
Abstract
Ehrlichiosis and rickettsiosis are two common bacterial tick-borne diseases in the southeastern United States. Ehrlichiosis is caused by ehrlichiae transmitted by Amblyomma americanum and rickettsiosis is caused by rickettsiae transmitted by Amblyomma maculatum and Dermacentor variabilis. These ticks are common and have overlapping distributions in the region. The objective of this study was to identify Anaplasma, Ehrlichia, and Rickettsia species associated with questing ticks in a Rocky Mountain spotted fever (RMSF) hotspot, and identify habitats, time periods, and collection methods for collecting questing-infected ticks. Using vegetation drags and CO2-baited traps, ticks were collected six times (May–September 2012) from 100 sites (upland deciduous, bottomland deciduous, grassland, and coniferous habitats) in western Tennessee. Adult collections were screened for Anaplasma and Ehrlichia (simultaneous polymerase chain reaction [PCR]) and Rickettsia using genus-specific PCRs, and resulting positive amplicons were sequenced. Anaplasma and Ehrlichia were only identified within A. americanum (Ehrlichia ewingii, Ehrlichia chaffeensis, Panola Mountain Ehrlichia, and Anaplasma odocoilei sp. nov.); more Ehrlichia-infected A. americanum were collected at the end of June regardless of habitat and collection method. Rickettsia was identified in three tick species; “Candidatus Rickettsia amblyommii” from A. americanum, R. parkeri and R. andeanae from A. maculatum, and R. montanensis ( = montana) from D. variabilis. Overall, significantly more Rickettsia-infected ticks were identified as A. americanum and A. maculatum compared to D. variabilis; more infected-ticks were collected from sites May–July and with dragging. In this study, we report in the Tennessee RMSF hotspot the following: (1) Anaplasma and Ehrlichia are only found in A. americanum, (2) each tick species has its own Rickettsia species, (3) a majority of questing-infected ticks are collected May–July, (4) A. americanum and A. maculatum harbor pathogenic bacteria in western Tennessee, and (5) R. rickettsii remains unidentified.
Introduction
R
The most common southeastern TBD is RMSF, caused by R. rickettsii, which is often fatal if left untreated (McDade and Newhouse 1986, CDC 2013b). The primary vector is Dermacentor variabilis (Ricketts 1906), yet R. rickettsii has not been recovered from southern collections of D. variabilis (Stromdahl et al. 2011). While Amblyomma americanum harbors the currently nonpathogenic “Candidatus Rickettsia amblyommii” (Goddard and Norment 1986), it was recently suggested as an additional vector of R. rickettsii in North Carolina (Breitschwerdt et al. 2011) and in Kansas (Berrada et al. 2011). In addition Amblyomma maculatum has a role in many pathogen transmission cycles, including rickettsiosis (R. parkeri) in humans (Sumner et al. 2007, Goddard and Varela-Stokes 2009, Trout et al. 2010). Diagnostic tests for RMSF are often cross-reactive with other spotted fever group rickettsiae (SFGR) (Parola et al. 2005, Eremeeva et al. 2006a, Raoult and Parola 2008) and the presence of R. parkeri and “Ca. Rickettsia amblyommii” has raised further concerns about the true prevalence of RMSF infections in the United States (Paddock et al. 2004, Paddock 2005, Whitman et al. 2007, Apperson et al. 2008, Trout Fryxell et al. 2015b). Consequently, test results may incorrectly implicate RMSF instead of another SFGR (McCall et al. 2001). It is possible that cross-reactivity of R. parkeri, R. montanensis, and “Ca. R. amblyommii” is responsible for the increased incidence, declining case fatality, occurrence of multiple symptoms of spotted fevers, and increase in winter cases. Together, this has sparked several questions regarding whether a diagnostic error is occurring (e.g., false or cross-reactive positives), or circulating strains of R. rickettsii are becoming less virulent (Dantas-Torres 2007, Raoult and Parola 2008).
Ehrlichiosis is the second most common TBD in the southeast and is attributed to several different bacteria, including Ehrlichia chaffeensis, Ehrlichia ewingii, and Panola Mountain Ehrlichia (Holland et al. 1985, Anderson et al. 1991, Ewing et al. 1995, Buller et al. 1999, Loftis et al. 2008). The primary vector and amplifying reservoir of Ehrlichia species is A. americanum, a dominant tick species likely responsible for a majority of tick bites in the southeast (Anderson et al. 1993, Childs and Paddock 2003, Stromdahl and Hickling 2012). Within the United States, the number of ehrlichiosis cases has risen in the last several years (CDC 2013a). Complicating matters further, the clinical presentations of ehrlichiosis and rickettsiosis are similar.
