
Abstract
Anaplasma species are obligate intracellular rickettsial pathogens that cause great economic loss to the animal industry. Few studies on Anaplasma infections in Mongolian livestock have been conducted. This study examined the prevalence of Anaplasma marginale, Anaplasma ovis, Anaplasma phagocytophilum, and Anaplasma bovis by polymerase chain reaction assay in 928 blood samples collected from native cattle and dairy cattle (Bos taurus), yaks (Bos grunniens), sheep (Ovis aries), and goats (Capra aegagrus hircus) in four provinces of Ulaanbaatar city in Mongolia. We genetically characterized positive samples through sequencing analysis based on the heat-shock protein groEL, major surface protein 4 (msp4), and 16S rRNA genes. Only A. ovis was detected in Mongolian livestock (cattle, yaks, sheep, and goats), with 413 animals (44.5%) positive for groEL and 308 animals (33.2%) positive for msp4 genes. In the phylogenetic tree, we separated A. ovis sequences into two distinct clusters based on the groEL gene. One cluster comprised sequences derived mainly from sheep and goats, which was similar to that in A. ovis isolates from other countries. The other divergent cluster comprised sequences derived from cattle and yaks and appeared to be newly branched from that in previously published single isolates in Mongolian cattle. In addition, the msp4 gene of A. ovis using same and different samples with groEL gene of the pathogen demonstrated that all sequences derived from all animal species, except for three sequences derived from cattle and yak, were clustered together, and were identical or similar to those in isolates from other countries. We used 16S rRNA gene sequences to investigate the genetically divergent A. ovis and identified high homology of 99.3–100%. However, the sequences derived from cattle did not match those derived from sheep and goats. The results of this study on the prevalence and molecular characterization of A. ovis in Mongolian livestock can facilitate the control of infectious diseases in livestock.
Background
T
A. marginale is an obligate intraerythrocytic pathogen that primarily affects cattle and causes bovine anaplasmosis (Kocan et al. 2010). Bovine anaplasmosis causes considerable economic loss to cattle farmers residing in tropical or subtropical regions (Aubry and Geale 2011) as it can be transmitted mechanically (by biting flies) and biologically (by ticks). Approximately 20 species of ticks have been implicated as vectors of this pathogen (Kocan et al. 2003).
A. ovis is another intraerythrocytic pathogen that primarily infects sheep or goats and causes ovine anaplasmosis (Renneker et al. 2013). It has been reported in many regions in the world (Ndung'u et al. 1995, de la Fuente et al. 2007, Renneker et al. 2013, and Yang et al. 2015). In China, a neighboring country to Mongolia, three biological vectors of A. ovis have been identified: Dermacentor nuttalli, Hyalomma asiaticum, and Rhipicephalus pumilio (Yin and Luo 2007).
A. phagocytophilum is the most problematic Anaplasma species because it is pathogenic to both humans and animals (Woldehiwet 2010). It replicates mainly in the granulocytes of its host and causes tick-borne fever in ruminants and granulocytic anaplasmosis in humans, dogs, and horses (Rikihisa 2011). The occurrence of A. phagocytophilum has been reported in many countries, most commonly in the Northern hemisphere, and its distribution is associated with several members of the Ixodes genus (de la Fuente et al. 2005, Cao et al. 2006, Kawahara et al. 2006, Carrade et al. 2009, Sainz et al. 2015).
A. bovis infects monocytes and causes monocytic anaplasmosis primarily in cattle (Rymaszewska and Grenda 2008) but also in other species, including goats (Liu et al. 2012), dogs (Sakamoto et al. 2010), and rabbits (Goethert and Telford 2003). It is commonly observed in South Africa (Harrison et al. 2013), Tunisia (Belkahia et al. 2015), India (Nair et al. 2013), China (Liu et al. 2012), Korea (Doan et al. 2013), and Japan (Ooshiro et al. 2008). Known vectors of the pathogen include Haemaphysalis longicornis (Doan et al. 2013), H. punctata (Palomar et al. 2015), and H. leporispalustris (Goethert and Telford 2003).
