
Abstract
Alkhumra hemorrhagic fever virus (AHFV) is a newly described zoonotic flavivirus that was first isolated during 1994–1995 from the Alkhumra district south of Jeddah, Saudi Arabia. Subsequently, the virus was also isolated from Makkah city (2001–2003) and Najran (2008–2009), Saudi Arabia. The virus causes acute febrile illness with hepatitis, hemorrhagic manifestations, and encephalitis. A case fatality rate of 25% was reported among hospitalized patients. Although several biological and molecular characteristics of the virus have been published, no data are available on electron microscopic features of the virus. In this article, we describe the morphological features and metrics of the AHFV particles under electron microscopy, and localization of the virus particles in brain cells of newborn Wistar rats and in Rhesus monkey (Macaca mulatta) kidney epithelial cells (LLC-MK2). Virus particles in both the LLC-MK2 cells and the rat brain cells showed dark hexagonal core (capsid) and a translucent envelope. The mean diameter of the enveloped virus particle was 40.59 ± 1.29 nm in the rat brain cells (n = 154) and 40.97 ± 1.40 nm in the LLC-MK2 cells (n = 105; p > 0.05). The virus particles, both in vitro and in vivo, were enclosed into cytoplasmic vesicles. In conclusion, the shape, size, and diameter of the AHFV particle lie within the framework of the genus Flavivirus, family Flaviviridae.
Introduction
A
From 2003 to 2007, eight confirmed cases of AHFV infections were sporadically reported from Najran in southern Saudi Arabia (Madani et al. 2011). Subsequently, an outbreak of AHFV infection occurred in Najran during 2008–2009 with 70 confirmed cases reported (Madani et al. 2011).
AHFV was reported only from Saudi Arabia until 2010 when two travelers returning to Italy from southern Egypt were confirmed to be infected with AHFV (Carletti et al. 2010). A recent study showed that ticks collected from cattle in Djibouti were infected with the virus (Horton et al. 2016). The latter two studies suggest that the geographic distribution of AHFV likely goes beyond Saudi Arabia.
The virus causes acute febrile illness with hepatitis, hemorrhagic manifestations, and encephalitis (Madani 2005, Madani et al. 2011). A case fatality rate of 25% was reported among hospitalized patients. Because of the close phylogenetical relationship between AHFV and the Kyasanur Forest disease virus (Mehla et al. 2009) that is transmitted by ticks (Bhat and Goverdhan 1973), AHFV was classified within the tick-borne encephalitis group in the genus Flavivirus of the family Flaviviridae (Pletnev et al. 2011).
This study presents the electron microscopic images of AHFV and localization of the virus particles in brain cells of newborn rats and in Rhesus monkey kidney epithelial culture (LLC-MK2) cells, and describes the morphological features and metrics of the virus particles in comparison with those reported for other members of the genus Flavivirus.
Materials and Methods
The virus
The AHFV used in this study, designated (AHFV/997/NJ/09/SA), was isolated originally from a patient's blood sample inoculated into newborn Wistar rat brains during the outbreak of the disease that occurred in Najran city, southern Saudi Arabia, in 2008–2009 (Madani et al. 2011). The virus was adapted and titrated in LLC-MK2 cell culture monolayers using 96-well cell culture microplates as previously described (Madani et al. 2014a, 2014b). Five replicate wells were employed. The plates were incubated at 37°C in CO2 atmosphere. The median tissue culture infective dose (TCID50) was calculated as described by Reed and Muench (1983). Its titer was 107.2 TCID50/mL.
The cell culture
The continuous Rhesus monkey (Macaca mulatta) kidney epithelial cells (LLC-MK2 cell line) were employed in this study. The selection of this cell line was based on a previous study showing that this cell line was superior to other cell lines used for growth of AHFV (Madani et al. 2016).