Entomological investigations of tick vectors in Kentucky and Tennessee included screening field-collected specimens for Rickettsia or Ehrlichia (Cohen et al. 2010, Moncayo et al. 2010, Fritzen et al. 2011, Pagac et al. 2014, Harmon et al. 2015); rarely is the same tick screened for both organisms. Cohen et al. (2010) screened field-collected A. americanum, D. variabilis, Ixodes texanus (Banks), I. cookie Packard, I. scapularis (Say), and A. maculatum from 29 Tennessee counties for Ehrlichia and found only A. americanum to be infected (2.6% E. chaffeensis and 0.8% E. ewingii). Harmon et al. (2015) screened 303 A. americanum collected from a community experiencing an ehrlichiosis outbreak and found E. ewingii (5.3%), E. chaffeensis (2.0%), and Panola Mountain Ehrlichia (2.0%) in the questing population. Moncayo et al. (2010) screened host-collected and field-collected ticks from 49 counties for Rickettsia and found both “Ca. R. amblyommii” and R. montana in both D. variabilis and A. americanum, and R. cooleyi in I. scapularis. In Kentucky, Fritzen et al. (2011) screened A. americanum and D. variabilis and identified Borrelia lonestari, E. chaffeensis, and “Ca. R. amblyommii” in A. americanum, and R. montana and R. parkeri in D. variabilis. In that study they also identified four “Ca. R. amblyommii”-positive A. americanum coinfected with E. ewingii (2 specimens), E. chaffeensis (1 specimen), and B. lonestari (1 specimen). In addition, Pagac et al. (2014) screened 105 A. maculatum and 299 D. variabilis collected from military training sites in Kentucky and Tennessee and identified R. parkeri in A. maculatum (14.3%) and R. montanensis in D. variabilis (3.3%). None of the recovered Rickettsia sequences was identified as R. rickettsii (Moncayo et al. 2010, Fritzen et al. 2011, Pragac et al. 2014). While these studies provide a foundation for which pathogen and tick are in the area, they do not explain the increased incidence of TBDs in the region; epidemiological studies are needed.
As mentioned, in the United States, RMSF fatalities are decreasing, while diagnoses are increasing and severity or health outcomes are variable (CDC 2013b). Of interest are the RMSF cases in Tennessee. Tennessee accounts for only 2.4% of hospitalized RMSF cases in the United States; however, hospitalizations in the Tennessee Valley region result in 26% of the nation's RMSF fatalities with a majority of these cases occurring in central and western Tennessee, indicating that this area of the country is an RMSF hotspot (Chapman 2006, Adjemian et al. 2009). The objective of this study was (1) to identify Anaplasma, Ehrlichia, and Rickettsia species associated with questing ticks in a rickettsiosis hotspot in a complex environment representative of the southeastern landscape and (2) to identify habitats, time periods, and collection methods for collecting questing-infected ticks. In this study, we collected more than 9000 ticks representing three species in a single year, screened the adults for Anaplasma, Ehrlichia, and Rickettsia, and analyzed the collections by collection variables (month, habitat, and trapping method) to better understand the complex TBD associations in an RMSF hotspot.
Materials and Methods
Site selection
Ames Plantation (35.1153N, −89.2167W) (AMES), a University of Tennessee-managed research and education facility in western Tennessee, is located within the rickettsiosis hotspot identified by Adjemain et al. (2009) and has had a number of employees who live at the center who have been previously diagnosed with SFGR or similar tick-borne ailments. The facility is used for research (forestry, wildlife, ecological, and historical), education (many courses are taught at the facility), crop production (timber, livestock, field and row crops), and private recreation (sport hunting and host to the annual National Championship field trials) making the site frequented by numerous people. The tick sampling effort was spread across AMES using a superimposed 100-acre grid (0.40 km2). One hundred grids were randomly selected and a 100 m transect placed to represent a single habitat type: 25 native warm season grassland, 37 upland deciduous, 19 bottomland deciduous, and 19 coniferous sites (Fig. 1a). These habitats were designated based on the vegetation and soil type within each site, and additional details are reported in Trout Fryxell et al. (2015a).