To date, three Anaplasma species have been detected in Mongolian livestock: A. marginale in cattle (Ybañez et al. 2013); A. ovis in sheep, goats, cattle (Sophia et al. 2012), and reindeer (Haigh et al. 2008); and A. phagocytophilum in sheep, goats, cattle, and horses (Sophia et al. 2012). These studies sampled animals from only few geographic regions, specifically the Khuvsgul or Khentii provinces; therefore, genetic information for characterizing these species is still limited. An Anaplasma species closely related to A. ovis (95.0% identity) based on the groEL gene was recently identified in Mongolian cattle (Ybañez et al. 2013); however, more studies are needed to characterize this organism. Although we have little information on the prevalence of Anaplasma infection in Mongolian livestock, pathogens, such as A. ovis, are known to cause severe disease in reindeer in the Khuvsgul Province (Haigh et al. 2008). Thus, additional studies are needed to examine the prevalence and molecular identity of Anaplasma species in various species of Mongolian livestock in different geographic regions.
In this study, we investigated the presence of A. marginale, A. bovis, A. ovis, and A. phagocytophilum in cattle, yak, sheep, and goats sampled from five different regions in Mongolia and determined the molecular characterization of A. ovis based on groEL, msp4, and 16S rRNA genes.
Materials and Methods
Study areas and samples
In total, 928 whole blood samples were collected from randomly selected free-range Mongolian livestock between June 2014 and July 2014 using BD K3 EDTA vacutainer blood collection tubes (Becton, Dickinson and Company). Of these, 517 blood samples were collected from bovine species: 100 samples from yaks (Bos grunniens) in Bulgan soum of Arkhangai Province or Songinokhairkhan district of Ulaanbaatar city, 117 from native cattle (Bos taurus) in Tsenkher soum of Arkhangai Province or Lun soum of Tuv Province, and 300 from dairy cattle (Holstein, Simmental, and Alatau) (B. taurus) in Bornuur soum of Tuv Province or Songinokhairkhan district of Ulaanbaatar city.
In addition, 411 blood samples were collected from sheep (Ovis aries) and goats (Capra aegagrus hircus). Of these, 211 samples were collected from sheep in Lun soum of Tuv Province or Tsenkher soum of Arkhangai Province and 200 samples from goats in Lun soum of Tuv Province (Fig. 1). All animals appeared healthy with no visible clinical symptoms during the sampling period. In addition, no tick was detected on any sampled animal.

Map of sampling regions in Mongolia.
DNA extraction
Soon after collecting the blood samples, genomic DNA was extracted using a Genomic DNA Purification Kit (Promega Corporation), according to the manufacturer's instructions. Total DNA was eluted with 100 μL conservation buffer and stored at −30°C until further use.
Diagnostic polymerase chain reaction assays for detecting Anaplasma species
A. phagocytophilum and A. bovis were screened using nested polymerase chain reaction (PCR) assay targeting the 16S rRNA gene (Kawahara et al. 2006), whereas A. marginale was screened using nested PCR assays targeting 16S rRNA (Weisburg et al. 1991, Ybañez et al. 2013), major surface protein 1 (msp1b) (Molad et al. 2006), and msp5 genes (Ybañez et al. 2013). In contrast, A. ovis was detected using single-step PCR targeting msp4 (de la Fuente et al. 2007) and nested PCR targeting 16S rRNA (Barlough et al. 1997) or heat-shock protein (groEL) (Ybañez et al. 2013) genes. The primers used in this study are shown in Table 1. In brief, 1.5 μL extracted sample DNA, 1.5 μL positive control DNA, or 1.5 μL double distilled water (DDW) as negative control was added to 28.5 μL reaction mixture comprising 3 μL 10 × buffer (Takara Bio, Inc., Japan), 2.4 μL dNTPs (Takara Bio, Inc., Japan), 1 μL (10 μM concentration) forward and reverse primers (Hokkaido System Science Company, Japan), 0.1 μL DNA Taq polymerase (Takara Bio, Inc., Japan), and 21 μL DDW. PCR amplification was performed under the following thermal cycle condition: initial denaturation at 94°C for 5 min, followed by 40 cycles of denaturation at 94°C for 30 s, annealing at each optimal temperature for 30 s, extension at 72°C for 1.5 min, and a final extension at 72°C for 7 min using the GeneAmp PCR System 9700 (Applied Biosystems). The amplified PCR products were separated by electrophoresis on 2% agarose gel and were observed under ultraviolet light.
Degenerate primer: R = A or G.
