
Abstract
Maintenance of sylvatic plague in prairie dogs (Cynomis spp.) was once thought unlikely due to high mortality rates; yet more recent findings indicate that low-level enzootic plague may be maintained in susceptible prairie dog populations. Another hypothesis for the maintenance of sylvatic plague involves small mammals, other than prairie dogs, as an alternative reservoir in the sylvatic plague system. These hypotheses, however, are not mutually exclusive, as both prairie dogs and small mammals could together be driving sylvatic cycles of plague. The concept of a bridging vector has been used to explain the transmission of pathogens from one host species to another. In the case of sylvatic plague, this would require overlap in fleas between small mammals and prairie dogs, and potentially other species such as carnivores. Our goal was to evaluate the level of flea sharing between black-tailed prairie dogs (Cynomis ludovicianus) and other small mammals in a mixed-grass prairie in South Dakota. We investigated the species richness of small mammals and small-mammal fleas in a mixed-grass prairie system and compared findings with previous studies from a short-grass ecosystem in Colorado. Over the summer field seasons 2014–2016 we live-trapped small mammals, collected fleas, and showed differences between both the flea and small mammal composition of the two systems. We also recorded higher densities of deer mice and lower densities of northern grasshopper mice in mixed versus shortgrass prairies. We confirmed, as is the case in shortgrass prairies, a lack of substantial flea species overlap on small mammal hosts and fleas from prairie dogs and their burrows. Moreover this study demonstrates that although small mammals may not play a large part in interepizootic plague cycling in shortgrass prairie ecosystems, their role in mixed-grass prairies requires further evaluation.
Introduction
T
There are two predominating hypotheses to explain dormancy maintenance in prairie dogs (Cynomis spp.), although they are not mutually exclusive. The first hypothesis is that interepizootic plague is maintained by periods of differential transmission and varying rates of infection in populations of prairie dogs, some of which may be resistant to plague (Rocke et al. 2012, have documented some resistance in prairie dogs from plague endemic regions), or through “smoldering plague transmission” envisioned as a cryptic transmission route in enzootic plague leading to essentially undetectable mortality rates in prairie dog colonies (Gage and Kosoy 2005, Hanson, et al. 2007, Salkeld et al. 2016). Hanson et al. (2007), Mize and Britten (2010), and Mize and Britten (2016), also provided evidence in support of the notion of interepizootic plague, detecting the presence of Y. pestis during interepizootic phases in fleas from prairie dogs in Montana, and Lower Brule, SD. The second involves enzootic plague in relatively resistant populations of small mammals (i.e., maintenance hosts) and their flea faunas, highlighting the potential importance of cryptic alternate reservoirs and spillover events into susceptible hosts (epizootic/amplification hosts) in the transmission of Y. pestis (Gage and Kosoy 2005, Salkeld and Stapp 2008); simply stated infected fleas survive on relatively resistant hosts, leading to spillover effects and occasional infections of relatively resistant hosts.
The importance of the second hypothesis, involving small mammals as potential alternative reservoirs has repeatedly been investigated, without uncovering definitive evidence (Gage and Kosoy 2005, Salkeld and Stapp 2008, Salkeld et al. 2010). The findings of Graham et al. (2014) have strengthened the argument for the importance of small mammals as a reservoir for sylvatic plague. Graham et al. (2014) outlined the three assumptions of the enzootic host model of the interepizootic maintenance of Y. pestis (here and after: enzootic host model) as: (1) Y. pestis persists between epizootics in a maintenance cycle between fleas and relatively resistant enzootic hosts (e.g., fleas must remain infected after feeding on resistant individuals); (2) infected fleas may infest different species, including both resistant and susceptible hosts; and (3) when a resistant individual is infected with Y. pestis it almost always seroconverts leading to resistant individuals, at least temporarily carrying antibodies to the pathogen and limiting disease. Graham et al. (2014) conclude that Xenopsylla cheopis and Oropsylla montana fleas remain infected after feeding on resistant hosts, demonstrating that at least one of the three assumptions has been met for the maintenance of sylvatic plague in small mammal populations. The conclusion of Graham et al. (2014) that fleas remain infected after feeding on resistant hosts strengthens the potential for small mammal interactions in assisting in the cultivation and maintenance of sylvatic plague. While the assumptions of Graham et al. (2014) may not be exhaustive, we aim to evaluate the second assumption of the aforementioned model in a mixed-grass system.
