of pathogens from the natural reservoir to the intermediate host and then to humans is assumed to be unidirectional, with a low probability of occurrence for transmission in the other direction.
Ai and μ i represent the natural natality rate and mortality rate, respectively. γ i denotes the recovery rate, and α i is the disease-induced mortality rate. β i denotes the per capita incidence rate in i, and β ij represents the per capita incidence rate from i to j, where i, j = N or H.
TYPE 2: Intermediate host as amplifier
Bat-related zoonoses (Sing 2015), derived from Chiropteran species, have given rise to many emerging infectious diseases exhibiting the transmission process under TYPE 2, including Ebola, SARS, MERS, Hendra (Mahalingam et al. 2012), and Nipah, among others.
In 2004, Leroy indicated that Ebola outbreaks in humans were accompanied with multiple simultaneous epidemics in gorilla, chimpanzee, and duiker carcass in Gabon and the Republic of Congo (Leroy et al. 2004). He stated that “Almost all human Ebola outbreaks in Gabon and RC have been linked to the handling of dead animals by villagers or hunters, and increased animal mortality always preceded the first human cases” (Leroy et al. 2004). He regarded scavenging for meat from dead apes as a process of Ebola virus transmission between a human and a nonhuman primate. Regardless, the natural reservoir of Ebola virus at the time was unknown to Leroy (Leroy et al. 2005, 2009, Guyton and Brook 2015).
A year later, Leroy presented in another article that the fruit bat was the natural reservoir of the Ebola virus after testing for the virus in more than 1000 small vertebrates in Africa (Leroy et al. 2005). Although nonhuman primates were not the natural reservoir species of the Ebola virus, they played an important role in Ebola transmission. The migration of a massive number of fruit bats with Ebola virus annually was speculated as the source of the Ebola epidemic. The beginning of the human Ebola epidemic could be traced to an infected dead ape (Leroy et al. 2005, 2009, Guyton and Brook 2015). Although the complexity of natural reservoir–intermediate host–human transmission increases the difficulty of zoonotic diseases research, mathematical models in epidemiology provide a way to more quickly and easily determine the zoonotic diseases transmission.
Under TYPE 2, pathogens can be directly or indirectly transmitted from natural reservoirs to humans. However, they can always be transmitted to humans through intermediate hosts. We assume that the pathogens can infect humans, but their natural reservoirs are too small to contain sufficient pathogens to transmit. Intermediate hosts act as amplifiers to maintain the transmission from natural reservoirs to humans (Fig. 3).
Transmission cycle under TYPE 2.
Apes acted as intermediate hosts (amplifiers) in Ebola outbreaks exhibiting transmission under TYPE 2, which is described in Model (2).
Compared with TYPE 1, TYPE 2 added intermediate hosts (I) as a new species in pathogen transmission. AI and μ I represent the natural natality rate and mortality rate of intermediate hosts, respectively. γ I denotes the recovery rate of intermediate hosts, and α I is the disease-induced mortality rate of intermediate hosts. β I represents the per capita incidence rate in intermediate hosts, and β NI is the per capita incidence rate from natural reservoirs to intermediate hosts. β IH is the per capita incidence rate from intermediate hosts to humans.
TYPE 3: Intermediate host as vessel for genetic variation
Pigs should be considered in the study of avian influenza virus transmission. The pig species is one of the intermediate hosts that act as a mixing vessel for the recombination of species-specific influenza virus strains from humans and birds. Coburn established a model to describe the effects of the pig species as a mixing vessel (Coburn et al. 2011). Meanwhile, the natural reservoir of the influenza is the bird species. Humans can be directly infected with several influenza subtypes from birds, including H5N1, H7N9, and H7N1, among others. However, the avian influenza virus, which can more easily cause a large-scale human epidemic, has infected pigs; examples of this virus are H1N1and H3N2 (Coburn et al. 2009, Busquets et al. 2010, Compans and Oldstone 2014).
Under TYPE 3, pathogens cannot be transmitted from natural reservoirs or intermediate hosts to humans, but can be transmitted to humans through intermediate hosts after reassortment or mutation. The assumption is that pathogens cannot infect humans but have the opportunity for variation to strains that are transmissible to humans. Intermediate hosts act as a vessel to strengthen the ability of the pathogens for variation to strains that are transmissible to humans (Fig. 4).

Transmission cycle under TYPE 3.
The transmission of H1N1 to pigs can be regarded as a process described under TYPE 3 in Model (3) (Iwami et al. 2007, Dorjee et al. 2013).