DNA cloning and sequencing
In total, 24 positive samples for each of the 3 genes (total 72 PCR products) were extracted using FastGene gel/PCR Extraction Kit (Nippon Genetics Company, Japan). The extracted PCR products were ligated into the pGEM-T Easy Vector (Promega Corporation), and the plasmid was transformed into Escherichia coli strain DH5α (Takara Bio, Inc., Company, Japan), plated on a Luria–Bertani (LB) agar (Thermo Fisher Scientific Corporation), and cultured in LB broth (Thermo Fisher Scientific Corporation). The plasmid DNAs were extracted from the positive clones in the LB culture using FastGene Plasmid Mini Kit (Nippon Genetics Company, Japan). Sequence amplification of the plasmids was performed using GeneAmp PCR System 9700 (Thermo Fisher Scientific Corporation).
The quality of the plasmid preparation for each pathogen gene was assessed using NanoDrop 8000 analytical equipment (Thermo Fisher Scientific Corporation), and pathogen sequencing analysis was performed using a CEQ8000 DNA analysis system (Beckman Coulter, Inc.).
Phylogenetic and homology analyses
The sequences were analyzed using Bio-Edit software (Hall 1999) and the basic local alignment search tool. Phylogenetic trees were constructed by MEGA 7 software (Tamura et al. 2007) using the neighbor-joining (NJ) method (Saitou and Nei 1987) and the maximum-likelihood method (Goldman 1990) with 1000 bootstrap replications. Consequently, the NJ trees were introduced as representatives in this study because of the lack of differences.
Results
In total, 928 blood samples collected from Mongolian livestock were screened for Anaplasma species using single-step or nested PCR assays. A. ovis was detected in the samples of all animal species (sheep, goats, cattle, and yaks) from all five sampling regions (Songinokhairkhan district of Ulaanbaatar city, Lun or Bornuur soum of Tuv Province, and Tsenkher or Bulgan soum of Arkhangai Province). A. phagocytophilum, A. bovis, and A. marginale were not detected. The overall livestock infection rates of A. ovis targeting groEL and msp4 genes were 44.5% and 33.2%, respectively. Pathogen prevalence in different hosts and different sampling regions is detailed in Table 2.
The sequences that were divergent compared with the sequences from other countries.
The sequences that were similar with the sequences from other countries.
In the sequencing analysis, 24 samples were identified to be positive for each of the three genes groEL, msp4, and 16S rRNA and each sequenced sample was associated with a host animal species and geographic location. Some samples were found to have each gene originated from the same animal. Consequently, 17 sequences of the groEL gene with 95.5–100% homology, 15 of the msp4 gene with 94.0–100% homology, and 3 of the 16S rRNA with 99.3–100% homology were identified. The sample sources, numbers, and accession numbers of sequences are detailed in Table 2.
In the phylogenetic tree, A. ovis sequences were separated into two distinct clusters based on the groEL gene. Six independent sequences derived from six sheep, three goats, and one yak were similar to those in isolates from other countries and were clustered together. Another 11 independent sequences obtained from 12 cattle and 2 yaks were similar to each other and were, therefore, grouped into a new cluster along with the previously identified isolates in Mongolian cattle (Fig. 2). The representative sequence alignments for the groEL gene of the Mongolian livestock pathogen were compared with sequences from animals in other countries, including South Africa, Cyprus, and China (Supplementary Fig. S1; Supplementary Data are available online at

Phylogenetic relationship between groEL gene sequences of Anaplasma spp. The phylogenetic tree was constructed by the neighbor-joining method using 580-bp-long sequences of the groEL gene of Anaplasma ovis; the data were supported by 1000 bootstrap replications.
In addition, the msp4 gene of A. ovis by using same and different samples with groEL gene demonstrated that 12 sequences derived from both bovine and ovine species (11 cattle, 1 yak, 6 sheep, and 3 goats) were clustered together and appeared genetically identical or similar to those in isolates from other countries. Three sequences derived from only bovine species (two yaks and one cattle) formed a separate branch (Fig. 3). The alignments of the representative msp4 gene sequences of the pathogen from Mongolian livestock were compared with alignments of sequences in isolates obtained from animals in other countries, such as Hungary, Italy, Spain, the United States, and China (Supplementary Fig. S2).