Flea compositions on small mammals in shortgrass prairie/steppe ecosystems have been repeatedly analyzed (Anderson and Williams 1997, Salkeld and Stapp 2008, Stapp et al. 2009, Franklin et al. 2010), yet little literature encompasses mixed-grass prairie ecosystems. Predominant themes among researchers are a lack of flea species overlap between small mammals and prairie dogs in shortgrass ecosystems, and a limited amount of data showing the ability of fleas from small mammals to maintain infections in interepizootic periods. One outlier is the northern grasshopper mouse (Onychomys leucogaster), which shares up to three species of flea with prairie dogs in the northern Colorado shortgrass prairie ecosystem (Salkeld and Stapp, 2009, Stapp et al. 2009). During epizootics Stapp et al. (2009) marked a host switch of flea species from prairie dogs to northern grasshopper mice upon the death of primary hosts, and asserted that northern grasshopper mice may drive plague epizootics in prairie dogs by increasing connectivity between colonies in shortgrass systems (Salkeld et al. 2010). This host commonality could provide the opportunity for infected fleas to infest additional species following the extirpation of primary hosts, supporting the first assumption of the enzootic host model (Graham et al. 2014) and the idea of small mammals as a reservoir for plague. The importance of northern grasshopper mice was investigated by Kraft and Stapp (2013) who concluded that at high densities, this species facilitates the spread of plague.
The role of small mammals in mixed-grass prairies has yet to be evaluated. Bridging vectors are responsible for the transmission of a pathogen from one host species to another. Some bridging vectors have greater potential for disease transmission, with specialist species perhaps being less likely than generalist species to function as a bridging vector (Eads et al. 2015). Oropsylla hirsuta, a specialist, has been documented to parasitize both black-tailed prairie dogs and northern grasshopper mice in shortgrass prairies using blood meal analysis (BMA) (Franklin et al. 2010) and stable isotopes (Stapp and Salkeld 2009). While this suggests that O. hirsuta may act as a bridging vector in sylvatic plague cycles, little evidence exists for fleas taking blood meals from small mammals and subsequently feeding on other small mammal species or prairie dogs. This indicates that O. hirsuta might not commonly function as a bridging vector, but under the right circumstances it may have the ability to fill the role. This study looked at the mixed-grass prairie ecosystem and the potential role of small-mammal fleas as bridging vectors between small mammals and prairie dogs in Lower Brule, South Dakota. The focus of this article is to further investigate the involvement of small mammals and their fleas in plague ecology on black-tailed prairie dog (C. ludovicianus) colonies in a mixed-grass prairie ecosystem. For the purposes of this study, we defined small mammals as nonprairie dog rodents and insectivores that can be captured using 7.6 × 8.9 × 22.9 cm aluminum live traps (slightly modified from Brinkerhoff et al. 2008).
Materials and Methods
Fleas were collected from live-trapped animals on 16 sites in two counties on the 53,613 hectare Lower Brule Sioux Indian Reservation in South Dakota (44.08°N 99.78°W), USA, from May 2014 to September 2016. This region is classified as mixed-grass prairie, dominated by western wheatgrass (Pascopyrum smithii), green needlegrass (Nassella viridula), buffalo grass (Buoteloua dactyloides), and needle-and-thread grass (Hesperostipa comata). Trapping took place on active prairie dog colonies (n = 8), as well as plague-extirpated colonies (n = 4) and native/restored prairie (n = 4), where prairie dogs were absent in a mixed-grass prairie ecosystem. Site selection was aimed at maximizing geographic coverage. As of 2016, there were a total of 82 active colonies spread across the reservation (S. Grassel, personal communication).
