
Abstract
West Nile virus (WNV) is a zoonotic single-strand RNA arbovirus (family Flaviviridae: Flavivirus), transmitted among avian hosts in enzootic cycles by a mosquito vector. The virus has a significant disease effect on humans and equines when it bridges into a cycle with various sequelae with epidemic potential. This study was carried out to identify the potential spectrum of WNV hosts in three geographic areas with climatologically distinct features: Malaysia, Qatar, and the United States of America (U.S.). Serum samples were collected from avian and mammal species suspected to be reservoirs for the virus at these areas in a cross-sectional epidemiologic study. The samples were tested for the presence of antibodies against the virus using an enzyme-linked immunosorbent assay. Data on putative risk factors were also collected and analyzed for significance of association with seropositivity using the logistic regression analysis. Among the tested avian and mammalian species, raccoons had the highest seroconversion rate (54%) followed by crows (30%), horses (27%), camels (10%), other avian species (7%), and canine species (3%). It was almost twice as likely to detect seroconversion among these mammalian and avian species in the fall in comparison to other seasons of the year. Only mammalian and avian species and seasons of the year were significantly associated with the likelihood of seroconversion to WNV when we controlled for other factors in the multivariate analysis. Our data from the U.S. showed that raccoons and camels are susceptible to infection by the virus and may play a role in the perpetuation of endemic foci for the disease.
Introduction
A
WNV is maintained in nature in a cycle between birds and mosquitoes (Blitvich, 2008). Infected mosquitoes do feed on other vertebrate animals leading to the spread of the WNV to a larger spectrum of hosts (Rossi et al. 2010). Some of these hosts could be exposed to the virus and yet not exhibit any clinical signs (Reisen et al. 2004). Transmission of the virus among hosts without an involvement of a vector is rare. Several factors are known to play a role in the transmission and perpetuation of WNV in ecological niches where the agent and environmental factors have received great attention, but information on the possible animal reservoir is lagging behind (Rossi et al. 2010, Tran et al. 2014, Chancey et al. 2015, Day et al. 2015). These niches evolve as a consequence of the presence of the appropriate hosts and vectors in an ecologically conducive environment. Identifying these factors will be better defined by examining different ecological niches and it was the focus of this research to depict the wide spectrum of WNV reservoirs, a cross-species, and geography that will illustrate the varying risk factors of the expanding nature of WNV.
Materials and Methods
Target and study population
The target population consisted of mammals and avians of different species located in three geographically and climatologically distinct areas: Malaysia, Qatar, and the U.S.
Malaysia
Blood samples were collected from captive birds from four different locations in Selangor, Malaysia, during the months of November and December. The first set of samples from birds belonged to a farmer who kept them as biological controls in the rice paddy fields at Tanjung Kajang, Selangor. The second set was from the teaching hospital at the Faculty of Veterinary Medicine, Universiti Putra Malaysia, where the birds were admitted for different conditions. The other two sets of samples were from the National Zoo and Sunway's Wildlife Park, respectively, in Kuala Lumpur, where the birds are kept for recreational activities in big net enclosures.
Qatar
All the mammalian samples in the State of Qatar were from horses that resided in different equine operations throughout the country. The horses were of different breeds and included thoroughbred, Arabian, and mixed breeds.
United States
Bovine and caprine
Blood samples from the bovines were obtained from dairy operations located in the New York City Watershed (NYCW). Samples from the bovine tail veins were obtained using a 20-gauge vacutainer® needle and a 5-mL vacutainer blood collection tube (BD; Becton Dickinson and Company, Franklin Lakes, NJ). Samples from caprine species were collected from goat operations in the NYCW and were obtained from the jugular veins using vacutainer tubes. Camel samples were acquired from samples submitted to the Animal Health Diagnostic Laboratory for routine vitamin E level determinations. These camels are kept as pet animals by private owners.
Raccoons
Raccoons were captured and blood sample collection was performed as previously described (Boulanger et al. 2008). The raccoons sampled in this study represent a population captured in a rabies vaccine study on Long Island, NY. Briefly, Tomahawk box traps (Tomahawk Live Trap Co., Tomahawk, Wisconsin) baited with Fur King (Blackie's Blend, Glenmont, Ohio), a commercial raccoon sweet paste, were used to trap raccoons during September–October 2003–2005. Trap locations were selected to maximize capture rate and to avoid disturbance from people, pets, and other wildlife. If no raccoons were captured at a trap site within three nights, the two traps were moved to another location. The captured raccoons were sedated and blood samples (10 mL) were collected from femoral blood vessels in vacutainer tubes. Sex, relative age, and weight of each raccoon were recorded. Raccoons were released at the site of capture after a recovery period. This research conforms to the requirements of Cornell University's Institutional Animal Care and Use Committee (Protocol No. 95-79-01).