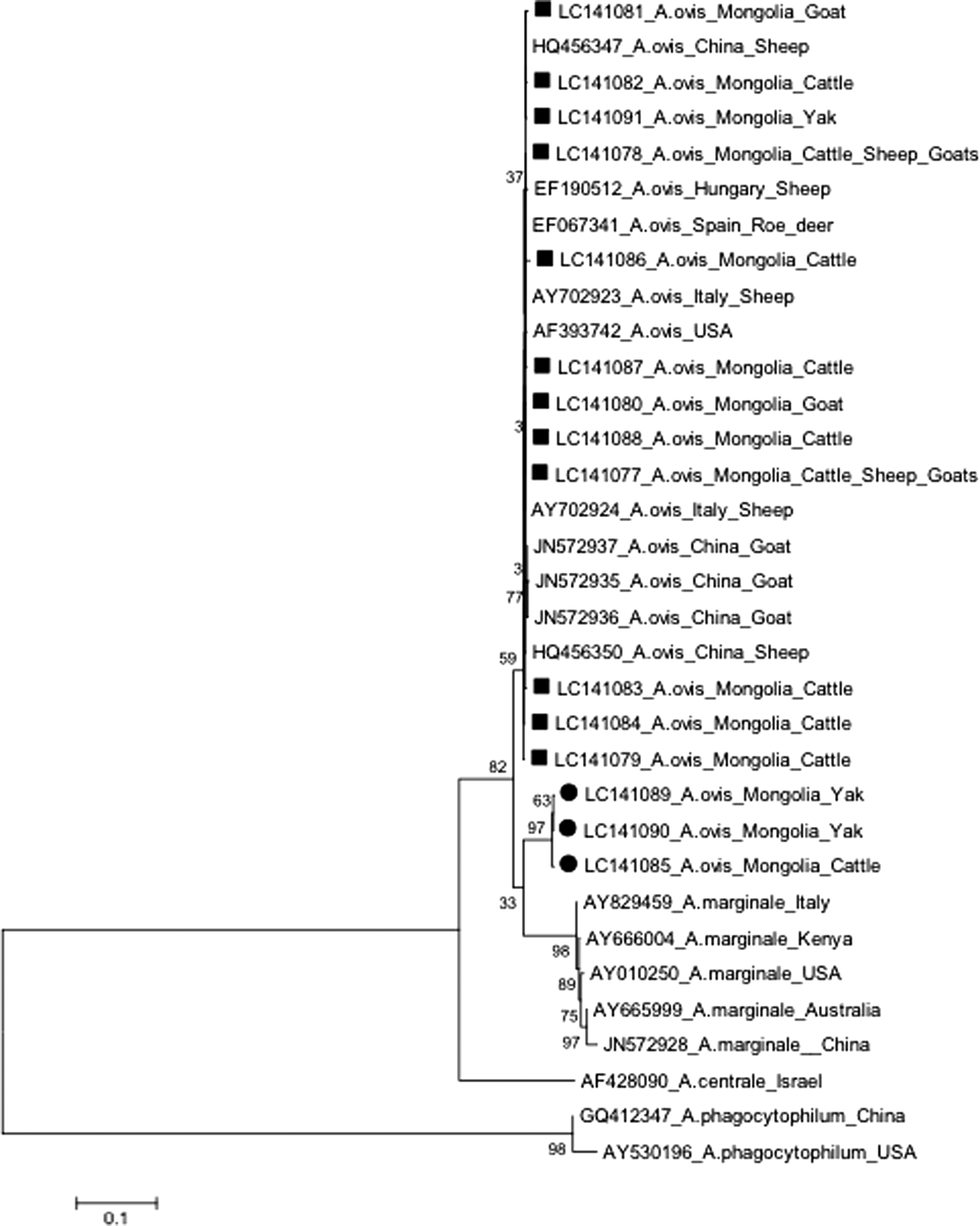
Phylogenetic relationship between msp4 gene sequences of Anaplasma spp. The phylogenetic tree was constructed by the neighbor-joining method using 870-bp-long sequences of the msp4 gene of A. ovis; the data were supported by 1000 bootstrap replications.
The relationships among these divergent clusters of A. ovis were elucidated by targeting the 16S rRNA gene, and three independent sequences were identified. Their nucleotide polymorphisms or divergences are detailed in Table 3. Two independent sequences (LC191433 and LC191434) derived from mainly ovine species (six sheep, three goats, and one yak) were similar to one another and were identical or similar to those in isolates from other countries, such as China. One independent sequence (LC191432) derived from bovine species (12 cattle and 2 yaks) appeared divergent from the other sequences in the phylogenetic tree (Fig. 4).

