
Abstract
Pathogens transmitted by ticks are the leading cause of arthropod-associated human diseases in the United States and managing the risk of exposure to potentially infected ticks is of vital public health importance. A 3-year integrated tick management program to control blacklegged ticks, Ixodes scapularis, the primary vector for the pathogenic agents of Lyme disease, human anaplasmosis, and babesiosis, was implemented in the town of Redding in southwestern Connecticut beginning in 2013. Combinations of white-tailed deer, Odocoileus virginianus, reduction, area application of the entomopathogenic fungus Metarhizium anisopliae, and fipronil-based rodent bait boxes were evaluated for their ability to reduce nymphal I. scapularis over 3 years. Interference from local hunters prevented sufficient, sustained deer removal previously reported to negatively impact I. scapularis abundances (i.e., <5 deer/km2). The combination of fipronil-based bait boxes and broadcast application of M. anisopliae had the most impact of any treatment combination; questing nymphs were reduced 78–95% within each year and Borrelia burgdorferi-infected questing nymphal I. scapularis encounter potential was reduced by 66% as compared with no treatment in the third year of the study. A combination of the broadcast application of M. anisopliae and small rodent-targeted fipronil-based bait boxes is an effective low-toxicity integrated approach that significantly reduced encounters with B. burgdorferi-infected questing nymphal I. scapularis on individual properties.
Introduction
L
Managing the risk of exposure to potentially infected I. scapularis has traditionally relied upon prevention through the use of personal protective measures and implementation of methods to control tick abundance. These measures include area-wide application of natural and synthetic compounds, vegetation management, and landscape modifications (Stafford and Kitron 2002, Williams et al. 2009, Williams and Ward 2010), host reduction or exclusion (Kilpatrick et al. 2014, Kugeler et al. 2016), host-targeted acaricides or vaccines (Telford et al. 2011, Gomes-Solecki 2014, Richer et al. 2014, Melo et al. 2016), and biological control (Ginsberg and Stafford 2005, Piesman and Eisen 2008, Eisen and Dolan 2016). Individual measures may be partially effective at reducing the abundance of ticks and prevalence of associated pathogens, however, recent emphasis has been given to the integration of a variety of methods within the framework of integrated tick management (ITM) (Ginsberg and Stafford 2005).
Currently, limited empirical data on the effectiveness of integrated tick control methods are available. In an earlier study, white-tailed deer, Odocoileus virginianus, exclusion, vegetation management, and an acaricide application in combination was found more effective in reducing the abundance of the lone star tick, Amblyomma americanum, compared with use of individual methods (Bloemer et al. 1990). In a more recent study, it was found that the combination of four-poster topical treatment devices (to target tick infestations on deer), fipronil-based bait boxes, and application of a pyrethroid acaricide reduced host-seeking nymphal I. scapularis abundance by 86.6% and 94.3% after 1 and 2 years, respectively (Schulze et al. 2007). Although these studies are foundational contributions to better understanding ITM, clearly the current dearth of field-based studies is a limiting factor for further development and application of tick control measures. Furthermore, although application of conventional acaricides has proven effective in some settings, there is increasing interest in integrating compounds with little to no environmental toxicity that have not been fully explored to date (Regnault-Roger et al. 2012).
Earlier studies have shown that significant deer removal can be effective in reducing I. scapularis abundance (Wilson et al. 1984, Stafford et al. 2003, Rand et al. 2004, Stafford 2007). More recently, one study showed that deer reduction significantly decreased the number of LD diagnoses in a residential community (Kilpatrick et al. 2014). However, these studies were conducted on islands or areas of an insular nature with limited immigration and emigration of white-tailed deer. It was our intent to determine if similar results could be achieved in a noninsular setting as white-tailed deer have been shown to be highly philopatric; deer typically will not vacate established home ranges to occupy areas of lower density (McNulty et al. 1997, DeNicola and Williams 2008, Simard et al. 2013).
The present study examined the effectiveness of an ITM approach based on minimally environmentally toxic components in reducing the abundance of nymphal I. scapularis in residential areas. The three components of the low-toxicity integrated strategy included (1) white-tailed deer population reduction (Stafford 1993, DeNicola et al. 2000, Kilpatrick et al. 2014), (2) spray application of the entomopathogenic fungus Metarhizium anisopliae (Bharadwaj and Stafford 2010, 2012, Stafford and Allan 2010), and (3) distribution of fipronil-based rodent bait boxes (Dolan et al. 2004). The objectives of this study were to (1) Develop an effective, reduced-risk ITM approach, within a residential community using an array of least toxic measures that are safe, inexpensive, and simple to implement. (2) Measure efficacy of combined methods to reduce questing nymphal densities as a means of reducing human exposure to B. burgdorferi. (3) Determine if deer reduction alone and/or in combination with other ITM interventions is an effective nymph reduction strategy in a noninsular setting. (4) Determine the within-year effectiveness of the M. anisopliae/fipronil-based rodent bait box treatment combination. We implemented the integrated control measures and observed their combined effectiveness within each year of implementation, during the period when peaks in nymphal densities and risk of human exposure to B. burgdorferi typically occur in this region (Stafford and Magnarelli 1993, Bacon et al. 2008) over the course of the study.