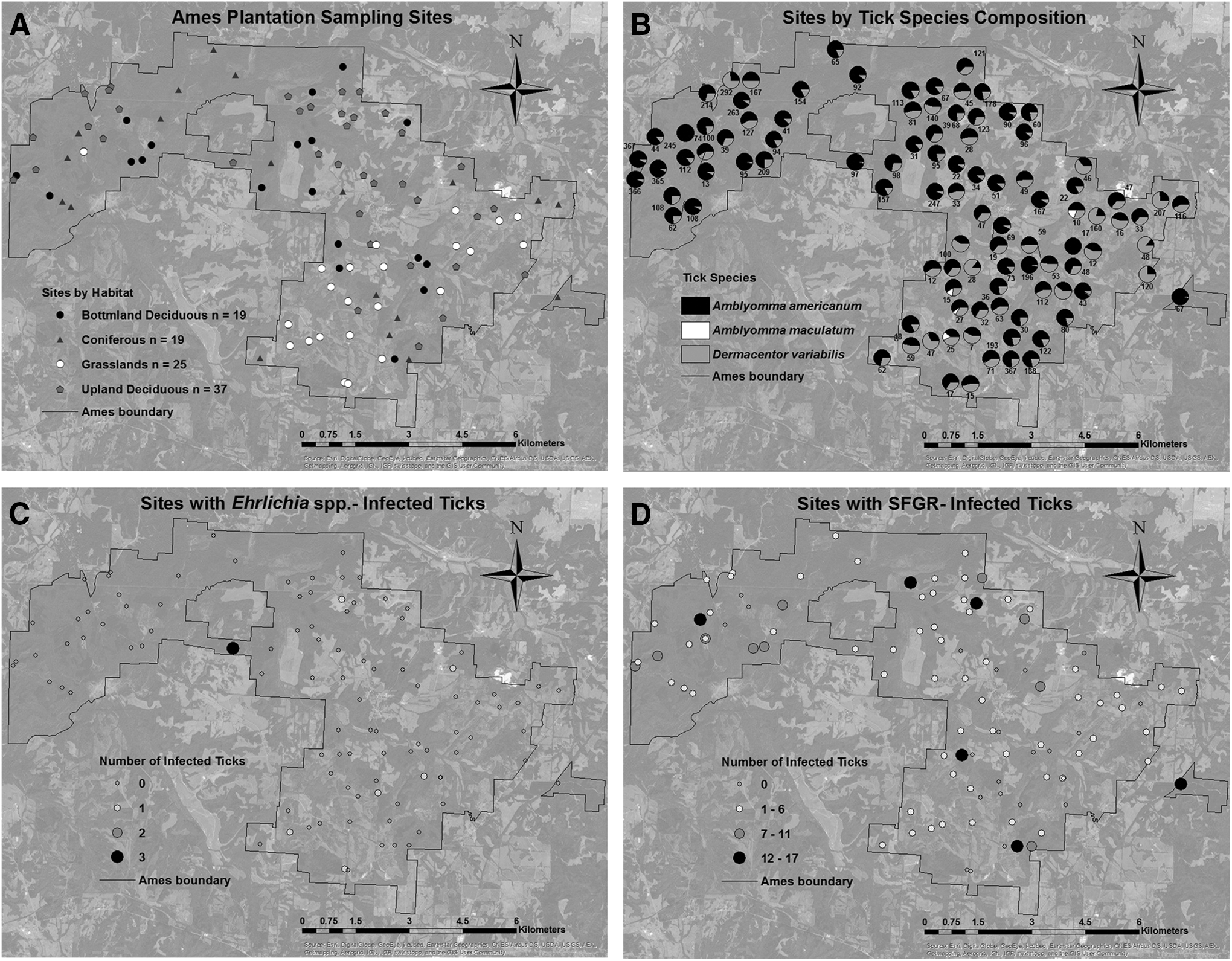
Ixodid ticks were collected from Ames Plantation (AMES) during 2012 from 100 sites representing native warm season grassland, upland deciduous, bottomland deciduous, and coniferous sites
Tick collections
Ticks were collected six times from May through September 2012, when a majority of Tennessee TBDs are diagnosed from each transect using vegetation drags and carbon dioxide (CO2)-baited traps (Falco and Fish 1992, Mays et al. 2016). Drags were constructed from a 1 m2 piece of corduroy cloth attached to a wooden dowel rod. The drag was inspected and ticks collected every 20 m along a 100 m transect. CO2-baited traps were made from a 1.9-L cylindrical blue cooler (Igloo Corporation, Katy, TX) with holes drilled around the circumference (Wilson et al. 1972). They were filled with ∼2.3 kg of dry ice and placed on a 1.37 m2 piece of light-colored duck cloth at the beginning of the 100 m transect and remained on site 12–16 h. Ticks attracted to the cloth and the cooler were removed in the field. CO2-baited traps were used at 10 sites in May, and then all 100 sites June through September. All ticks were collected and stored in trap/site-specific vials containing 80% ethanol, and specimens were identified to species, life stage, and sex (Cooley and Kohls 1944, Keirans and Litwak 1989, Keirans and Durden 1998).
Bacterial screening of adult ixodid ticks
Adult specimens were used in this study because they had the greatest chance of acquiring pathogens. Molecular methods were conducted in different facilities with equipment specific to each procedure to prevent contamination. Tick disruption procedures were identical to those reported in Hendricks (2013). Briefly, ticks were longitudinally bisected with a sterile razor blade and DNA was extracted using the Fermentas Gene Jet Genomic DNA Purification Kit and protocol (Thermo-Fisher Scientific, Pittsburgh, PA), which yielded 200 μL of genomic DNA; the total genomic DNA (tick and potential pathogen/host) was stored at −20°C until bacterial screening.