TYPE 3 includes a special variable, ɛ, which represents the variation rate. VI represents the number of intermediate hosts infected with pathogens after the variation; γV is the recovery rate of VI ; αV denotes the disease-induced mortality rate of VI ; and β V represents the per capita incidence rate of VI in intermediate hosts.
Results
This study aims to evaluate the effect of intermediate hosts on different emerging zoonoses. We construct three mathematical models depicting different transmissions to describe the roles of intermediate hosts. For the natural reservoir epidemic model, we can determine the basic reproduction number of natural reservoirs
If
Numerical simulation results under TYPE 1, TYPE 2, and TYPE 3 are shown in Figures 5 –11. To clearly compare the effect of intermediate hosts, we choose similar parameters (shown in Appendix B) under the three transmission types. The initial numbers of natural reservoirs, intermediate hosts, and humans are 12,000, 1000, and 2000, respectively (China Agricultural Yearbook Editing Committee 2012). Figure 5 shows the prevalence of the epidemic under different types. The assumption are that RN0 = RI0 = RV0 = 2 and RH0 = 1.2 (Saenz et al. 2006). The human epidemic would vary according to transmission types. The curve depicting the human epidemic exhibits different trends according to the different roles of intermediate hosts, including epidemic peak time and the number of infected individuals at peak time. Comparison of the curve depicting the human epidemic under TYPE 2 with that under TYPE 3 indicates that the variation rate can delay the peak time as more time is required to accumulate more infected individuals that can infect next infectious species.
Epidemic prevalence curves for natural reservoirs, intermediate hosts, and humans under TYPE 1
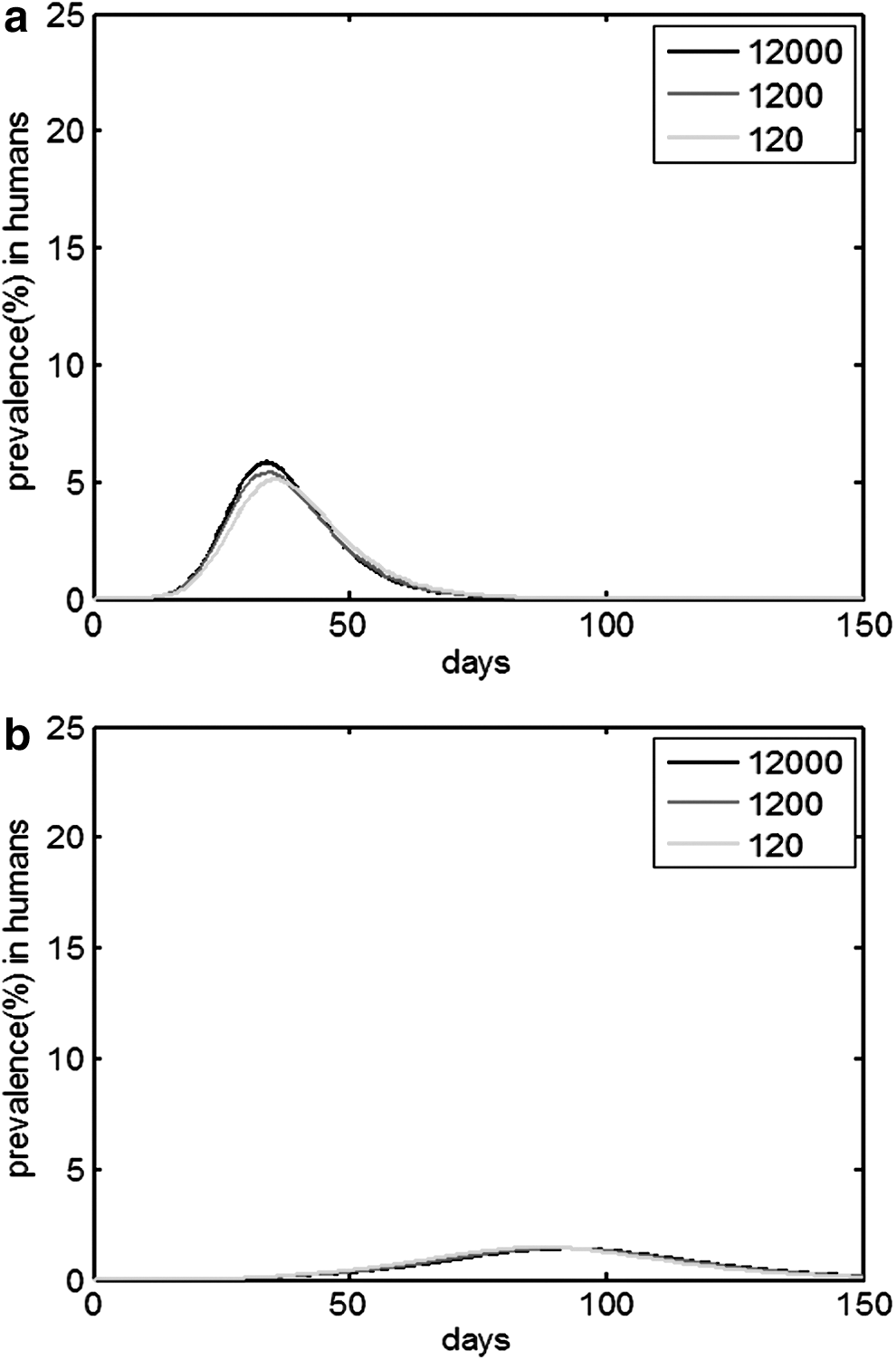