Materials and Methods
Study area and experimental design
This study was conducted from January 2013 through September 2016, in the town of Redding, Connecticut (41.3044°N, −73.3928°W). Redding encompasses 83 km2 in Fairfield County in southwestern CT, where LD has become endemic. From 2009 through 2016, 8–18 human cases of LD (87–197 cases/100,000 population) were reported annually in Redding (Connecticut Department of Health 2017).
Experimental plots were distributed across residential properties in four targeted 2.6 km2 study sites. The four sites were selected based on uniformity in size and landscape characteristics. Considerations were made with respect to intersite distances to minimize the possibility of deer movement between study sites (Kilpatrick and Spohr 2000, Rhodes et al. 2010). Experimental plots within target sites were selected based on the presence of woodland–lawn edge of at least 100 meters on the property, as well as accessibility and homeowner participation. Average property size was ∼2 ha.
Selected properties were utilized for inclusion in the study based on homeowners' permission to access their land. While a few properties had shared boundaries, the majority were not contiguous to one another. In 2013, access to five properties was secured within one study site to receive the deer removal-only treatment (deer removal). Elsewhere in town, six properties were secured to receive a combination of the broadcast application of the M. anisopliae acaricide (Met52; Novozymes Biologicals, Inc., Salem, VA) and distribution of fipronil-based rodent bait boxes (Select TCS™; Tick Box Technology Corporation, Norwalk, CT) (bait box/Met52). Additionally, four properties were secured to receive all three interventions: Met52, fipronil-based rodent bait boxes, and deer removal (deer removal/bait box/Met52). Finally, six properties were used as experimental controls and received no intervention (control).
In 2014, additional properties were added to each of the four study sites: three were added to the deer removal treatment, seven to bait box/Met52, one to deer removal/bait box/Met52, and six to the control, increasing total properties from 21 in 2013 to 38 in 2014. The 21 properties were sampled and received interventions in 2013, and all 38 properties were sampled and received assigned interventions in 2014 and again in 2015. Met52 was not available in 2016 and as a result, was not applied to assigned properties. That, and the fact that deer removals ceased in early 2015 due to safety concerns resulted in minimal intervention in 2016. As a result, the majority of data presented are from 2013 to 2015. See Table 1 for treatment assignments and number of participating properties.
Number of participating properties are in parentheses.
Met52/ M. anisopliae
The active ingredient in Met52, M. anisopliae, is a naturally occurring soil-borne fungus that causes green muscardine disease in insects and has been shown to be pathogenic to I. scapularis nymphs and adults in the laboratory (Zhioua et al. 1997, Benjamin et al. 2002, Kirkland et al. 2004, Bharadwaj and Stafford 2012) and field settings (Bharadwaj and Stafford 2010). The deer removal/bait box/Met52 and bait box/Met52 properties received broadcast applications of Met52, which contained 11% w/w of M. anisopliae or 5.5 × 109 CFU/g of product, which was applied at a rate of 0.63–0.96 mL/m2, twice per season in early June and July 2013, 2014, and 2015 or as dry weather conditions dictated. Each property was sprayed by a licensed applicator (Connecticut Tick Control, Norwalk, CT) within an area encompassing 3 meters on either side of a woodland–lawn transect where drag sampling also occurred (described below).
Fipronil-based bait boxes
The fipronil-based rodent bait boxes (Select TCS; Tick Box Technology Corporation) targeted subadult I. scapularis parasitizing small rodents. The boxes permitted free passage of small rodents attracted to two nontoxic bait blocks on either side of the rear of the box. A wick attached to the top of the box was saturated with a 0.7% solution of a topical formulation of fipronil, which small rodents entering the box were forced to make contact with. A galvanized metal shroud prevented larger mammals from gnawing through the plastic to access the bait. The box/shroud combination was staked to the ground to prevent movement or theft of boxes by medium-sized mammals. The use of similar passive acaricide delivery devices was shown to reduce nymphal and larval I. scapularis parasitizing white-footed mice, Peromyscus leucopus, the principal reservoir for several pathogens, including B. burgdorferi, by 68% and 84%, respectively, in field applications (Dolan et al. 2004). More recently, similar fipronil-based bait boxes with doxycycline hyclate-laden bait were shown to reduce nymphal I. scapularis burdens on small rodents by 76% while reducing B. burgdorferi infection in reservoir hosts by 96% (Dolan et al. 2016).