After DNA extraction, ticks were screened for Rickettsia species by PCR amplification of the 190 kDa protein antigen gene also referred to as the ompA gene hereafter (Eremeeva et al. 1994) and for Ehrlichia/Anaplasma species by a simultaneous nested PCR amplification of the groEL gene (Tabara et al. 2007; Takano et al. 2009), in separate master mixes using Maxima Hot Start Green PCR Master Mix (Thermo Scientific, Pittsburg, PA). Extracts were initially tested in pools: 5 μL of 10 individual extracted specimens were pooled into a single tube (∼50 μL pooled DNA), and 5 μL of that pooled DNA was used for primary screening for each pathogen in a 50 μL reaction (5 μL pooled DNA, 25 μL of Maxima Hot Start Green PCR Master Mix, 1 μL each of forward and reverse primers, and 18 μL nuclease-free water). If a pool tested positive, individual extracts from the positive pool were then screened for each pathogen in a 25 μL reaction (2 μL of individual tick DNA, 12 μL of Hot Start Master Mix, 0.5 μL each of the forward and reverse primers, and 10 μL of nuclease-free water). Primers and thermal cycler conditions are provided in Hendricks (2013). Positive controls consisted of tick DNA extractions previously testing positive for E. chaffeensis (99% identical to GenBank KJ907753L10917) or “Ca. R. amblyommii” (99% identical to GenBank HM446483). Positive PCR products were identified via gel electrophoresis (1.5% agarose gel: 1 × TAE buffer with ethidium bromide for 2 h at 120 V).
Selected positive amplicons were bidirectionally sequenced at the University of Tennessee Division of Biology Sequencing Facility to confirm positivity, to identify the bacterial species, and to classify the bacterial genotype (Trout et al. 2010). Due to the few groEL PCR positives, all groEL amplicons were sequenced. A random sample of Rickettsia PCR products were selected for sequencing because many of the ticks were positive. Resulting sequence reads were aligned in Sequencher 5.1 (Gene Codes Corporation Ann Arbor, MI) and compared to GenBank submissions via NCBI BLAST using the default conditions (other database, nonmodel organism optimized for highly similar sequences). All unique sequences were submitted to GenBank (KX158256-158278).
Identifying bacterial coinfections within adult ixodid ticks
In addition, to check for coinfections (more than one Ehrlichia, Anaplasma, or Rickettsia within a tick sample), all specimens PCR positive for either Ehrlichia or Rickettsia PCR were subjected to species-specific PCR testing as described in Gaines et al. (2014). Briefly, the six-plex test described in Gaines et al. was split into two three-plex reactions based on genus. Testing was performed in the ABI 7900 HT Fast Real-time PCR system (#4351405 Life Technologies; Thermo Fisher Scientific, Inc. Pittsburgh, PA). Reactions were performed with the TaqMan Real-time PCR master mix (Life Technologies) in a 25 μL reaction volume with 5 μL of DNA extract. Thermal cycling conditions, primers, and TaqMan probes (Life Technologies) were used as previously described (Gaines et al. 2014).
Epidemiological-collection associations (species, month, habitat, and trap)
To determine overall trends in the data set ANOVA, summary statistics, and pathogen abundance (overall and within each tick species) were calculated using JMP® Pro 11.1.1 (SAS Institute, Inc. Cary, NC) (α = 0.05). Pathogen prevalence was calculated by dividing the number of PCR-positive samples by the total number of ticks screened. Contingency tests (χ 2 ) were used to compare the frequencies of positive and negative ticks to determine if the tick species (3 species), collection period (3 species × 5 collection trips), habitat (3 species × 4 habitats), or trapping method (3 species × 2 trapping methods) was associated with the probability the tick would be PCR positive for either pathogen. For the contingency tests, Bonferroni's correction with 36 comparisons (species, collection period, habitat, and trapping method) was used to adjust calculated p values to prevent type I error caused by the various comparisons. Based on Bonferroni's correction, comparisons that yielded a p value ≤0.0014 were considered statistically significant. An additional analysis of logistic regression was calculated in SAS 9.4 TS1M3 for Windows 64 × (SAS Institute, Inc., Cary, NC) to determine the odds of collecting a positive tick for Ehrlichia or Rickettsia in a specific habitat and/or during a specific period. The trapping method variable was not included in the analysis because of the inherent differences in sampling methods (dragging for 5 min vs. baiting for 12+ h).