Figure 6 shows the prevalence of the human epidemic in different scales of natural reservoirs under TYPE 1. The assumption is that RN0 = 2 and RH0 = 1.2; however, the initial number of natural reservoirs varies from 12,000 to 1200 and then to 120. If no intermediate host exists, the percentage of infected humans decreases as the scales of natural reservoirs decrease.
Epidemic prevalence curves for humans in different scales of natural reservoirs from 12,000, to 1200, to 120 under TYPE 1.
Figure 7 shows the prevalence of the human epidemic in different scales of natural reservoirs under TYPE 2 and TYPE 3. Figure 8 shows the prevalence of the human epidemic in different scales of intermediate hosts under TYPE 2 and TYPE 3. The assumptions are that RN0 = RI0 = RV0 = 2 and RH0 = 1.2; however, the initial number of natural reservoirs varies from 12,000 to 1200 and further decreases to 120, as shown in Figure 7. The number of intermediate hosts varies from 10,000 to 1000 and further to 100, as shown in Figure 8. If intermediate hosts exist, the scales of intermediate hosts determine the prevalence of the human epidemic comparing Figures 6 –8. The percentage of infected humans decreases as the scales of intermediate hosts decrease.
Epidemic prevalence curves for humans in different scales of natural reservoirs from 12,000, to 1200, to 120 under TYPE 2

Epidemic prevalence curves for humans in different scales of intermediate hosts from 10,000, to 1000, to 100 under TYPE 2
Figure 9 presents the curve depicting the prevalence of the human epidemic under TYPE 2. The assumptions are that RN0 = 2 and RH0 = 1.2, 1.1, 0.9; however, RI0 = 2 in a, RI0 = 1.5 in b, and RI0 = 0.9 in c. If RN0 > 1, an epidemic of emerging zoonoses in humans could occur whether RH0 > 1 or not. Figure 10 shows the curve depicting the prevalence of human epidemic under TYPE 3. The assumptions are that RN0 = 2 and RH0 = 1.2, 1.1, 0.9; however, RI0 = RV0 = 2 in a, RI0 = RV0 = 1.5 in b, and RI0 = RV0 = 0.9 in c. Although only a small proportion of people would be infected under TYPE 3, for a small variation rate ɛ, an outbreak of emerging zoonoses would always occur when RN0 > 1.