Bait boxes were distributed on plots approximately every 10 meters along property perimeters. In 2013 at the bait box/Met52 treatment properties, 65 bait boxes were placed at the 6 properties and 47 were distributed at the 4 deer removal/bait box/Met52 properties in May. All bait boxes were replaced with new boxes midseason at each location. Boxes were removed completely and inspected for usage and bait consumption in early fall of 2013. In 2014, 2015, and 2016, 152 bait boxes were placed at the 13 bait box/Met52 properties and 61 at the 5 deer removal/bait box/Met52 properties. Those boxes also were replaced midseason and were retrieved in early fall. All rodent bait boxes showed signs of heavy usage for all years of the study.
Deer removals
Deer were removed by professional sharpshooters (White Buffalo, Inc., Moodus, CT) throughout the deer removal and the deer removal/bait box/Met52 treatments as regulated hunting as a management tool in noninsular suburban settings has yet to achieve area-wide densities much below 17 deer/km2 (Williams et al. 2013). Deer were concentrated with bait on cooperating homeowners' properties and euthanized with a single 0.223 caliber bullet to the center of the brain, consistent with the American Veterinary Medical Association and Sikes and Gannon (2011) standards for humane euthanasia and in compliance with The Connecticut Agricultural Experiment Station's Institutional Animal Care and Use Committee (#P18-13) and Connecticut Department of Energy and Environmental Protection (Volunteer Authorization #1315006b). All venison was donated through local charities (WhiteTail Solutions, LLC, Town of Redding Food Pantry, Bridgeport Rescue Mission). Goal deer density sought for I. scapularis reduction was below 4.0 deer/km2 on each of the two research areas.
Questing I. scapularis nymph sampling
Host-seeking, or questing I. scapularis nymphs were sampled from all participating properties from May to July in 2013, 2014, and 2015 by dragging vegetation with a tick drag consisting of 1.0 m2 of white cloth attached to a wooden dowel. Follow-up drag sampling was done on the control and bait box/Met52 properties in 2016. During each sampling session, we dragged along permanent transects of known distance running the length of woodland–lawn edge habitats. Drag sampling occurred specifically along the lawn-side of the woodland–lawn edge, the area that corresponds to peak density of questing nymphal I. scapularis in peridomestic habitats (Stafford and Magnarelli 1993). All plots were sampled approximately every 2 weeks, and at the end of the study, density of sampled questing nymphs (nymphs/100 m2) was determined for each property within each treatment area. Nymphs were removed from drags, transferred to labeled vials, and ultimately held in humidity chambers at 4°C. We later confirmed species identities and life stages of all collected tick specimens and stored them at −80°C for B. burgdorferi screening.
B. burgdorferi screening in questing I. scapularis nymphs
Genomic DNA was extracted from field-collected ticks by using the DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA) or DNAzol (Molecular Research Center, Cincinnati, OH) according to the manufacturer's recommendations, with some modifications (Molaei et al. 2006a, b). Briefly, ticks were first washed three times in 500 μL sterilized, deionized water, and homogenized in microtubes containing 400 μL DNAzol manually or by using a copper BB and a vibration mill. The homogenates were centrifuged at 14,000 g for 10 min. After adding 3 μL of Polyacryl Carrier (Molecular Research Center) to the supernatant, DNA was then precipitated by using absolute ethanol. The DNA pellet was washed twice with 75% ethanol, air-dried briefly, reconstituted in 30 μL of 1 × TE buffer (10 mM Tris-HCl [pH 8.0], 1 mM EDTA), and stored at −20°C for further analysis. Isolated DNA from the ticks served as templates in subsequent PCR reactions to screen for infection with B. burgdorferi using specific primer sets for flagellin (Barbour et al. 1996), the 16S rRNA (Gazumyan et al. 1994), and Osp A (Persing et al. 1990) genes. DNA isolated from B. burgdorferi cultures and from uninfected laboratory-reared ticks was used as positive and negative controls, respectively, in all PCR reactions. The Taq PCR Core Kit (Qiagen, Germantown, MD) was used for all PCR reactions according to the manufacturer's recommendation. A 50-μL reaction volume was prepared with 3 μL template DNA, 4 μL each primer (0.1–0.5 μM), 5 μL 10 × Qiagen PCR Buffer (containing 15 mM MgCl2), 1 μL dNTP mix (10 mM each), 0.25 μL Taq DNA polymerase (1.25 U/reaction), and 32.75 μL water. PCR cycling condition for flab included an initial reaction activation step at 95°C for 3 min, followed by 40 cycles of denaturation at 95°C for 1 min, annealing at 55°C for 1 min, and extension at 72°C for 1 min. The final cycle was completed with 5 min of extension at 72°C. Cycling condition for 16S rRNA included an initial reaction activation step at 95°C for 10 min, followed by 35 cycles of denaturation at 95°C for 1 min, annealing at 54°C for 1 min, and extension at 72°C for 1 min and 20 s. The final cycle was completed with 7 min of extension at 72°C. Cycling condition for Osp A included an initial reaction activation step at 94°C for 10 min, followed by 45 cycles of denaturation at 95°C for 1 min, annealing at 50°C for 45 s, and extension at 72°C for 2 min. The final cycle was completed with 7 min of extension at 72°C. All PCR reactions were performed with Veriti or the GeneAmp PCR System 9700 (Applied Biosystems, Foster City, CA). PCR-amplified products were run on 0.6–1.2% agarose gel, stained with ethidium bromide and visualized under UV light and documented using GelDoc system (UVP, Upland, CA).