There was one trapping grid at each of the 16 sites. Each grid consisted of five parallel transects of ten Sherman live traps spaced fifteen meters apart for a total of 50 traps per grid. Traps were left open for three trap nights and two trap days to sample both nocturnal and diurnal animals. Each grid was visited two times per field season with an early trapping session in April/May, and a later trapping session in June/July. We did a third trapping session in August/September only on the set of grids with the highest trapping success in previous trapping sessions each year. Animals were anesthetized with isoflurane, marked, and measured, fleas were collected and stored in 95% ethanol, and small mammals were released alive for recapture purposes. Animal handling procedures were approved by the Institutional Animal Care and Use Committee (30-11-13-16C) at the University of South Dakota, following guidelines of the American Society of Mammalogists (Gannon and Sikes 2007). Fleas were identified to species using keys from Fox (1940), Furman and Catts (1982), Benton (1983), and Lewis (2000, 2002) under a dissecting microscope. Flea loads were determined by dividing the total number of fleas recovered by the total number of mammal captures/recaptures; this reflects captures/recaptures within trapping session and across seasons as available. The last substantial plague epizootic in Lower Brule occurred in 2013, and the first documented was in 2011 (S. Grassel, personal communication).
Whole flea DNA was extracted from individual fleas using the prepGEM® Insect Kit (Zygem, New Zealand) following the manufacturer's specifications. Nested PCR was used to survey fleas for the presence of Y. pestis. Methods were slightly modified from those of Jones et al. (2012), Jones and Britten (2010), and Hanson et al. (2007) and originally modified from Hinnebusch and Schwan (1993). DNA extracted from fleas was pooled by small mammal host for the Y. pestis assays.
Results
A total of 970 unique small mammals of eight species, including 706 deer mice (Peromyscus maniculatus), 64 western harvest mice (Reithrodontomys megalotis), 61 thirteen-lined ground squirrels (Ictidomys tridecemlineatus), 49 northern grasshopper mice, 37 hispid pocket mice (Chaetodipus hispidus), 24 prairie voles (Microtus pennsylvanicus), 23 plains harvest mice (Reithrodontomys montanus), and 6 northern short-tailed shrews (Blarina brevicauda) were trapped over field seasons 2014–2016. We found low densities of northern grasshopper mice, which made up only 5% (n = 49) of the unique captures (n = 970) (Fig. 1). Ten flea species were recovered (n = 1641): Aetheca wagneri (n = 1190), Orchopeas leucopus (n = 116), Peromyscopsylla hesperomys (n = 147), Pleochaetis exilis (n = 74), Epitidia wenmani (n = 38), O. hirsuta (n = 24), Thrassis fotus (n = 13), Thrassis bacchi (n = 10), Oropsylla tuberculata (n = 5), and Doratopsylla blarinae (n = 3) (Table 1).

Comparison of small mammal compositions from mixed-grass prairie in Lower Brule, South Dakota on the left, and in shortgrass prairies of the Pawnee National Grasslands, CO (Stapp and Salkeld 2009) on the right. This figure demonstrates the differences between the two systems principally in the most common small mammal species encountered in each system.
An overall flea load of 0.85 fleas per animal (range = 0–24, SD = 1.77) was found, with A. wagneri alone accounting for 0.65 (range = 0–10, SD = 1.75) fleas per animal. We observed an overwhelming abundance of A. wagneri (Fig. 2), which was found across all mammal species. A. wagneri represented 73% of all fleas detected having been found on 558 individual captures, with much smaller detection rates of all other flea species.