Detection of antibodies to WNV
The presence of antibodies to WNV was detected using an enzyme-linked immunosorbent assay (ELISA) (ID Screen West Nile Competition, IDVet, Grabels France). The test was performed according to the manufacturer's protocol in all three countries due to its multispecies capability. Briefly, using the microplates provided in the ELISA kit, 50 μL of dilution buffer 2 was added to each microwell that had been coated with purified antigens. Following the addition of dilution buffer, 50 μL of both negative and positive controls were added to the specified wells. Finally, 50 μL of tested sample was added to the remaining wells. This preparation process was continued for all of the samples. Following the addition of the buffer, negative/positive controls, and tested samples, fully loaded microplates were incubated at 21°C for 90 min. An antibody peroxidase conjugate was prepared by dilution, concentrated conjugate 10 × to conjugate 1 × by diluting the concentrated conjugate to 1/10 of its original concentration in dilution buffer 2. After the 90-min interval, the wells were emptied and washed three times with ∼300 μL of the wash solution, and 100 μL of the conjugate (1 × ) was added to each well. The microplate was incubated at 21°C for an additional 30 min. The wells were once again washed three times with 300 μL of the wash solution to eliminate excess conjugate. With the excess conjugate eliminated, 100 μL of the substrate solution was added to each well and incubated for 15 min at 21°C in the dark. Following the heating of each well, 100 μL of the stop solution was added to stop the reaction. The absence of antibodies resulted in the color transition from blue to yellow after the addition of the stop solution. Conversely, in the presence of antibodies, no coloration appears. Finally, microplates were read at 450 nm to determine the optical density (OD) of each sample. The results were interpreted as positive or negative based on the ratio of the sample's OD to the control negative OD.
Statistical analyses
The mammal and avian species specific prevalence was computed as the proportion that tested positive to WNV among all samples collected from all the respective species (Table 1). The likelihood of seroconversions for each species in the study was compared to that of the raccoons (as a reference category) and was computed using the odds ratio (OR) (Table 2). The significance of association between each of the putative risk factors (species of the mammals and avians, season of sample collection) was evaluated using the univariate logistic regression. Factors that were significantly associated with the likelihood of seroconversion to WNV in the univariate analyses were jointly evaluated in multivariate logistic regression analyses (Table 3). The rationale was to assess the significance of association of each factor while simultaneously controlling for the presence of other factors. The magnitude of the respective association was measured using the OR.
Number of samples.
CI, confidence interval.
All statistical analyses were performed using the SPSS v23 (IBM Statistical Software, White Plain, New York) and significance was evaluated at type I error ≤0.05.
Results
A total of 560 samples collected from different mammals and avians inhabiting Malaysia (69), Qatar (161), and the U.S. (330) were analyzed in this study. The seroconversion proportion among the samples from Malaysia was 7% (95% confidence interval [CI] = 1–14), Qatar 27% (95% CI = 20–34), and the U.S. was 25% (95% CI = 21–30). The seroconversion among samples collected from Qatar or the U.S. was significantly higher than that in samples collect in Malaysia. There was no significant difference in the seroconversion proportion between samples collected in Qatar and the U.S.
The samples were collected from seven different mammal and avian species in those countries: canine, bovine, caprine, equine, camelid, avian, and procyonid (Table 1). Samples collected from raccoons had the highest seroconversion prevalence followed by crows, horses, and camels, 54, 30, 27, and 10%, respectively. Samples collected from canine species in the U.S. and avian species from Malaysia had low seroconversion prevalence, 3% and 7%, respectively (Table 1). None of the samples collected from caprine and bovine species, from the U.S., tested positive.
All the samples were collected from animals located on one of the following ecological landscapes: forest, rural, suburban, urban, and agricultural (Fig. 1). None of the samples from agricultural areas was positive (95% CI = 0–0.38). We compared the mean difference among these four landscape areas using the analysis of variance and the Tukey's posthoc test. Animals located at the suburban areas had a significantly higher prevalence compared to animals on other landscape areas. There was no significant difference in the prevalence of seroconversion among animals located on the other three areas.