Preparation of the AHFV from the LLC-MK2 cell culture for electron microscopy
The LLC-MK2 monolayers were grown in three 75 cm2 Corning tissue cell culture flasks at 37°C in Eagles minimum essential medium (EMEM) supplemented with 10% fetal calf serum (FCS; Sigma, St. Louis). When the monolayers were 70% confluent, each flask was inoculated with 1 mL of the prototype AHFV in the form of undiluted newborn rat brain suspension, passage 1, and then incubated at 37°C for 1 h to adsorb. This was followed by the addition of 15 mL of EMEM containing 2% FCS and incubation at 37°C. The monolayers were observed daily under an inverted microscope for the presence of discernible cytopathic effect (CPE). When the CPE affected 30% or greater of the monolayer, the three flasks were removed from the incubator and the media were collected and pooled. The cell sheet from the surface of each flask was gently scraped by a cell scraper (Falcon, NJ). Fifteen milliliters of the pooled media was added to the scraped cells in each flask. The flasks were then shaken gently and the contents of the three flasks were pooled. This pool was spun in a cooled bench centrifuge (5702; Eppendorf) at 141.98 g (1000 rpm) for 10 min. The pellet was resuspended in 5 mL phosphate-buffered saline (PBS) pH 7.4 and fixed in 2.5% glutaraldehyde in PBS pH 7.4 for 2 h at room temperature. This was followed by three successive rinsing in distilled water, fixation in PBS–1% osmium tetroxide for 20 min, and prestaining en bloc with 2% uranyl acetate for 10 min and rinsing with distilled water. This was followed by dehydration in an ethanol series as follows: 50% ethanol for 3 min, 70% ethanol for 3 min, 90% ethanol for 3 min, and two changes of 100% ethanol for 3 min each. The dehydration process was followed by two changes of propylene oxide for 5 min each. This was followed by infiltration with a mixture of propylene oxide and epoxy (PELCO® Eponate 12™ Kit; Ted Pella, Inc.) with a final infiltration in pure resin and final embedding in fresh epoxy resin. Thin sections (70–90 nm thick) were cut from the block and stained in 4% uranyl acetate and Reynold's lead citrate and examined using a transmission electron microscope (Philips—CM 100, The Netherlands). Sections of noninoculated LLC-MK2 cells were prepared and treated as mentioned and examined under an electron microscope.
AHFV inoculation into newborn Wistar rats
Eleven in-house bred 2-day-old Wistar rats, herein referred to as “rats,” were used in the experiments. All inoculations, care, and handling of the inoculated rats were performed in a biosafety level-3 facility. Seven rats were each inoculated intracerebrally with 20 μL of the AHFV as described previously (Madani et al. 2014b). The remaining four rats were each injected intracerebrally with EMEM and kept as controls. All the rats were observed daily for any clinical manifestations or death. The brains of dead or moribund rats were harvested, placed in 2.5% glutaraldehyde, and processed for electron microscopy as described hereunder.
Preparation of the AHFV from the baby rat brains for electron microscopy
A transmission electron microscope (Philips—CM 100, The Netherlands) was calibrated to observe the virus in the sections and to measure its dimensions. The brains from the dead, the moribund, and the control rats were removed and fixed in 2.5% glutaraldehyde in PBS pH 7.4 for 2 h at room temperature. Further fixation, embedding in fresh epoxy resin, thin sectioning (70–90 nm thick), and staining in 4% uranyl acetate and Reynold's lead citrate were performed followed by examination under a transmission electron microscope.
Results
The AHFV-inoculated rats started to show mild neurological signs on day 3 postinoculation. From day 4, the neurological manifestations progressed into severe tremors, irritability, convulsion, opisthotonos, and spastic paresis. All the AHFV-inoculated rats, as compared with the control rats, were emaciated and dehydrated. Five of the inoculated rats died within 5–7 days postinoculation. The remaining two moribund rats were euthanized.
The electron microscopic examination of sections of both the AHFV-inoculated LLC-MK2 cells and rat brains revealed presence of spherical virus particles inside cytoplasmic vesicles (Figs. 1 and 2). Arrays of virus particles were also seen in the cytoplasm of the rat brain cells (Fig. 3). Despite thorough examination of sections from the inoculated LLC-MK2 and rat brain cells, no virus particles were seen in the nucleus or the mitochondria. The noninoculated LLC-MK2 and rat brain cells showed intact cellular organelles with defined structures and membranes. The virus particles in both the LLC-MK2 cells and the rat brain cells showed dark hexagonal core (capsid) and a translucent envelope (Figs. 3 and 4). The mean diameter of the enveloped virus particle was 40.59 ± 1.29 nm in the rat brain cells (n = 154 virus particles) and 40.97 ± 1.40 nm in the LLC-MK2 cells (n = 105 virus particles). The difference between the mean diameter of the virus particles in the LLC-MK2 and the rat brain cells was not statistically significant (p > 0.05).