Results
Tick collections
Three tick species were collected totaling 9450 ticks: 6900 A. americanum (5974 nymphs and 926 adults), 2530 D. variabilis (5 nymphs and 2525 adults), and 20 adult A. maculatum (Fig. 1b). No I. scapularis or Rhipicephalus sanguineus specimens were collected. Twenty A. americanum adults were saved as voucher specimens (i.e., not included in the pathogen analyses). Ten of the D. variabilis lacked appropriate collection labels and were not included in the categorical analyses (collection month, trapping method, or habitat).
Briefly, A. americanum nymphs were collected significantly more in bottomland deciduous sites at the end of June (third sampling period) with a CO2 trap and rarely collected in upland deciduous sites or past the August sampling event (fourth sampling period) (F = 3.3867; df = 46, 1063; p < 0.0001). A. americanum adults were collected significantly more at coniferous sites during early May and June (first and second sampling period) with a CO2 trap and rarely collected past the August sampling event (fourth sampling period) (F = 7.6422; df = 46, 1063; p < 0.0001).
D. variabilis adults were collected significantly more from both deciduous sites during early June (second sampling period) with a CO2 trap than in grassland or coniferous sites later in the sampling season (F = 6.2883; df = 46, 1063; p < 0.0001). Amblyomma maculatum were not collected sufficiently for comparisons, but 16 of the 20 specimens were collected from grassland sites. Overall, a majority of ticks were collected from deciduous sites early in the sampling period (F = 1073.93; df = 46, 1063; p < 0.0001).
Bacterial screening of adult ixodid ticks
None of the D. variabilis or A. maculatum was groEL PCR positive. Ehrlichia or Anaplasma was identified in 17 (1.8%) of the 926 A. americanum (Table 1); 12 were 97–100% identical to E. ewingii (GenBank KJ907744, identified as Ehrlichia 1–4), 2 were 100% identical to Panola Mountain Ehrlichia (GenBank HQ658904, identified as Ehrlichia-5), 1 was 99% identical to E. chaffeensis (GenBank KJ907753, identified as Ehrlichia-6), and 2 were 99% identical to Anaplasma odocoilei sp. (GenBank JX876642, identified as Ehrlichia-7) (Table 1).
Rickettsia was identified in 353 (38.12%) of the 926 A. americanum, 92 (3.66%) of the 2515 D. variabilis, and 5 (25%) of the 20 A. maculatum. From the 45 sequenced A. americanum ompA amplicons, 5 different Rickettsia sequence variants were identified (Table 1), but all were identified as >99% identical to “Ca. R. amblyommii” (GenBank KT722804) (identified as Rickettsia 1–5, Table 1). Four of the A. maculatum Rickettsia-positives were 100% identical to R. parkeri (GenBank KF782320, identified as Rickettsia-6) and one was 100% identical to R. andeanae (GenBank KF179352, identified as Rickettsia-7). The 31 Rickettsia amplicons sequenced from D. variabilis were >99% identical to nonpathogenic R. montanensis ( = R. montana) (GenBank AY543681); 1 was identified as Rickettsia-8 (Table 1) and 30 were identified as Rickettsia-9 (Table 1).
Identifying bacterial coinfections within adult ixodid ticks
Eight of the A. americanum specimens were coinfected; six were coinfected with E. ewingii and “Ca. R. amblyommii,” two with Panola Mountain Ehrlichia and “Ca. R. amblyommii.” The number of coinfected ticks was not different from expected (X2 = 0.07; df = 1; p = 0.2463). No trend or pattern was evident in the coinfected ticks; they were collected from eight different sites, throughout the sampling period (2 specimens in May, 5 specimens in July, and 1 specimen in August), with both trap types (2 specimens with a drag and 6 with the CO2 trap). Five coinfected ticks were collected in upland deciduous sites and three were collected in coniferous sites. Coinfections of more than one Ehrlichia or Rickettsia were not identified using the novel real-time PCR assays described in Gaines et al. (2014). The multiplex data corroborated the sequencing results—identification of R. parkeri in A. maculatum and identification of “Ca. R. amblyommii” in A. americanum (Table 1).
Ehrlichia/Anaplasma-infected ixodid ticks, epidemiological-collection associations
Ehrlichia and Anaplasma were only amplified from 17 adult A. americanum ticks, in May (5/215), July (10/242), and August (2/78) (Table 2), indicating positive ticks were collected throughout the season; but more were collected from June 25 to July 6 (4.13% positive) compared to one sampling trip prior (May 28–June 8; 0 positive ticks out of 384 screened) (X2 = 14.9; df = 4; p = 0.005). Slightly but not significantly more Ehrlichia/Anaplasma-infected A. americanum were collected from grassland sites (4.69% positive) compared to bottomland deciduous sites (0.52% positive) (F = 2.1597; df = 3, 922; p = 0.00913) (Fig. 1c). The collection method (F = 0.7786; df = 1, 924; p = 0.3778) and sex of A. americanum (F = 0.4504; df = 2; p = 0.6375) were not significantly associated with Ehrlichia/Anaplasma infection. The logistic model for Ehrlichia indicated that period (χ2 = 0.0133; df = 4; p = 1.00), habitat (χ2 = 0.0088; df = 3; p = 0.9998), and their interactive effects (χ2 = 0.7275; df = 4; p = 1.00) were not significantly associated with encountering an Ehrlichia-infected A. americanum. Prevalence rates for each habitat and period for Ehrlichia/Anaplasma infection are presented in Table 2.