Aerial deer counts over snow
We attempted a total count of all animals within each of the four research areas using the double observer method, similar to Beringer et al. (1998) as weather conditions permitted on February 15, 2013, January 24, 2014, and March 3, 2015. We used a Robinson 44 helicopter (Robinson Helicopter Company, Torrance, CA) with a pilot, two experienced observers on either side of the rear of the aircraft, and a data recorder in the copilot seat. We flew 200-m-wide transects that were pre-established in the geographic information system program ArcView (version 3.3; ESRI, Inc., Redlands, CA) and were overlain on a GPS-ready, heads-up display moving map software program (Geolink® Version 6.1; Michael Baker Jr., Inc., Shreveport, LA) on a digital aerial sketch mapper (DASM; Dell Latitude XT2 XFR Hammerhead). The pilot attempted to maintain an altitude of 60 meters and air speed of 40 km/h, although altitude and air speed varied somewhat throughout the flights. Observers maintained a 100 meters search area either side of the aircraft and communicated deer sightings to the data recorder who entered their location and abundance into the DASM in real time. The pilot navigated from transects displayed on the DASM to insure full coverage of each area. Snow varied in depth from 15 cm in 2014 to 76 cm in 2015. Based on previous research using this technique and Beringer et al. (1998), we assumed that two experienced observers had 80% detection, but had to adjust values in target areas with heavy coniferous or mountain laurel, Kalmia latifolia, cover that was difficult to visually penetrate from the air.
Statistical analyses
Nymphs were not encountered on 62% of sampling occasions and as a result, data were severely non-normally distributed and were unable to be normalized through standard data transformations. As a result, we used Pearson's chi-squared test for categorical data where sampling sizes were large. We used Pearson's chi-squared test with Yate's correction for continuity in the B. burgdorferi-infected questing nymph analysis, where there were small sample sizes (i.e., single-digit encounter data) for within-year comparisons. For within years and for the combined 3 years, we used Pearson to determine multiple pairwise differences in treatment combinations for encountering at least one questing nymph per sampling occasion, for encountering at least one B. burgdorferi-infected questing nymph per sampling occasion, and for B. burgdorferi infection prevalence in questing I. scapularis. I. scapularis encounter data were deemed significantly different at p < 0.05.
We also calculated percent control (i.e., percent reduction in questing nymphal density) associated with the within-year bait box/Met52 treatment. We used mean density (nymphs/100 m2) for 2–3 sampling occasions in early spring (late May to early June) before the Met52 treatment was administered (baseline) to compare 5–6 nymphal sampling occasions post-Met52 treatment through the end of July (summer) for each of the 4 years at all locations. The formula used was: % control = 100[1−(XcYt)/(XtYc)], where Xc and Xt are baseline densities on control properties and bait box/Met52 properties, respectively, and Yc and Yt are the summer densities from the same respective plots (Henderson and Tilton 1955). We calculated percent control on questing nymphs as well as B. burgdorferi-infected questing nymphs using infection rates derived from ticks tested each year for each property. We also included data from 2016 to compare effectiveness of fipronil-based bait boxes only without the integrated approach of including the broadcast application of Met52. We used comparable baseline and summer sampling dates for 2016 to previous years as there was no application of Met52.