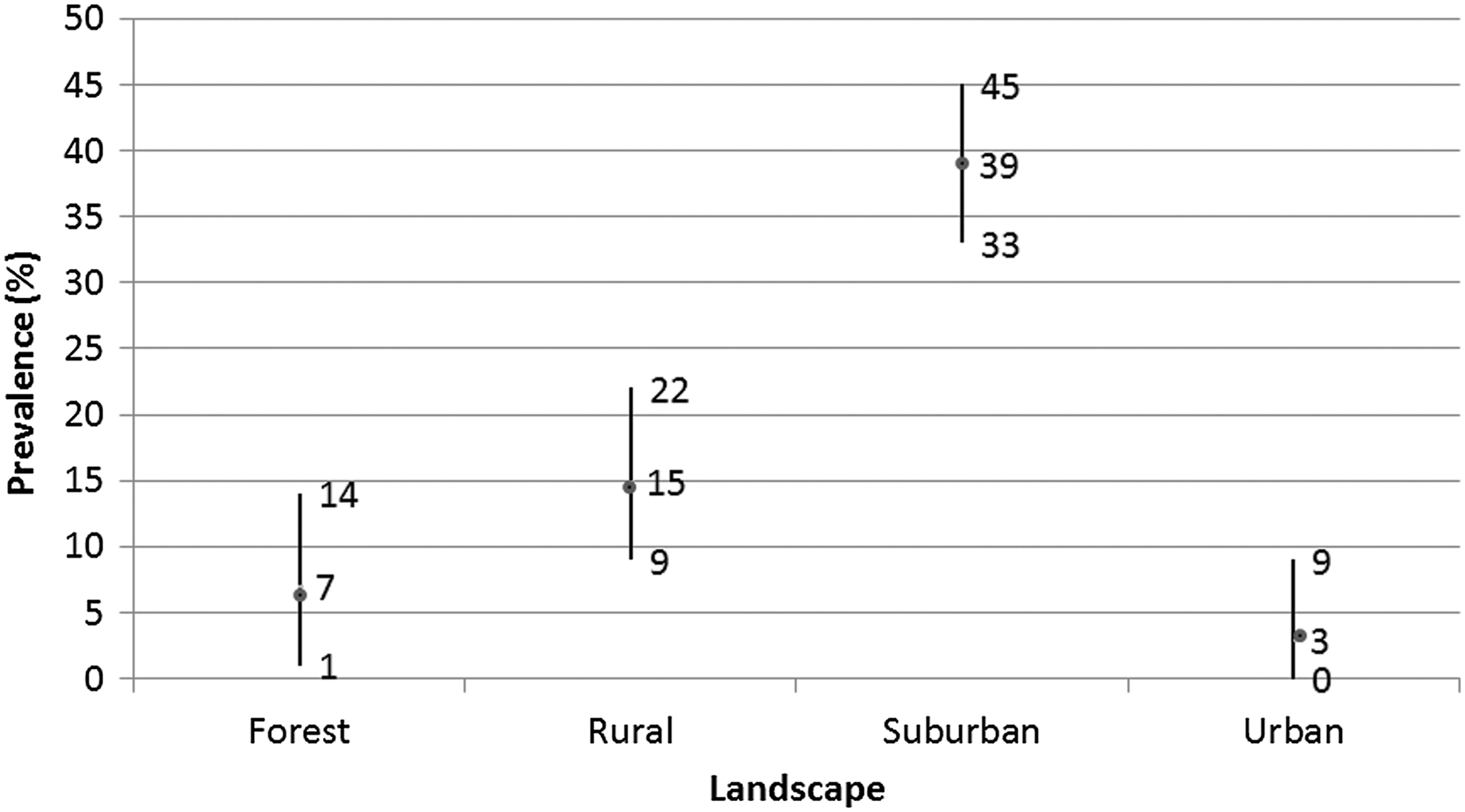
The distribution of the prevalence of seroconversion to WNV by land use among the study population and the 95% confidence interval. WNV, West Nile virus.
Table 2 shows the results of the univariate logistic regression. There was a significant association between the species of the mammal or avian and the likelihood of seroconversion to WNV. All the species evaluated had significantly low likelihood of seroconversion to WNV in comparison to raccoons. Mammals and avians sampled during the fall (late August to early October) season of the year were almost twice as likely to seroconvert to WNV in comparison to the other seasons combined (OR = 1.83, 95% CI [1.2–2.78]).
There was significant association between the land use and the likelihood of seroconversion to WNV in the univariate analysis (Table 2). Animals sampled in suburban areas were more than 20 times likely to seroconvert to WNV in comparison to animals sampled in urban areas (OR = 20) (Table 2). Although the likelihood of seroconversion to WNV among animals sampled in forest or rural areas was high in comparison to the urban areas, the OR was not significant in comparison to the urban areas (Table 2).