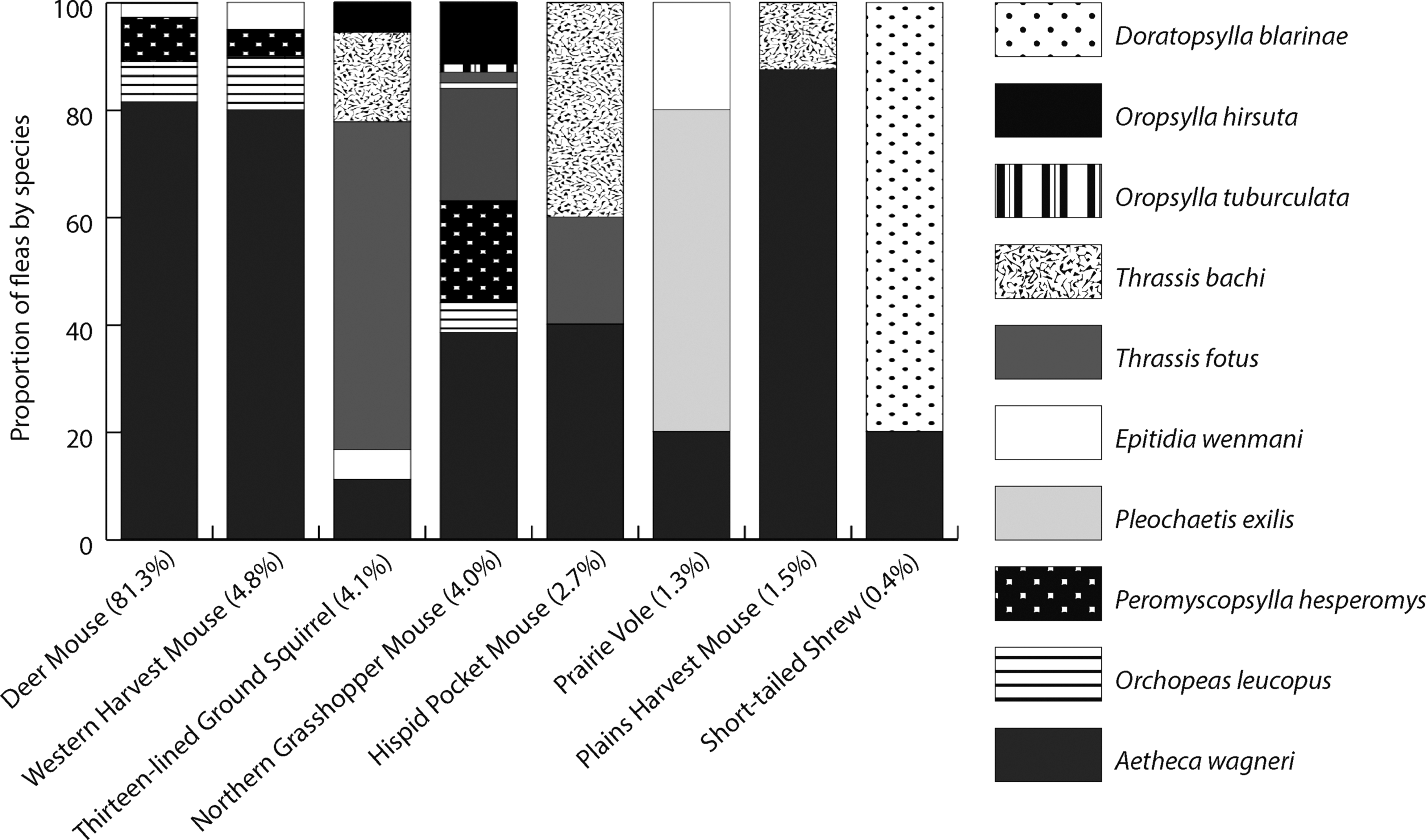
Flea composition and overlap showing that Aetheca wagneri was the most commonly encountered flea found across host species. Flea faunas infesting small mammals sampled at Lower Brule, South Dakota, 2014–2016. The numbers in parentheses are the proportions of the total of all sampled small mammals by species.
On the most commonly encountered small mammal, the deer mouse, fleas were detected on 589 captures/recaptures of 706 individual deer mice. Flea loads from these mice were 0.89 total fleas per animal (range = 0–14, SD = 1.64) and of those, 0.73 (range = 0–10, SD = 1.76) were A. wagneri. This equates to 505 deer mouse captures with A. wagneri. One deer mouse was infested with O. tuberculata (n = 2) for a flea load of 0.001 (range = 0–2, SD = 0.025) O. tuberculata per deer mouse; and one deer mouse was infested with T. fotus for a flea load of 0.0007 (range = 0–1, SD = 0.018) T. fotus per deer mouse (Table 2).