Rickettsia-infected ixodid ticks, epidemiological-collection associations
While each tick species was positive with its own Rickettsia, Rickettsia prevalence rates (no. positive/no. screened) were significantly greater in A. americanum (38.12% positive) and A. maculatum (25.0% positive) compared to D. variabilis (3.64% positive) (F = 450.9094; df = 2, 3468; p < 0.0001). Rickettsia-positive ticks were slightly but not significantly associated with habitat (Table 3, F = 3.5346; df = 3, 3457; p = 0.0142) such that collections of Rickettsia-infected ticks were more abundant in coniferous sites (15.84% positive) compared to grassland sites (8.78% positive) (Fig. 3d). Rickettsia infection did not vary by month within each species: A. americanum (χ2 = 5.79; df = 4; p = 0.215), A. maculatum (χ2 = 1.78; df = 4; p = 0.777), and D. variabilis (χ2 = 2.05; df = 4; p = 0.726); however, there was an overall significant collection effect such that more Rickettsia-infected ticks were collected earlier in the season (May through July) compared to later in the season (August through September) (F = 9.0641; df = 5, 3465; p < 0.0001). This was confirmed when the data set as a whole was analyzed in the logistic model, which indicated that only period (χ2 = 35.5775; df = 5; p < 0.0001) exerts significant effect on the odds of encountering a Rickettsia-infected tick; this was not true for habitat (χ2 = 0.6030; df = 3; p = 0.6030) or period × habitat (χ2 = 18.1107; df = 12; p = 0.1124). Odds ratios are presented in Table 4. Interestingly, significantly more Rickettsia-infected ticks were collected via dragging (19.53% positive) compared to the CO2 trap (11.29% positive) (F = 17.8556; df = 2, 3468; p < 0.0001; eight ticks were collected with an unknown method).
Bolded values are significant and period 6 was not included in the table; all periods versus period 6 had odds ratio >999 (<0.001 to >999).
Within species, there was no significant habitat, seasonal, or collection effect for A. americanum or A. maculatum (p > 0.05), but there was a slight habitat effect (which was not significant after the correction) within D. variabilis. Contingency tests indicated that slightly but not significantly more Rickettsia-positive D. variabilis were collected in upland deciduous (4.5% positive) and coniferous sites (4.4%) compared to grassland (1.3% positive) and bottomland deciduous sites (1.8% positive) (χ2 = 12.9; df = 3; p = 0.005). Contingency tests also indicated significantly more Rickettsia-positive A. americanum were collected in bottomland deciduous sites (41.4% positive) compared to grassland sites (32.8% positive) (X2 = 43.6; df = 3; p < 0.0001). The logistic model confirmed these findings. The logistic model indicated there was no significant risk of encountering Rickettsia-infected A. americanum by period (χ2 = 5.3804; df = 4; p = 0.2504), habitat (χ2 = 3.0887; df = 3; p = 0.3782), or their interactive effect (χ2 = 9.2827; df = 10; p = 0.5055). The logistic model also indicated there was no significant risk of encountering Rickettsia-infected A. maculatum by period (χ2 = 0.6562; df = 4; p = 0.9566), habitat (χ2 = 0.0043; df = 2; p = 0.9978), or their interactive effect (χ2 = 0.000; df = 1; p = 0.9980). The logistic model also indicated there was no significant risk of encountering Rickettsia-infected D. variabilis by period (χ2 = 0.0194; df = 5; p = 1.0), habitat (χ2 = 1.6862; df = 3; p = 0.64), or their interactive effect (χ2 = 13.1052; df = 12; p = 0.3614). Interestingly, contingency tests indicated CO2 trapping collected significantly fewer Rickettsia-infected D. variabilis in grassland sites (0.5% positive) compared to upland deciduous (4.7% positive) and coniferous (5.3% positive) sites (χ2 = 16.2; df = 3; p = 0.001).