Electron micrograph showing the presence of numerous virus particles in intact and ruptured cytoplasmic vesicles in a section from AHFV-inoculated LLC-MK2 cells (arrows). AHFV, Alkhumra hemorrhagic fever virus.
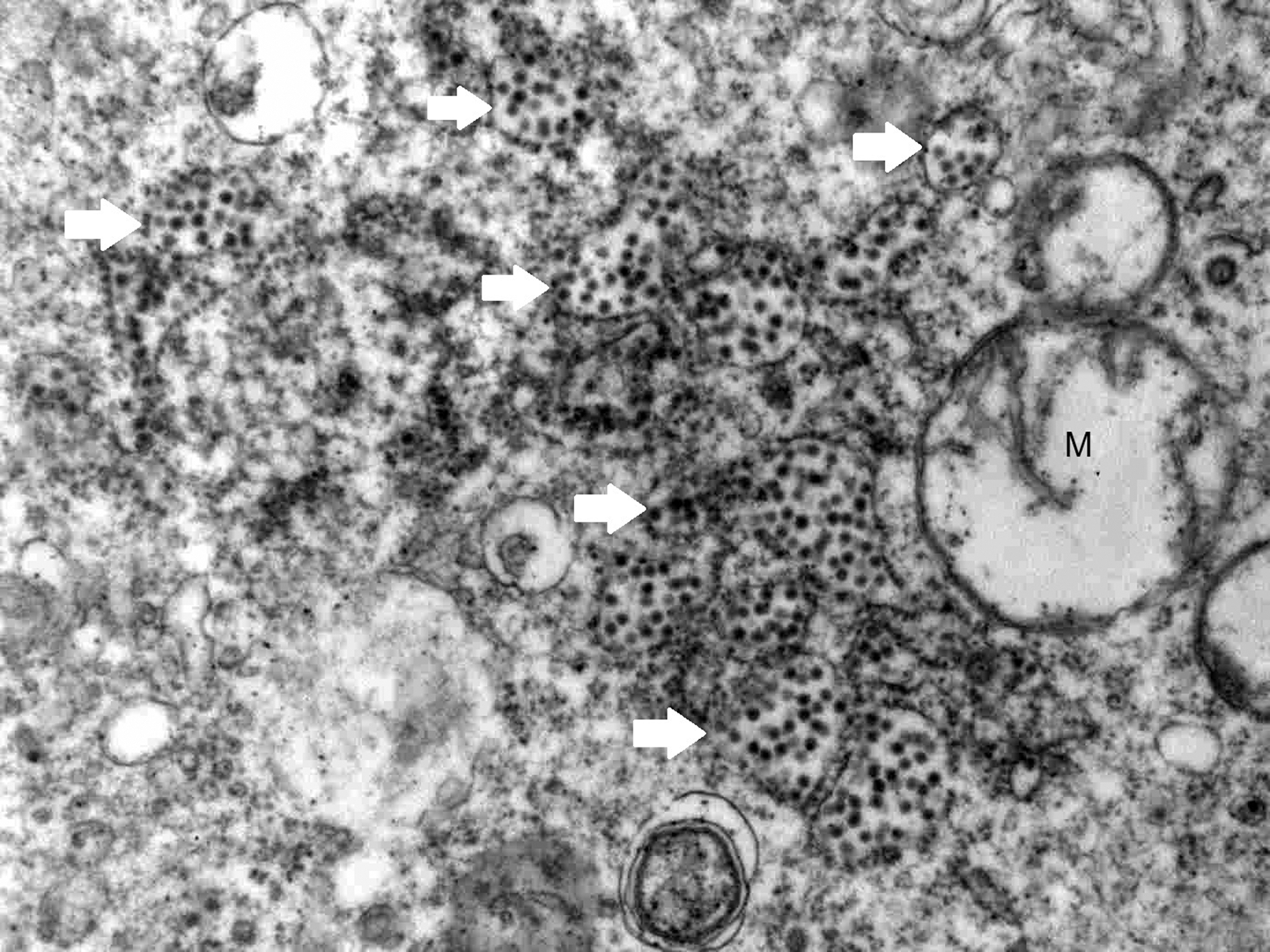
Electron micrograph of a section of inoculated rat brain cells, showing numerous intact and ruptured virus-containing vesicles (arrows) near mitochondria (M).