Phylogenetic relationship between 16S rRNA gene sequences of Anaplasma spp. The phylogenetic tree was constructed by the neighbor-joining method using 1430-bp-long sequences of the 16S rRNA gene of A. ovis; the data were supported by 1000 bootstrap replications.
The sequences that were divergent compared with the sequences from other countries.
The sequences that were similar to the sequences from other countries.
Discussion
The occurrence of infectious diseases in Mongolian livestock has been increasing every year and has been causing great economic loss to the livestock industry, a major income source (Beard et al. 2010, Batsukh et al. 2013, Wieland et al. 2015, Undrakhbayar et al. 2016). Reports on Anaplasma species, such as A. marginale, A. ovis, and A. phagocytophilum, in Mongolian livestock have been published (Sophia et al. 2012, Ybañez et al. 2013), but the prevalence and molecular characterization of these pathogens from various geographic regions remain unclear. Occasional occurrences of Anaplasma species in livestock lead to potential economic and public health threats (Haigh et al. 2008, Sophia et al. 2012). Anaplasma species may have been propagated through the animal population of the country and introduced in Mongolian livestock from neighboring countries because of geographical expansion or through Anaplasma pathogen vector tick distribution caused by global warming (Kocan et al. 2003).
Using nested or single-step PCR targeting both groEL and msp4 genes, A. ovis was detected in all animal species in all sampling regions. Higher infection rates based on the groEL gene were observed in native Mongolian animals, such as cattle (47.0%), sheep (95.2%), and goats (57.5%), than in yaks (27.8%) and dairy cattle (7.3%). These results may represent genetic variability of the pathogen or differing sensitivities of single-step or nested PCR or amplification conditions for PCR (e.g., unsuitable annealing temperature). A. ovis was more prevalent in Mongolian livestock than previously reported. In Khuvsgul and Khentii provinces, pathogen infection rates were 47.2% in goats, 39.8% in sheep, and 13.0% in cattle (Sophia et al. 2012). In Khuvsgul Province, the infection rate was 80.0% in reindeer (Haigh et al. 2008).
Previous studies on A. ovis have mostly used small ruminants, such as goat from Kenya (Ndung'u et al. 1995); sheep and deer from the United States (de la Fuente et al. 2007); sheep from Iraq, Turkey, Sudan, and Portugal (Renneker et al. 2013), and China (Yang et al. 2015); and sheep, goats, and cattle from Mongolia (Sophia et al. 2012). A. ovis appears host specific, primarily infecting small ruminants. In this study, A. ovis was associated with high rates of infection in Mongolian cattle and yaks (13.0%). This result demonstrated that A. ovis can infect cattle or yaks; moreover, our study may be the first to report on A. ovis infections in yaks. The overall prevalence of A. ovis was 95.2% for sheep. Goats from Mongolia displayed higher prevalence (57.5%) than those from Turkey (31.4%), Sudan (41.6%; Renneker et al. 2013), China (40.5%; Yang et al. 2015), or the United States (37%; de la Fuente et al. 2007). Prevalence was similar to or lower than that observed in Iraq (66.6%) and Portugal (82.5%) (Renneker et al. 2013). These results indicated that A. ovis is widely distributed among Mongolian livestock, especially native animals, including cattle, sheep, and goats. Dairy cattle and yaks were less susceptible to infection with A. ovis.
A. ovis causes mild clinical symptoms in infected animals, and stress factors, such as drought or heavy tick infestation, induce the virulence of this pathogen (Renneker et al. 2013). A previous report has described that reindeer, naturally infected with A. ovis, displays various clinical symptoms, such as fever, lethargy, and pale mucous membranes, occasionally causing sudden death in Khuvsgul Province of Mongolia (Haigh et al. 2008). In this study, A. ovis-infected animals (cattle, yak, sheep, and goats) appeared healthy and demonstrated no visible clinical symptom during the sampling period. Mongolian animals may have a tolerance or resistance to A. ovis because of the widespread prevalence of the pathogen throughout Mongolia.