Fleas were detected on 51 captures/recaptures of 49 individual northern grasshopper mice. Northern grasshopper mice had the highest individual flea loads with 3.11 fleas per animal (range = 0–24, SD = 4.04), A. wagneri were found on 32 captures/recaptures of northern grasshopper mice representing a flea load of 1.04 (range = 0–7, SD = 1.64). There were 11 captures/recaptures of northern grasshopper mice, where O. hirsuta were detected and a flea load of 0.31 fleas per animal (range = 0–6, SD = 0.99). O. tuberculata and Thrassis bacchi were each detected on 2 separate individual grasshopper mice representing a flea load of 0.04 (range = 0–2, SD = 0.28) O. tuberculata, and 0.05 (range = 0–2, SD = 0.35) T. bacchi per animal, with the remainder being small numbers of fleas not known to be important in sylvatic plague maintenance (Table 2).
Fleas were found on 13 captures/recaptures of 61 individual thirteen-lined ground squirrels. We detected flea loads of 0.25 (range = 0–5, SD = 1.33) fleas per animal. Two thirteen-lined ground squirrels were infested with A. wagneri and we detected a flea load of 0.03 (range = 0–1, SD = 0.27). Five thirteen-lined ground squirrels were infested with T. fotus with a flea load of 0.15 (range = 0–5, SD = 1.06), and two individuals were infested with T. bacchi representing a flea load of 0.04 (range = 0–2, SD = 0.51). O. hirsuta was detected on one individual with a flea load of 0.01 (range = 0–1, SD = 0.15) O. hirsuta per animal (Table 2).
Of note was the presence of three species of vector-competent fleas, although in low numbers. These include fleas of the genera Thrassis and Oropsylla. We found that 1.77% of detected fleas were O. hirsuta, and O. tuberculata, which are putative vectors of epizootic plague in prairie dogs (Cynomis spp.) (Abbott and Rocke 2012), and found that 1.40% of the detected fleas were Thrassis spp., which are thought to be important in sylvatic plague maintenance in ground squirrels (Abbott and Rocke 2012). This results in a total of 3.17% of small mammal fleas being species of known importance in sylvatic plague maintenance. Small mammal flea composition data collected were compared with data from fleas collected from prairie dogs in the same area by Mize and Britten (2010) who found that 99% of fleas from prairie dogs were O. hirsuta, with the remaining 1% being Oropsylla that could not be identified to species.
Polymerase chain reaction showed 17 positive pools from a total of 362 pools (from a total of 832 captures/recaptures with fleas or 1607 total fleas). Fleas from P. maniculatus constituted 16 of the 17 positive pools, with one positive pool coming from fleas on an O. leucogaster. Two positive pools came from small mammals in native prairie plots, and one came from a plague extirpated site. The remainder of the positive pools came from active prairie dog colonies.
Discussion
Small mammal diversity and the associated flea communities differ markedly between short and mixed-grass ecosystems. Although, small mammal and flea populations are known to change across years and season in each system, some overlap in season of trapping and the gross extent of differences between the two systems justifies the comparison. Data from a shortgrass prairie ecosystem in Colorado (Pawnee National Grassland) (Stapp et al. 2009) show differences from the small mammal fauna found in our study site in mixed-grass prairie. Shortgrass prairie small mammal communities are dominated by (highest to lowest densities) O. leucogaster, I. tridecemlineatus, P. maniculatus, and other species (Stapp et al. 2009) (Fig. 1). In the mixed grass prairies of Lower Brule, this composition is essentially reversed with P. maniculatus making up the majority of animals trapped, and O. leucogaster being less frequently encountered (Fig. 1). The flea fauna of small mammals in shortgrass prairies has been found to overlap little with that of prairie dogs (Stapp et al. 2009). Similarly, in mixed-grass prairies, there is little overlap of fleas between prairie dogs and other small mammals. In both systems there is substantial overlap in flea faunas between deer mice and northern grasshopper mice. In contrast to the shortgrass system, densities of northern grasshopper mice do not seem sufficient for the maintenance of plague in our system, even given their small overlap in flea fauna with prairie dogs.