Electron micrograph showing arrays of whole enveloped virus particles in a section of rat brain inoculated with the AHFV with a dark core representing the capsid, and a translucent envelope.
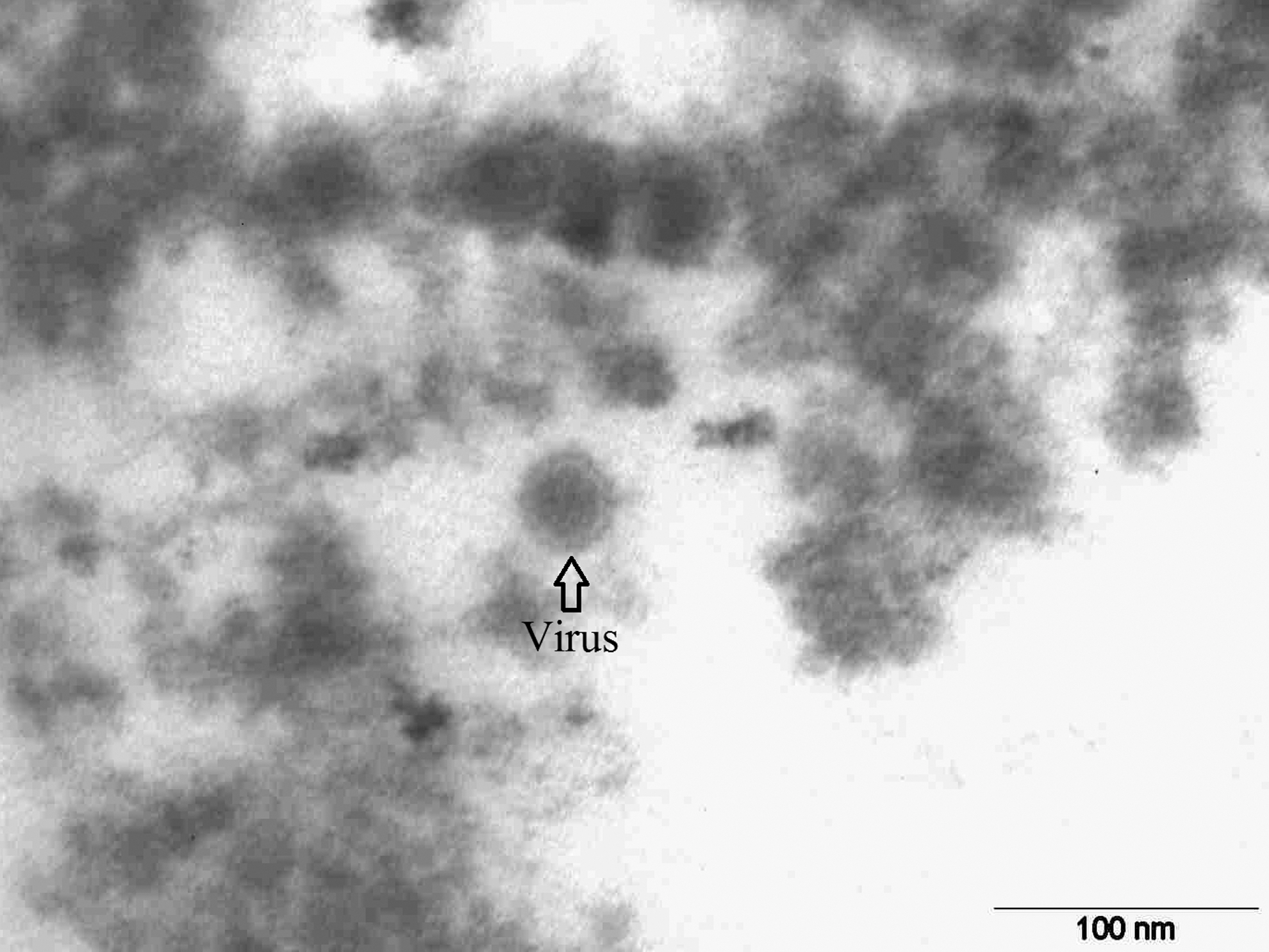
A single enveloped virus particle with conspicuous hexagonal capsid.