Other Anaplasma species have been reported in Mongolian livestock. A. marginale is reportedly found in 8.7% of cattle (Ybañez et al. 2013) in Khentii, Uvurkhangai, and Uvs provinces. A. phagocytophilum is reportedly found in various species (35.8%) throughout Khentii and Khuvsgul provinces (Sophia et al. 2012). The vectors of A. phagocytophilum were Ixodes persulcatus and D. nuttalli from Selenge Province in Mongolia (Javkhlan et al. 2014). No report on A. bovis in Mongolian livestock has been published; however, pathogen transmission is possible, and it has been reported in the neighboring country China (Yang et al. 2015). In this study, we could not detect several Anaplasma species. A. marginale was screened by targeting three different genes using the same methods as those used in a previous study by our group in Philippines (Ochirkhuu et al. 2015). A disadvantage of PCR assays for A. phagocytophilum and A. bovis is the lack of a positive control; however, PCR was performed using several different conditions, such as different annealing temperatures. A. marginale, A. phagocytophilum, and A. bovis were not prevalent in livestock from our sampling regions.
The phylogenetic analysis of A. ovis from Mongolian livestock revealed two divergent varieties based on groEL gene sequences. The first branch containing sequences from sheep and goats was genetically similar to the sequences in common isolates from other countries, such as China, South Africa, and Cyprus. The second branch comprising sequences from cattle and yaks was quite distinct compared with the sequences from sheep and goats in this study as well as the available Genbank sequences. An exception to this was a previously published single sequence (Ybañez et al. 2013), identified as an Anaplasma species (JQ735903), from Mongolian cattle. This sequence was not declared as A. ovis because of limited study (Fig. 2).
Consequently, msp4 gene sequences of A. ovis using samples with both similar and different groEL gene sequences demonstrated that all sequences were derived from both ovine and bovine species. An exception was three sequences derived from two yaks and one cattle that were similar to each other and were clustered with several isolates from other countries, such as China, Italy, Spain, Hungary, and the United States (Fig. 3). Interestingly, a sequence from a yak demonstrated divergence based on both groEL and msp4 genes (LC141112 and LC141089). Two divergent sequences based on the msp4 gene (LC141085 and LC141090) from cattle and yak were not sequenced for the groEL gene because of mismatched PCR results. They may belong to a divergent cluster based on the groEL gene because all sequences from bovine species, except for one from yak (LC141103), were grouped into a divergent cluster in the phylogenetic tree (Fig. 2).
The 16S rRNA gene is a well-conserved bacterial region that is used to classify Anaplasma species (Dumler et al. 2001). We confirmed genetic differences in A. ovis isolated from Mongolian animals. The sequences from sheep (LC194133) and goats (LC194134) demonstrated 99.8% homology to each other but 99.3% divergence from sequences derived from cattle. Two sequences from yaks were identical to sequences from cattle and another sequence was identical to a sequence from sheep (Fig. 4). Based on groEL and 16S rRNA genes, two genetically distinct A. ovis sequences were identified in Mongolian livestock: one was identified in sheep and goats (with up to 95.5% homology) and the other was derived only from cattle (with up to 99.3% homology). Each genetically divergent A. ovis sequence was identified in yaks. These sequences were identical or similar to one another, based on the msp4 gene. In contrast, three sequences derived from yaks and cattle were up to 94.0% divergent from others.
Infectious diseases threaten animal husbandry and can cause enormous economic losses. Veterinary authorities should consider that the infectious Anaplasma species, including A. ovis, are widely prevalent and should include strategies for preventing and managing diseases caused by them in livestock. Additional studies on the molecular traits, pathogenesis, and tick vectors of all Anaplasma species in Mongolian livestock will generate additional information and a more comprehensive understanding of these pathogens.
Availability of Data and Material
The data sets supporting the conclusions of this article are included within the article and its additional file. Sequences are submitted in the GenBank database under accession nos. LC141077-LC141103 and LC194127-LC194134.
Footnotes
Acknowledgments
This work was partially supported by the Program for Leading Graduate Schools, Hokkaido University. This work was supported by the Research Program on Emerging and Re-emerging Infectious Diseases from Japan Agency for Medical Research and Development (AMED), grants-in-aid for Scientific Research from the Japan Society for the Promotion of Science (JSPS), Technology Research Promotion Program for Agriculture, Forestry, Fisheries and Food Industry, Japan (grant 26058B to S.K.), and the NARO, Bio-oriented Technology Research Advancement Institution (the special scheme project on regional developing strategy: grant 16817557 to S.K.).
Authors' Contributions
N.O. and S.K. designed this study and critically revised the article. S.M. and K.O. participated in its design, coordination, and article revision. N.O., S.K., and R.O. participated in sample collection. N.O. and S.K. performed the experiments, data analysis, and drafted the article. All authors read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.