We also examined the relationship between the age of the animal and the likelihood of seroconversion to WNV (Table 2). Data on the age of the animal were available from canine (n = 33), crows (n = 64), horses (n = 101), and camels (n = 32) of the animals in the study. Although there was a trend of increased probability of seroconversion to WNV as shown in Figure 2, the trend was not significant.
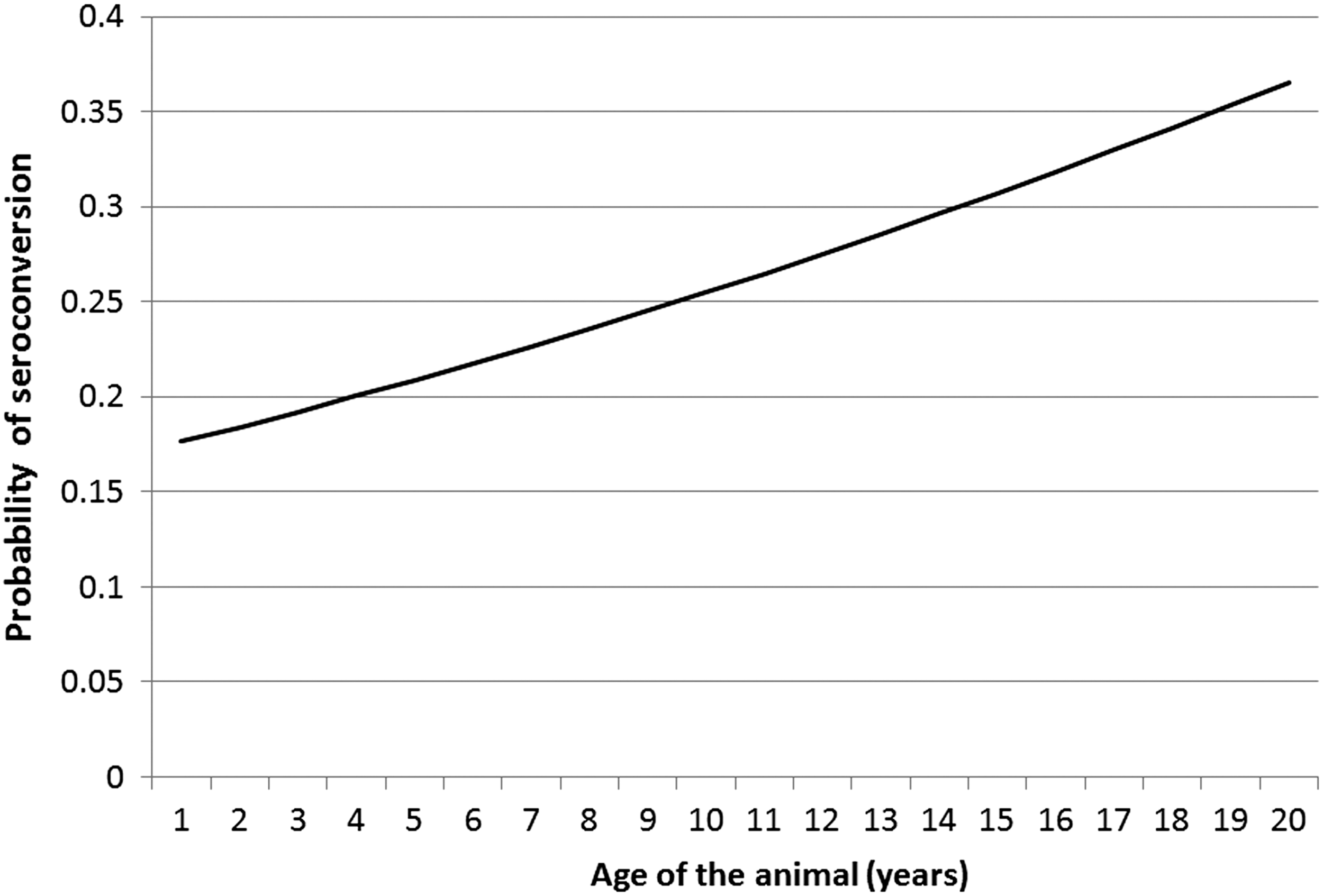
The probability of seroconversion to WNV, given the age of the animal as computed from the logistic regression analysis.