Discussion
Since the early reports of observation of viruses by the electron microscope during the 1930s of the past century (Kausche et al. 1939), the electron microscope has been utilized extensively to get the best images of viruses. The electron microscope has been used with great success for virus diagnosis by using various techniques (Madeley 1997), observation of virus subunits, studying the architecture of viruses (Klose et al. 2010), understanding dynamic virus–cell interactions, and follow-up of intracellular life-cycle of viruses (Roingeard 2008). The electron microscope has also been useful for the initial identification of emerging viruses causing disease outbreaks (Roingeard 2008). The modern uses of electron microscopy in virus research were reviewed by Goldsmith and Miller (2009).
In this study, we used a simple procedure to prepare AHFV for electron microscopic examination. Thin section micrographs of both LLC-MK2 cells and rat brain cells infected with the AHFV showed large quantities of positively stained virus particles, each composed of a hexagonal capsid surrounded by a translucent envelope. The virus particles were predominantly seen inside vesicles within the endoplasmic reticulum of the cell cytoplasm. These features were also described for other members of the genus Flavivirus in cell culture, for example, dengue virus (Barth et al. 1997), a new flavivirus isolated from mosquitoes (Okuno et al. 1984), Japanese encephalitis virus (Okuno et al. 1984), and yellow fever virus (Gould et al. 1985, Yoshii et al. 2012).
According to the ICTV, the diameter of the whole enveloped virus particles of members of the genus Flavivirus is reported to be 40–70 nm (Pletnev et al. 2011). The results of this study showed that the diameter of the AHFV (40.59–40.97 nm) lied within that of the genus Flavivirus and that the other morphological characteristics are similar. There was no significant difference between the electron microscopic features of AHFV in vivo (rat brain cells) and in vitro (LLC-MK2 cells). In both systems, the virus size was similar and the virus multiplication was intracytoplasmic.
In conclusion, the simple procedure described in this study enabled observation of the AHFV successfully using the electron microscope. This success can pave the way to conduct more studies on various applications of electron microscopy in the field of AHFV research, including diagnosis of AHFV from clinical samples, employment of immunoelectron microscopy, cryoelectron microscopy, and ultrastructural studies in cell culture infected with the virus.
Footnotes
Acknowledgments
The authors thank Sheikh Mohammed Hussein Al-Amoudi for funding this research and the Scientific Chair for Viral Hemorrhagic Fever at King Abdulaziz University, Jeddah, Saudi Arabia. We thank Mr. Helmy Abdullah, Mr. Abdullah Al-Salawaty, and Mrs Ola Alahmadi for their excellent technical assistance, and Mr. Saad Alhusaiki for his assistance in preparing the images.
Financial Support
This study is one of the research products of the Scientific Chair of Mohammad Hussein Alamoudi for Viral Hemorrhagic Fevers, King Abdulaziz University, Jeddah, Saudi Arabia. The sponsor, Sheikh Mohammad Hussein Alamoudi, had no involvement in the study design, in the collection, analysis, and interpretation of data, in the writing of the article, or in the decision to submit the article for publication.
Authors' Contribution
T.A.M. and E.M.E.A. conceived and designed the study and wrote the article; E.M.E.A. performed isolation and titration of AHFV, designed and performed the procedure for preparing the virus for electron microscopy, and examined the virus under the electron microscope; S.M.J. facilitated the electron microscopy examination and processed the fixed AHFV samples for examination under the electron microscope; A.M.H. assisted in the preparation of LLC-MK2 cell culture and in the preparation, cultivation, and titration of the virus; H.A. assisted in the arrangement, setting up, performing the electron microscopy examination, and interpretation of micrographic images; E.I.A. and H.M.S.A. critically revised the article; and all authors read and approved the final article.
Ethical Approval
King Abdulaziz University's policy on the care and use of laboratory animals was followed. Ethical approval was obtained from the Research Ethics Committee at the Faculty of Medicine, King Abdulaziz University, Jeddah, Saudi Arabia.
Author Disclosure Statement
No competing financial interests exist.