Epidemic prevalence curves for humans in different RH0
, RH0 = 1.2, 1.1, 0.9 under TYPE 2 with RI0 = 2
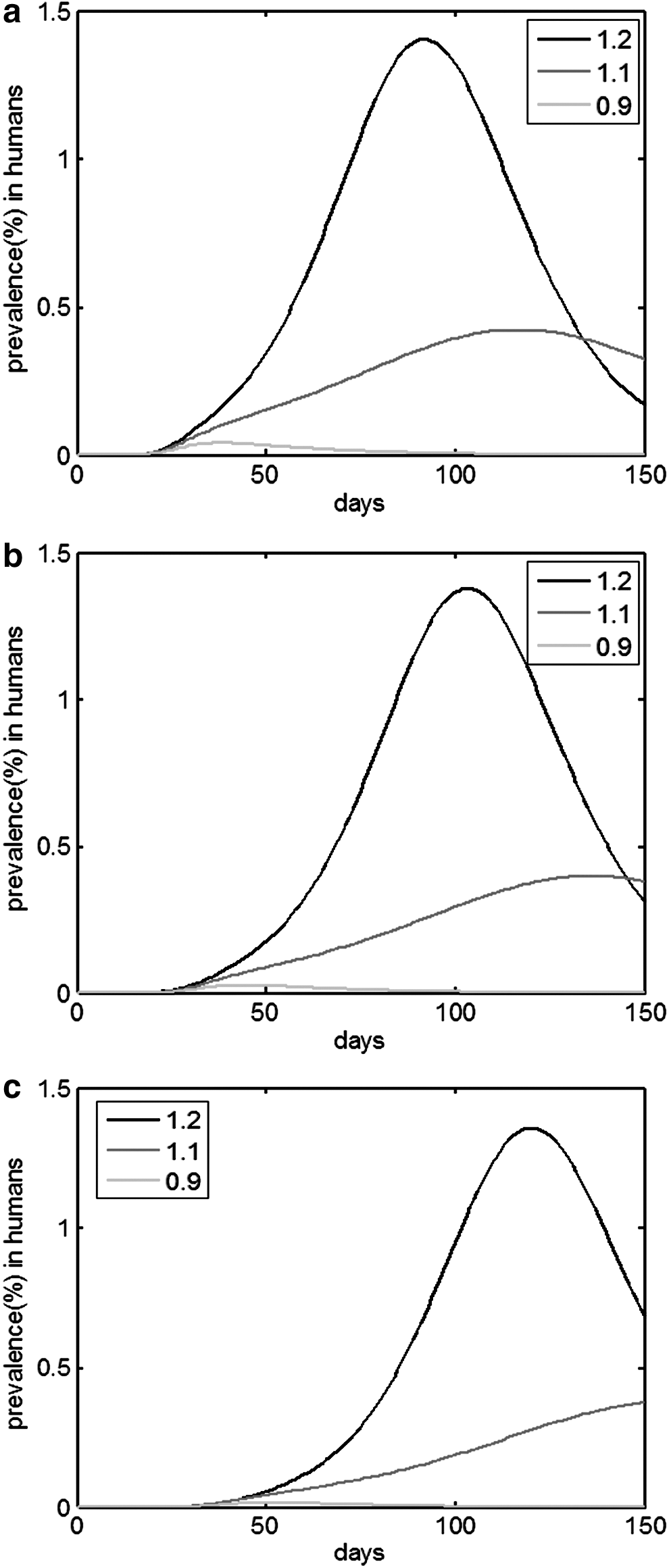
Epidemic prevalence curves for humans in different RH0
, RH0 = 1.2, 1.1, 0.9 under TYPE 3 with RI0 = RV0 = 2
However, if natural reservoirs or intermediate hosts are not taken into consideration in Model (4), the human epidemic would die out when RH0 < 1, as shown in Figure 11. The condition RN0 > 1 in natural reservoirs can determine the outbreak of zoonoses in humans; meanwhile, intermediate hosts do not possess this characteristic. In addition, when RN0 > 1, an epidemic occurs. The intermediate hosts can control the peak time and number of infected humans, which is much stronger than natural reservoirs. Fewer humans would be infected with a lower scale of intermediate hosts and a smaller basic reproductive number in intermediate hosts. However, the number of infected humans does not decrease as much with the same low scale of natural reservoirs.