Among the three geographical areas that are represented by the respective countries, Malaysia had the least prevalence. The likelihood of seroconversion among avians sampled in Malaysia was significantly lower than mammals and avians sampled in the U.S. (OR = 0.15, 95% CI [0.06–0.39]). Although the likelihood of seroconversion to WNV among mammals sampled in a dry area (Qatar) was less in comparison to mammals and avians sampled in the U.S. (OR = 0.7, 95% CI [0.45–1.09]), the difference was not significant.
In the multivariate analyses, only the species of the animal and the season of the years were significantly associated with the likelihood of seroconversion to the WNV (Table 3). Among the different species of animals, the pattern of risk remained the same as in the univariate analysis. All species investigated in the study had significantly lower likelihood of seroconversion to WNV in comparison to raccoons. Raccoons were 33 times more likely to seroconvert to WNV in comparison to the avian species in the study (OR = inverse of 0.03). In contrast, raccoons were three times more likely to seroconvert to WNV in comparison to crows or horses (inverse of the respective OR) (Table 3).
Discussion
The long-term goal of our investigative research is to shed light on the factors that play a role in the spread of the virus to new ecological areas and, within these areas, the factors that play a role in the transformation of epidemic foci onto endemic niches. In the context of this objective, we opted to select these three countries that represent distinct ecologic zones, Malaysia (tropical), Qatar (subtropical desert), and the U.S. (temperate), to compare the exposure among a variety of potential mammal and avian reservoirs (Chancey et al. 2015).
Although exposure to the virus had been reported before in the U.S., no evidence had been cited in either Malaysia or in Qatar among animals (Chancey et al. 2015). In Malaysia, evidence of seroconversion in humans to the WNV has been reported among Orang Asli (native population) in the forest area (Marlina et al. 2014). None of the avians that were sampled had any clinical signs consistent with the diagnosis of WNV disease. All the samples collected in Malaysia were from avian species since data to date have emphasized the role of birds as the amplifying reservoir for the virus.
Mammal and avian species sampled in the U.S. included raccoons, crows, dogs, bovine, goats, and camels. The rationale for including these species stems from the fact that the WNV disease is endemic in the U.S. and the prevalence of exposure among the avian species and horses is known (DeCarlo et al. 2011, Gray and Webb, 2014).
The role of mesopredators as possible hosts of WNV has not been determined (Blitvich et al. 2009). The role of raccoons in the perpetuation of WNV cannot be concretely judged since there are no laboratory studies that quantitate their peak viremia level B. The viremia level needed for transmission is 105 pfu mL−1 (Turell et al. 2000, Sardelis et al. 2001, Blitvich, 2008). The prevalence of seroconversion in this study is consistent with the previous literature (Dietrich et al. 2005, Root et al. 2005, Bentler et al. 2007).
All the samples collected from bovine and caprine species had zero prevalence of seroconversion to WNV. This is similar to what has been reported in Northwest Senegal (Davoust et al. 2016).
In situations where the numerator of the proportion was zero, we used the approximation formula for the 95% CI for no event (Hanley and Lippman-Hand, 1983). The upper limit for the 95% CI for zero events calculates the maximum risk of infection under the normal distribution hypothesis. The maximum risk of seroconversion to exposure to WNV in dairy cattle and in goats in this study was 5% and 11%, respectively.
No data on the seroconversion to WNV were reported in camels sampled in similar ecological zones in North America; however, samples collected from camels in the Astrakhan region in Europe reported a prevalence of seroconversion to WNV at a range of 5–41% (Vasil'ev et al. 2005). It is worth noting that in the study in Europe the serologic test used was the hemagglutination inhibition test.
Because of the differences in the climate among the three countries, we have classified the time of sampling into two seasons, fall (late August to early October) and other times of the year. Our findings in relation to season are consistent with the studies in similar Northeastern U.S. areas reporting that seroconversion and isolation of WNV are highest in early fall among mammals, humans, avians, and mosquitoes (Roehrig et al. 2002, Andreadis et al. 2004).
Conclusions
We are surprised to observe high seroconversion to WNV among samples collected in Qatar. It is worth noting that all the samples were collected from horses in that country and there had been no report on evidence of exposure or isolation of WNV in Qatar. None of the horses had a history of vaccination. However, with an almost one-third exposure rate, a WNV equine vaccination protocol should be contemplated in this region. Vaccination can reduce the risk of death by 44% (Blitvich 2008).
Footnotes
Acknowledgments
This research is partially funded by a grant from the Qatar Foundation (NPRP no. 8-1854-04-027) and from the Agricultural Research Station, Cornell University. The authors express their appreciation to IDVet Innovative Diagnostic, Grabels, France, for providing the ELISA kits as well as the granting organizations.
Author Disclosure Statement
No competing financial interests exist.