Results
Deer removals
Over 3 weeks in February to March 2013, 51 deer were removed from the two study areas resulting in a 12.0 deer/km2 reduction of the deer removal only treatment (n = 31) and 7.7 deer/km2 reduction at the deer removal/bait box/Met52 treatment (n = 20). Results from the 2013 aerial survey indicated that we were approaching goal densities. While there were fewer deer in the deer removal/bait box/Met52 treatment area, dominated by a 113-ha piece of municipally owned forested land, densities were likely double what we sought. After initial reduction, local hunters intervened by promoting a smear campaign of misinformation on the project. As a result, the deer removal portion of the study was severely compromised, had to be prematurely suspended in 2015 due to safety concerns, and goal densities were not achieved. Despite this, 25 more deer were removed in 2014 resulting in a 4.2 deer/km2 reduction from the deer removal only treatment (n = 11) and 5.4 deer/km2 reduction from the deer removal/bait box/Met52 treatment (n = 14). Eleven deer were removed in 2015 resulting in a 2.3 deer/km2 reduction from the deer removal only treatment (n = 6) and 1.9 deer/km2 reduction from the deer removal/bait box/Met52 treatment (n = 5). In our 2014 aerial survey, we reported initial conservative densities of 14.7 deer/km2 in the deer removal only treatment and 7.3 deer/km2 in the deer removal/bait box/Met52 treatment. Despite seemingly approaching goal densities, at least in the deer removal/bait box/Met52 treatment, our final postremoval aerial snow count in March 2015 resulted in 19.7 deer/km2 in the deer removal only treatment area and 6.2 deer/km2 in the deer removal/bait box/Met52 treatment area, both above target densities of a sustained annual 4.0 deer/km2. Despite not reaching goal densities, a total of 87 deer were removed from the two 2.6 km2 research areas over 3 years, largely after the 4.5 month regulated hunting season had expired.
I. scapularis questing nymph encounters
In 2013, each of the 21 cooperating properties were drag sampled for nymphs on 10 different occasions, during summer months corresponding to peak nymphal activity (May to July). This sampling effort resulted in 171 nymphs. In 2014, the 38 properties were each sampled on nine different occasions resulting in 109 nymphs collected. In 2015, the same 38 properties were sampled on eight occasions resulting in the collection of 138 nymphs (Table 2).
In 2013, questing nymphal I. scapularis encounters were significantly lower for the bait box/Met52 treatment as compared with control (χ2 = 6.3, p < 0.01), deer removal only (χ2 = 8.4, p < 0.01), and the deer removal/bait box/Met52 treatment (χ2 = 7.3, p < 0.01) (Fig. 1). In 2014, no significant differences existed between any treatments (Fig. 1). In 2015, nymphal I. scapularis encounters were significantly lower for the bait box/Met52 treatment as compared with control (χ2 = 12.3, p < 0.001) and deer removal only treatment (χ2 = 4.1, p < 0.03). Differences also existed between control and the deer removal/bait box/Met52 (χ2 = 3.0, p < 0.05) (Fig. 1). Combined results for all 3 years were also similar to 2013: nymphal I. scapularis encounters were significantly lower for the bait box/Met52 treatment as compared with control (χ2 = 12.0, p < 0.001), deer removal only (χ2 = 13.3, p < 0.001), and the deer removal/bait box/Met52 treatment (χ2 = 7.8, p < 0.01) (Fig. 1).

Positive encounters for ≥1 questing Ixodes scapularis nymph for May to July 2013, 2014, 2015 and the average for all 3 years combined for all treatment assignments. Columns with the same letter are not significantly different within each year.
B. burgdorferi-infected questing nymph encounters
To increase sample size for evaluating infection prevalence only, 109 additional nymphs were sampled on participating properties in the different treatments areas, but were off established transects. The prevalence of infection with B. burgdorferi for all 418 on-transect and 109 off-transect questing nymphs averaged 29% for all 3 years. Differences in infection prevalence between treatments were not significant in 2014 or 2015 (Table 3).
Percent infection with the same letter is not significantly different within each year.
In 2013, deer removal resulted in the perceived increased potential to encounter B. burgdorferi-infected questing nymphs. The deer removal and deer removal/bait box/Met52 treatments showed the highest encounter potential and were not significantly different (χ2 = 0.1, p = 0.6) and both were significantly higher than control (χ2 = 5.0, p < 0.02, χ2 = 3.8, p < 0.03, respectively) (Fig. 2). The bait box/Met52 treatment had no infected nymphs, which was significantly lower than for the deer removal only treatment (χ2 = 17.7, p < 0.001), control (χ2 = 6.2, p < 0.01), and the deer removal/bait box/Met52 treatment (χ2 = 15.9, p < 0.001) (Fig. 2). In 2014, there were no statistical differences between any treatment combinations (Fig. 2). In 2015, the bait box/Met52 treatment B. burgdorferi-infected questing nymph encounter potential was significantly lower than control (χ2 = 5.6, p < 0.01) and the deer removal/bait box/Met52 treatment (χ2 = 5.2, p < 0.02) (Fig. 2). Encounter potential for all 3 years combined resulted in the bait box/Met52 treatment significantly lower than the deer removal only treatment (χ2 = 19.1, p < 0.001), control (χ2 = 6.4, p < 0.01), and the deer removal/bait box/Met52 treatment (χ2 = 16.4, p < 0.001) (Fig. 2). Additionally, the deer removal only treatment encounter potential with a B. burgdorferi-infected questing nymph was significantly greater than control (χ2 = 3.7, p < 0.04) (Fig. 2).