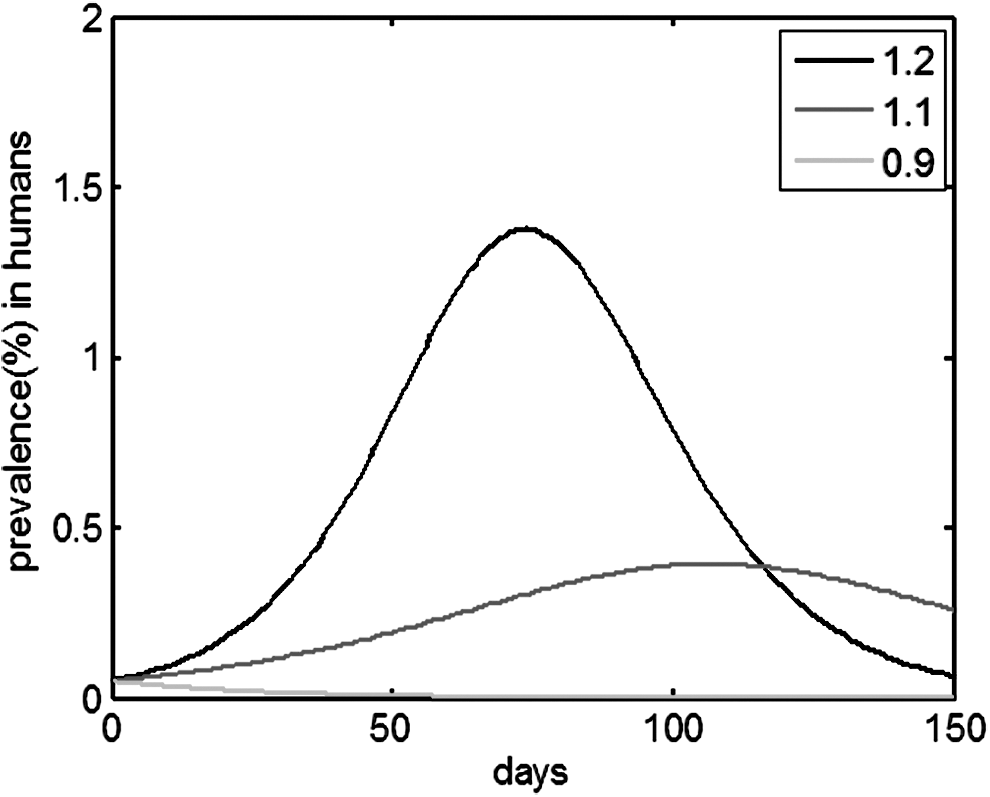
Epidemic prevalence curves for humans in different RH0 , RH0 = 1.2, 1.1, 0.9 of Model (4).
Discussion
In this study, we present three types of pathogen transmission to evaluate the effect of intermediate hosts on emerging zoonotic diseases in human epidemics. These types are identified as follows: TYPE 1, pathogen transmission without an intermediate host for comparison; TYPE 2, pathogen transmission with an intermediate host as an amplifier; and TYPE 3, pathogen transmission with an intermediate host as a vessel for genetic variation. In addition, we established three mathematical models to elucidate the mechanisms underlying zoonotic disease transmission according to these three types. Although the complexity of natural reservoir–intermediate host–human transmission increases the difficulty of zoonotic diseases research, mathematical models in epidemiology provide a way to more quickly and easily determine the zoonotic diseases transmission.
By stability analysis and numerical simulation, we find that animals play different roles in human epidemics under different assumptions. The intermediate host as an amplifier or vessel can change the peak time of a human epidemic and the total number of infected humans. Simultaneously, the existence of natural reservoirs and intermediate hosts can increase the difficulty of controlling emerging zoonoses. The effect of intermediate hosts on emerging zoonoses is reflected in the decrease in the prevalence of human epidemics, whereas some other forms of zoonoses already cause an outbreak; this effect of intermediate hosts varies from that of natural reservoirs. Natural reservoirs exert less effect on the prevalence of human epidemics when intermediate hosts exist, but they can ultimately determine whether zoonosis can become an outbreak in humans or in all members. Being able to distinguish the different effects of species on zoonotic pathogen transmission can be significant in addressing problems in public health.
More research on natural reservoirs should be conducted before or after the occurrence of a zoonotic outbreak. If a zoonotic disease dies out, human infection would not be a concern. Nonetheless, if a zoonotic disease was already an outbreak and we were living in the prevalence, focus should be directed toward controlling intermediate hosts rather than studying natural reservoirs to obtain better results.
In addition, the existence of intermediate hosts would change the peak time of an epidemic. Delay in peak time provides a greater opportunity to implement control measures; however, it also leads people to ignore the severity of emerging zoonoses initially affecting a small number of humans. Monitoring susceptible animals as intermediate hosts is important in the control of emerging and reemerging zoonoses.
Li verified that Rhinolophus is the natural reservoir of SARS-like coronaviruses (Li et al. 2005). The masked palm civet and two other species were identified as intermediate hosts infecting humans. They were found in live animal markets in China, where the first case of SARS was first reported. The transmission from a natural reservoir to an intermediate host used to be considered an incidental event. However, with the expanding population, deforesting, and other environmental changes caused by anthropogenic factors, contact between susceptible animals and humans has increased, immediately influencing pathogen transmission. A monitoring network for susceptible animals as natural reservoirs or intermediate hosts should be established to promptly respond to emerging zoonoses.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (11371048).
Author Disclosure Statement
No competing financial interests exist.