Host choice is an important ecological aspect of disease transmission. Parasites that feed multiple times on multiple individuals have greater potential for transmitting disease. Rodent fleas are primarily associated with either one, or a few closely related primary hosts (Marshall 1981) (where the flea reproduces), and fewer secondary hosts (hosts which co-occur with the primary host and are routinely exposed to the same fleas) (Krasnov 2008). It is important to note that even a rarely encountered or low-nutrient animal may serve as a transport host, moving fleas across the landscape, while not providing significant blood meals (Franklin et al. 2010). The possibility of significant flea sharing between deer mice and northern grasshopper mice as well as prairie dogs are potentially important interactions, which have not yet been fully investigated in mixed-grass prairies. While we tried to make these evaluations, our sample size did not allow for statistically meaningful evaluation, and more intensive trapping efforts for describing densities of northern grasshopper mice in mixed-grass prairies are necessary.
Formal comparisons between the two systems are difficult, since fleas were collected in different seasons and years. The difference in flea loads on small mammals between the two systems however is considerable, with Kraft and Stapp (2013) finding flea loads of 8.1 fleas per grasshopper mouse in shortgrass prairie in comparison to our finding of 3.11 fleas per grasshopper mouse in the mixed-grass ecosystem; Salkeld and Stapp (2008) found similar flea loads of 0.75 fleas per deer mouse, where we found 0.89 fleas per deer mouse. Comparison of our small rodent data to a study of fleas infesting black-tailed prairie dogs (C. ludovicianus) from the same area (Mize and Britten 2010) showed that flea sharing between small mammals and prairie dogs is minimal at Lower Brule. In 2009–2010, 99% of fleas taken from prairie dogs at Lower Brule were of two species, O. hirsuta or O. tuberculata; the remaining 1% was identified as unknown Oropsylla spp. Of the 1641 small mammal fleas collected, 1.77% (n = 29) of the fleas recovered were “prairie dog fleas” (O. tuberculata, O. hirsuta).
Our findings are consistent with those of Stapp et al. (2009), who reported 247 of 3485 fleas from northern grasshopper mice, 3 of 322 fleas from deer mice, and a single flea from a kangaroo rat (Dipodomys ordii) to be O. hirsuta or 5% (251/4716) of their total number of small mammal fleas. These fleas have the potential to serve as bridging vectors between prairie dogs and small mammals with the ability to move from host to host on occasion, particularly during epizootics when primary host numbers are down. However, their relatively small numbers in the mixed-grass system, in particular, call into question their ability to act as bridging vectors. However, under the right circumstances, predominantly rare flea sharing events could become more common and play a larger role in the circulation of Y. pestis. For example, if small mammals were serving as short-term reservoirs during enzootic periods, or shortly thereafter, and if the right conditions were to arise, levels of flea sharing could increase leading to potential spillovers into susceptible species such as prairie dogs helping to precipitate an epizootic phase. This type of scenario was proposed by Salkeld et al. (2010) as a means for minor components of the small mammal community associated with prairie dog colonies in shortgrass systems to accelerate and widen plague epizootics on the landscape. Our data from a mixed-grass system are consistent with this possibility.
Finally, it is important to note that although flea-borne plague is widely considered the primary mode of plague transmission, several other transmission routes may exist. Soil has been suggested as a reservoir (Karimi 1963, Malek et al. 2017) although it is not likely a major transmission route under natural conditions (Boegler et al. 2012), but could have implications on plague cycles under epizootic conditions. Direct transmission through consumption of infected carcasses, and/or inhalation of infectious droplets is potentially important, especially during epizootics (Richgels et al. 2016).