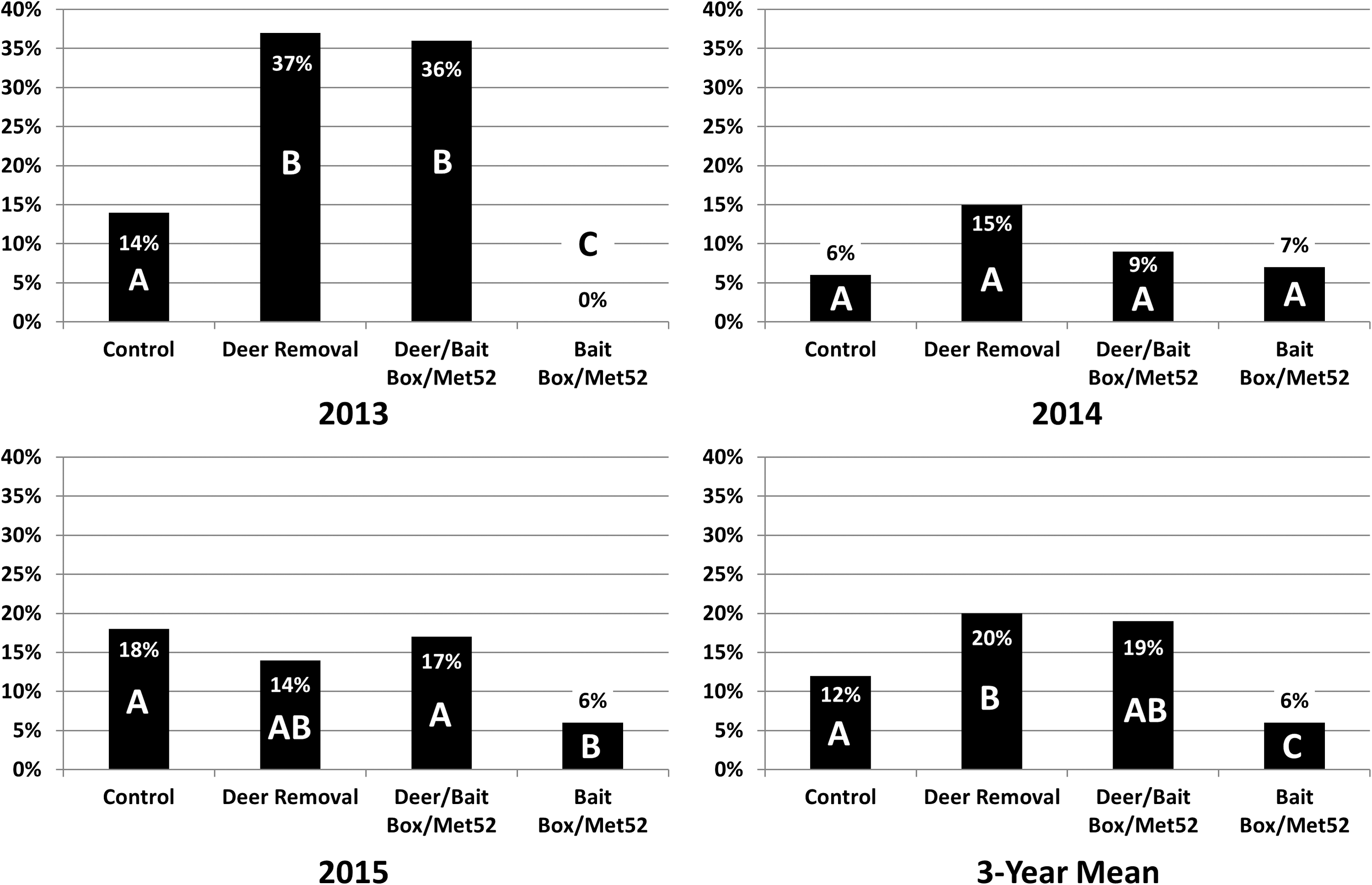
Positive encounters for ≥1 Borrelia burgdorferi-infected questing I. scapularis nymph for May to July 2013, 2014, 2015 and the average for all 3 years combined for all treatment assignments. Columns with the same letter are not significantly different within each year.
Within-year M. anisopliae effectiveness
The combination of the application of M. anisopliae and rodent bait box distribution was effective in significantly reducing the total number of questing I. scapularis nymphs as well as B. burgdorferi-infected questing I. scapularis nymphs within each of the 3 years it was applied (Tables 4 and 5). However, in 2016, the year M. anisopliae was not applied, the distribution of rodent fipronil bait boxes alone was ineffective at maintaining reduced density of both questing I. scapularis nymphs and B. burgdorferi-infected questing I. scapularis nymphs (Tables 4 and 5).