Discussion
The confirmed identification of pathogenic Ehrlichia and Anaplasma in A. americanum and pathogenic Rickettsia in A. maculatum warrants additional research in the southeast and at this study site. This study also addressed bacterial associations with ticks collected concurrently from a site in a known RMSF hotspot environment. Adults were used in this study; however, investigations into nymph collections from different habitats and seasons may be more informative as they have had only one blood meal, are numerous (representing 63% of our collection, primarily A. americanum), and more difficult to detect making them more likely to stay on a host and transmit a pathogen (Falco et al. 1999). A majority of questing-infected ticks were collected May–July (period 1–3) regardless of habitats, during the same period when Tennesseans are diagnosed with rickettsiosis and ehrlichiosis. Additional investigations should be conducted just before determining when these infected ticks begin to quest for a host, and during this period to identify habitats, trapping methods, and evaluate methods for control.
Our results were similar to previous investigations as nonpathogenic Rickettsia, pathogenic R. parkeri, and several pathogenic Ehrlichia/Anaplasma were identified, while R. rickettsii was absent (Cohen et al. 2010, Moncayo et al. 2010, Harmon 2010, Fritzen et al. 2011, Stromdahl et al. 2011, Pagac et al. 2014). Our 38% Rickettsia-positive rate in A. americanum adults is important to consider as this species is most likely to bite humans in the southeastern United States and are typically faster and more aggressive than the other species collected at the site. While in the field working on this study, we encountered 167 ticks of which 158 were A. americanum, 7 were D. variabilis, and 2 were A. maculatum. We also encountered a number of larval ticks, but they were neither identified nor counted. All of the nonpathogenic Rickettsia identified within this study are cross-reactive with pathogenic R. rickettsii in diagnostic tests (McCall et al. 2001, Paddock et al. 2004, Paddock 2005, Paddock et al. 2005, Parola et al. 2005, Eremeeva et al. 2006a, b, Whitman et al. 2007, Apperson et al. 2008, Raoult and Parola 2008). Other reasons that R. rickettsii may not have been recovered include (1) R. rickettsii may have been in the positive nonsequenced A. americanum (n = 308), (2) Rickettsia coinfections such that the most abundant Rickettsia were identified with PCR and R. rickettsii was not amplified, (3) R. rickettsii may be in the substantially greater nymph population, which was not screened (5979 nymphs out of 9450 ticks, 63% of collection), or (4) that the site is not a true RMSF hotspot and the disease of concern is misdiagnosed as RMSF. In addition, the use of a nested PCR for Rickettsia may have amplified additional positive samples, some of which may have been R. rickettsii.
The few Ehrlichia/Anaplasma -infected ticks made it difficult to identify preferred habitats and trapping methods (Table 2); however, it was clear that A. americanum harbors several different Ehrlichia/Anaplasma species; collected with each trapping method and from each habitat, suggesting habitat might not play a pivotal role in Ehrlichia/Anaplasma transmission. In Tennessee, ehrlichiosis cases begin in May and steadily increase with a peak of cases in September (TNDOH 2013), which aligns with A. americanum adult and nymph activity demonstrated here. Previously, we identified the entire study site as a suitable habitat for A. americanum and speculated that host movement may be the best indicator for this species (Trout Fryxell et al. 2015a). White-tailed deer are primary hosts for A. americanum, move widely across a variety of habitats, and are known Ehrlichia and Anaplasma reservoirs; they are likely fundamental in transmission (Childs and Paddock 2003, Paddock and Yabsley 2007, Harmon et al. 2015). Because A. americanum was the only tick species infected with Ehrlichia/Anaplasma, studying the dispersal patterns along with movement patterns of their infected reservoirs should provide a mechanism for pathogen surveillance and vector control.
Both A. americanum and A. maculatum had significantly higher Rickettsia infection rates compared to D. variabilis, which allowed comparison of trapping success and prevalence between habitat types. Rickettsia infection was not specific to habitat, collection period, or trapping method; however, data indicate that ticks encountered earlier in the season (May, June, and July) have a greater chance of being Rickettsia infected regardless of habitat (Tables 3 and 4). In this study, our data indicate that the risk of encountering a Rickettsia-positive tick is nearly three times greater in periods 1 (May 8–16), 2 (May 28–June 8), and 3 (June 25–July 6), compared to periods 4–6. The risk for encountering a Rickettsia-positive tick in period 6 (September 21–29) was minimal compared to the other periods. In addition, dragging (19.5% positive) was significantly more effective at collecting Rickettsia-infected ticks compared to the CO2 trap (11.3% positive), indicating dragging is sufficient to obtain a representative of the questing and infected tick populations. Interestingly, the epidemiology of RMSF in human cases in Tennessee is bimodal with peaks in April and a second larger peak in July/August (TNDOH 2013), which corresponded with both adult and nymph A. americanum and A. maculatum adult abundance. Since A. maculatum are rare at the site, it would be beneficial to test the hypothesis that A. americanum nymphs are responsible for the second RMSF peak.