Within the small mammal community, A. wagneri is commonly encountered and widely distributed among host species. However A. wagneri has been shown to have low vector competency when compared with species such as O. montana (Eisen et al. 2008). Eisen et al. (2008) estimated that flea loads of this species would need to approach 68 fleas per deer mouse for enzootic maintenance due to its low vector efficiency. Our findings support those of Salkeld and Stapp (2008); Eisen et al. (2008); and Lang (2004) that deer mouse A. wagneri transmission alone, in mixed-grass prairies, probably is not capable of supporting enzootic plague. This is similar to what has been seen in shortgrass prairies of Colorado, and deer mouse populations in California (Lang 2004). In the mixed-grass prairie system, deer mice likely serve primarily as spillover hosts during plague epizootics. Similarly, the role of small mammals as a whole would seem to mirror that of deer mice.
Rocke et al. (2012) documented some resistance in prairie dogs from plague endemic regions. The possibility of resistant prairie dogs serving as a significant reservoir seems implausible due to the requirement of near terminal levels of infection for uptake of bacteria in feeding fleas. Additionally, results from Rocke at al. (2012) suggest that plague resistance is rare in naive prairie dog populations such as those studied here. While resistance may become a more prominent factor in maintaining Y. pestis in mixed-grass systems over time, other mechanisms such as the smoldering plague transmission hypothesis (Salkeld et al. 2016) and enzootic plague (persistence) (Biggins et al. 2010, Matchett et al. 2010) are probably currently more important. Potentially resistant mammals are nonetheless important in plague ecology in other systems, as they can play the role of maintenance host allowing fleas to feed, replicate and spread to susceptible individuals leading to potential spillover events. The reservoir of plague is likely augmented by alternative short-term reservoirs (O. leucogaster) (Stapp and Salkeld 2009) and routes of transmission (Richgels et al. 2016), such as through potential flea dispersal on carnivores (Gage 1994).
These findings, although based on a small number of sample locations, indicate that the composition of small mammals and their fleas differ between shortgrass and mixed-grass prairies, but the degree of flea overlap between prairie dogs and small mammals may not. Brinkerhoff et al. (2008) showed that in the presence of prairie dogs, the composition of the small mammal community may be influenced, and in turn the dominant flea fauna may shift leading to alternate disease paradigms. In this case, we have demonstrated that the second assumption of Graham et al. (2014) would not be met with any regularity, but further investigation in mixed-grass systems is warranted; especially with regard to the third of Graham's assumptions. The importance of resistant animals almost always seroconverting is that over time, the enzootic host population maintains at least some antibodies allowing for immune response upon further exposure and thereby limiting the disease (Jawetz and Meyer 1943). Furthermore, we do not know at the current time the directionality of feeding and the impact of small mammals on the cycles of sylvatic plague in mixed-grass systems. Inferences regarding the first and second assumptions of the enzootic host model could be strengthened through BMA, which would help to clarify the directionality of flea feeding. In the mixed-grass ecosystem, BMA would be necessary for more definitive analysis of the feeding patterns of potential bridging vectors both pre- and postepizootic. While Mize, and Britten (2010), found a 2% prevalence of Y. pestis from prairie dog fleas in the same area. Our data suggest that there is a low potential for Oropsylla spp. flea sharing between prairie dogs and other small mammals in mixed-grass systems. This study shows no evidence to counter the smoldering plague transmission hypothesis and is consistent with similar studies in shortgrass systems with respect to the presence of enzootic plague.
Footnotes
Acknowledgments
The authors acknowledge the contributions of S. Grassel, S. Maestas, L. Riley, and E. Mize for technical assistance in the field, assistance with figure formatting, and in the identification of fleas. S. Kempema, South Dakota Department of Game, Fish and Parks, was instrumental in the early development of this study. The authors thank the Lower Brule, Sioux tribe for access to land and wildlife and assistance in the acquisition of traps, and establishing trapping locations. This study was funded in part by federal funding through the State Wildlife Grant T-60-R1, Study #2471, administered through the U.S. Fish and Wildlife Service.
Author Disclosure Statement
We have no known conflicts of interest, nor do we have any commercial associations.