Discussion
Results of this study indicate that an integrated strategy involving a combination of a spray application of the entomopathogenic fungus M. anisopliae and distribution of fipronil-based rodent bait boxes provided the most effective control of I. scapularis, the principal vector of B. burgdorferi in North America (Piesman et al. 1987). Following 3 years of intervention, this treatment combination proved to significantly reduce both encounter potential with a questing nymph (Fig. 1) and encounter potential with a B. burgdorferi-infected nymph (Fig. 2) as compared with no treatment. Previous studies also have reported on the effectiveness of integrated control methods in reducing tick abundance (Bloemer et al. 1990, Schulze et al. 2007); however, area-wide application of synthetic acaricides was included in these studies. Although many acaricides are considered to be relatively safe for use in peridomestic habitats, their unintended consequences for the environment and public health cannot be entirely disregarded (Wargo 1996, Stafford 2007). By contrast, the integrated control strategy implemented in this study relies on an entomopathogenic fungus that occurs naturally and has no known mammalian toxicity (Zimmerman 2007) and a low-dose host-targeted acaricide that is frequently used as an antiparasitic treatment for domestic dogs and cats (Jacobson et al. 2004). It is thus a low-toxicity alternative to the suite of integrated strategies shown to be effective in controlling I. scapularis.
In summer 2013, several months after significant deer reduction, questing B. burgdorferi-infected nymphal I. scapularis encounters at both the deer removal only treatment and the deer removal/bait box/Met52 treatment were more than 2.5 times higher than control (Fig. 2). This temporary amplification was likely the result of the acute removal of numerous large-bodied, although, pathogenic dead-end hosts. Perkins et al. (2006) documented similar amplification of questing I. scapularis in small areas (<2.5 ha), where deer were excluded, although documented a reduction in questing I. scapularis in deer exclusion areas >2.5 ha. Similar amplification of questing adult I. scapularis was documented on Monhegan Island, ME the same year deer were eliminated, but I. scapularis abundance declined precipitously to insignificant densities over the next 3 years in the absence of deer (Rand et al. 2004). Adult female I. scapularis loads on deer were documented to increase proportionally with the number of deer harvested in coastal Massachusetts; as more deer were harvested, I. scapularis load/deer increased (Deblinger et al. 1993), but ultimately, after several years, I. scapularis abundances declined significantly, following similar results as Wilson et al. (1984).
I. scapularis nymphs cause the majority of human cases of LD, despite generally lower infection prevalence than adults (Piesman and Spielman 1979, Piesman et al. 1986, Mather et al. 1990, Kugeler et al. 2016). This phenomenon is due, in part, to their small size and the fact that their peak activity corresponds to the summer months when humans are most active outdoors (Stafford 2007) and human LD cases are highly correlated with the abundance of B. burgdorferi-infected I. scapularis nymphs (Stafford et al. 1998). Approximately 95% of adult female I. scapularis require deer for their final bloodmeal (Wilson et al. 1990), but nymphs and larvae also commonly feed on deer (Watson and Anderson 1976, Anderson and Magnarelli 1980, Magnarelli et al. 1995, Garnett et al. 2011). It appears that deer as hosts for subadult I. scapularis has been generally underreported and underappreciated in large part due to the fact that deer are rarely sampled when subadult I. scapularis stages are active; most handling of deer occurs during hunting season (October to December) when only adult I. scapularis are active (Stafford 2007). Deer are considered a keystone host for adult female I. scapularis and are reservoir incompetent for the majority of tick-borne pathogens, particularly B. burgdorferi. Nevertheless, it is clear that deer are also important hosts for subadult stages of I. scapularis. While this temporary amplification phenomenon in the absence of deer has been previously reported in adults (Rand et al. 2004, Perkins et al. 2006, Elias et al. 2011), we believe this is one of the first reports to document it can also occur in nymphal I. scapularis populations as the result of initial deer reduction.
In 2013, both the deer removal/bait box/Met52 treatment and deer removal only treatment, questing I. scapularis nymphs were significantly higher, in fact double, for B. burgdorferi infection than both control and the bait box/Met52 treatment (Table 3) suggesting that in the absence of the 51 reservoir incompetent deer that were removed, remaining I. scapularis prioritized bloodmeals from competent reservoirs, specifically white-footed mice. Similar increases in B. burgdorferi infection in adult I. scapularis (from 45% to 75%) were documented on Monhegan Island, ME and declined as deer were eliminated (Rand et al. 2004). After the initial reduction in 2013, B. burgdorferi infection prevalence in questing nymphs for all treatment areas did not differ significantly in 2014 and 2015 (Table 3).