In this study, we found questing-infected ticks just before peak ehrlichiosis and RMSF human diagnoses in Tennessee (June–August), and this helps to explain the epidemiology of southern TBD cases. Tennessee TBD cases are likely acquired (person was likely bit by an infected tick) during May–July (or earlier) and a majority of cases are diagnosed 1–2 weeks after (indicating immune response and bacterial pathogenesis occur shortly after bite) (TNDOH 2013). The identification of habitats and temporal periods of questing-infected ticks provides public health professionals with information to prevent transmission. For example, signs can be posted outside of state parks with dense conifer vegetation in June to remind visitors to avoid the area or take extra precautions against tick bites (e.g., Deet on self, permethrin on clothing, tick checks). Similarly, this information can help physicians diagnose patients with a tick-borne illness if the patient indicates he/she/they visited a coniferous site in late June. Surveillance efforts can be concentrated during this period to identify specific land use areas with questing-infected ticks. Moreover, this information can also be used to develop and test management methods for the control of southern ticks as has been demonstrated for I. scapularis in the northeast (Ginsberg and Stafford 2005). Tick management methods include altering habitats conducive to infected ticks by applying acaricides, preventing hosts from entering the habitat, or any of the many other vegetation management options such as mowing, burning, and/or removing leaf litter (Ginsberg and Stafford 2005). Future research should focus on habitat/vector associations for each tick species, developing effective methods for collecting typically underrepresented tick species (such as A. maculatum), and creating surveillance programs for each species and pathogen.
Southeastern ehrlichiosis and rickettsiosis cases continue to be diagnosed and their vector distributions are expanding. In addition, the recent discovery of tick-borne viruses transmitted by A. americanum within the region, including Heartland virus (Savage et al. 2013) and Bourbon virus (Kosoy et al. 2015), warrants future studies to identify the true prevalence of these viruses along with their interactive effects with bacteria. Additional research needs to address the reasons for rickettsiosis cases in the southeast and identify the cause of increased fatalities in the Tennessee Valley compared to other regions with diagnoses but without high mortality (Adjemian et al. 2009). These reasons could be entomological (vector populations, frequencies, and/or densities), microbial (multiple Rickettsia coinfections, synergistic bacteria, and/or antagonistic bacteria), environmental (microbial, soil, host, abiotic, and/or landscape), and/or human mediated (medical doctors, patient behavior). Systematic studies examining biotic and abiotic parameters associated with southeastern ticks and TBDs should be conducted to develop a firm understanding of the interactions and dependencies between pathogens and hosts, the role of genetics and microbiomes, and how the ticks/pathogens/hosts partition or coexist in representative habitats. Field studies should be conducted at regional scales, allowing meaningful extrapolation to the southeastern ecological systems and human occupation patterns. Until R. rickettsii is identified in a tick vector or another pathogen is shown as a causative agent, the enigma surrounding TBDs in the south will remain (Dantas-Torres 2007, Cunha 2008, Raoult and Parola 2008, Adjemain et al. 2009).
Conclusions
Several bacteria causing ehrlichiosis were identified in A. americanum, and SFGR bacteria of unknown or varying pathogenicity were identified in A. americanum (“Ca. R. amblyommii”), A. maculatum (R. parkeri and R. andeanae), and D. variabilis (R. montanensis). These tick species are encountered commonly, have overlapping distributions, and positive specimens tend to be collected May–July, which correlate with ehrlichiosis and rickettsiosis cases in Tennessee. This period should be used as a time for increased awareness, self-protection, and avoidance by the public; surveillance by public health professionals and physicians, and habitat and host management by land managers. In this study, we report several pathogenic Ehrlichia/Anaplasma species amplified from A. americanum and R. parkeri amplified from A. maculatum, the absence of R. rickettsii, and identify May–July as peak tick transmission in Tennessee.
Footnotes
Acknowledgments
We thank Dr. Abelardo Moncayo with the Tennessee Department of Health for his contribution of E. chaffeensis and “Ca. Rickettsia amblyommii-”positive samples used as positive controls during the pathogen screening portion of our experiment. We also want to thank the reviewers for improving this manuscript, and Xiaocun Sun for help with additional statistical tests. We thank the board of trustees and the employees of Ames Plantation for accommodations, travel support, and so much more. In particular, we thank Larry Teague and James Morrow for their assistance with tick collection and vehicle extractions. Funding for this project was provided by the American Kennel Club Canine Health Foundation Acorn Grant (01894-A), Ames Foundation, and USDA Tennessee Hatch Project (TN00433). Fellowship support for B. Hendricks was provided by the University of Tennessee Institute of Agriculture's Department of Entomology and Plant Pathology, for S. Mays by the J. Wallace and Katie Dean Graduate Fellowship at the University of Tennessee, and for K. Pompo by Merial Veterinary Scholar Program at the University of Tennessee College of Veterinary Medicine.
Author Disclosure Statement
No competing financial interests exist.