Despite an increase in cooperating homeowners from 21 to 38 in 2014, and an increased sampling effort, questing nymphal I. scapularis numbers declined considerably as compared with 2013 (Table 2), particularly at the control where there was no intervention. We can only assume this drop was due to environmental factors and/or I. scapularis intercycle variation. As a result, there were no detectable significant differences between any treatment combination for questing nymph or B. burgdorferi-infected questing nymph encounters in 2014.
In 2015, questing I. scapularis sampling resulted in more nymphs than in 2014, particularly at the control, the only area where encounter potential increased. Only 11 deer were removed from the deer removal only and the deer removal/bait box/Met52 treatments combined in 2015, although questing I. scapularis encounters continued downward trends from 2013 (Fig. 1). However, questing nymph and B. burgdorferi-infected questing nymph encounters remained lowest at the bait box/Met52 treatment. We attribute these results to the sustained targeting of subadult I. scapularis feeding on mice by use of bait boxes and broadcast treatment of Met52. Vertebrate host abundance was not manipulated at the bait box/Met52 treatment and resulting temporary amplification from host switching by subadult I. scapularis was not witnessed. Additionally, this treatment combination provided excellent control of questing nymphs and questing B. burgdorferi-infected nymphs within each year from 2013 to 2015 at the bait box/Met52 treatment (Tables 4 and 5), but in the absence of the Met52 application in 2016, nymphal control was not maintained. We can only speculate why this occurred, but feel it may be due, in part, to the treatment of single residential properties scattered throughout the town. Because small rodents are oblivious to human-derived boundary lines, fipronil-based bait boxes are likely more effective when deployed in a clustered or blanket approach, where multiple properties with shared borders are treated (Dolan et al. 2004).
It was initially assumed that with increased interventions, such as use of a combination of deer removal, bait boxes, and Met52, we would witness increased reductions in B. burgdorferi infection and abundance of questing nymphs, but this did not occur. This assumed achieving deer densities low enough to negatively impact I. scapularis abundance over the course of the study (Wilson et al. 1984, Stafford et al. 2003, Rand et al. 2004, Stafford 2007). There are numerous studies that document a positive correlation between I. scapularis and deer abundances, however, the majority have occurred in areas of an insular nature with limited ingress and egress of deer (Kugeler et al. 2016). While it was our intention to further investigate and document this positive relationship in a noninsular setting, ultimately, societal pressures through direct interference from the local hunting community prevented us from achieving a sustained low density of deer needed to negatively impact I. scapularis populations.
Conclusion
We were unable to determine if sufficient deer removal in a noninsular setting is a viable I. scapularis management strategy. Encounter potential with a B. burgdorferi-infected nymph remained similar for deer removal areas and the control in year 3 of the study. Marginal deer removal efforts after year 1 of this study likely resulted in subadult I. scapularis host switching and the subsequent temporary increase in nymphal B. burgdorferi infection as previously documented (Deblinger et al. 1993, Rand et al. 2004, Perkins et al. 2006, Kugeler et al. 2016). Without the continued decline in deer abundance, marginal deer removal in combination with broadcast application of the entomopathogenic fungus M. anisopliae and distribution of fipronil-based rodent bait-boxes proved to be significantly less effective than the use of M. anisopliae and bait boxes alone. Unfortunately, the efficacy of deer reduction as a tick management strategy in noninsular settings still remains unclear (Kugeler et al. 2016). Unless there is sufficient public and political support to commit to reducing deer densities to levels in the vicinity of 5/km2, individual homeowners looking for less toxic options could use a strict regimen of a combination of broadcast treatment of M. anisopliae and fipronil-based rodent bait boxes to significantly reduce encounters with nymphal I. scapularis and associated B. burgdorferi on their properties.
Footnotes
Acknowledgments
The authors are grateful to Redding town staff and elected officials, specifically Natalie Ketchum, Julia Pemberton, and Doug Hartline for their support and assistance as well as all the collaborating homeowners in town for allowing us access to their properties. They thank Laura Hayes for her earlier contribution to this project. They also recognize Michael Short, Heidi Stuber, Elizabeth Alves, and Saryn Kunajukr of The Connecticut Agricultural Experiment Station and seasonal research assistants Mark Morris, Benjamin DeMasi-Sumner, Stephanie Shea, Heather Whiles, Adam Misiorski, Kelsey Schwenk, Jamie DeNicola, Jarrod Bridge, Pronoma Srivastava, Magalí Bazzano, and Alex Diaz and for their technical assistance. Met52 was kindly made available by Novozymes Biological, Inc. This research was funded by a Cooperative Agreement between the Centers for Disease Control and Prevention and The Connecticut Agricultural Experiment Station (award # 5U01 CK000182-01).
Author Disclosure Statement
No competing financial interests exist.