
Abstract
From the group of parasitic protozoa, Giardia and Cryptosporidium are the most common pathogens spread in surface water sources, representing a continuous threat to public health and water authorities. The aim of this survey was to assess the occurrence and human infective potential of these pathogens in treated wastewaters and different surface water sources. A total of 76 western Romanian water bodies in four counties (Arad, Bihor, Caraș-Severin and Timiș) were investigated, including the effluents of wastewater treatment plants (n = 11) and brooks (n = 19), irrigation channels (n = 8), lakes (n = 16), and ponds (n = 22). Water samples were collected through polyester microfiber filtration. Giardia cysts and Cryptosporidium oocysts were isolated using immunomagnetic separation, according to the US EPA 1623 method, followed by their identification and counting by immunofluorescence (IF) microscopy. All samples were screened through PCR-based techniques targeting the gdh gene for Giardia spp. and the 18S rRNA gene for Cryptosporidium spp., followed by sequencing of the positive results. Cryptosporidium-positive samples were subtyped based on sequence analysis of the GP60 gene. Giardia spp. was found in all tested water types with a cumulative detection rate of 90.1% in wastewaters, 26.3% in brooks, 37.5% in irrigation channels, 31.2% in lakes, and 36.4% in ponds. Except for ponds, all monitored water bodies harbored the Giardia duodenalis AII subassemblage with human infective potential. In addition, the ruminant origin assemblage E was widely distributed, and the domestic/wild canid-specific assemblage D was also recorded in a pond. Three (27.3%) wastewater samples were Cryptosporidium positive, and the identified species was the zoonotic Cryptosporidium parvum, with IIaA15G2R1 (n = 2) and IIdA18G1 subtypes. The results highlight that this threat to the public health must be brought to the attention of epidemiologists, health officials, and water authorities.
Introduction
T
Molecular epidemiologic studies conducted over the last two decades show a remarkable genetic diversity within G. duodenalis and Cryptosporidium spp. Thus, G. duodenalis, the only species of six within the genus Giardia that has been detected in humans, encompasses eight genetically distinct assemblages (from A to H), with some that have further genetic substructuring, resulting in a variable number of subassemblages (e.g., AI–AIII, BIII, and BIV). Of the assemblages, only two (A and B) are responsible until now for intestinal infections in humans, with the remaining six (from C to H) assemblages with strong host specificity (Feng and Xiao 2011). Currently, of the over 30 valid Cryptosporidium species, C. parvum and C. hominis, which include 14 (IIa-IIo) and nine (Ia-Ij) subtype families, respectively, are most frequently associated with human cryptosporidiosis. In addition, the zoonotic potential of other species such as C. meleagridis, C. canis, C. felis, C. cuniculus, C. ubiquitum, and C. viatorum is occasionally reported (Ryan et al. 2016).
Because of the continuous reduction of drinking water supplies, as a consequence of global warming, emerging pathogens in natural surface water bodies are an increasing problem worldwide. Based on several epidemiology studies, different aquatic systems can harbor G. duodenalis cysts and Cryptosporidium oocysts as environmentally resistant and transmissible stages of these parasites. Thus, important (oo)cyst contamination levels have been detected in the effluents of biologically treated wastewater treatment plants and in recycled sewage (Castro-Hermida et al. 2008, Plutzer et al. 2008, Spanakos et al. 2015), which raise concern about the potential contamination of water sources used for agricultural, domestic, and industrial purposes. Giardia and Cryptosporidium (oo)cysts with human infective potential are also found in other surface water types used for recreation and irrigation or as a supply of domestic and drinking water (e.g., flowing water courses, lakes, ponds, and irrigation channels) (Coupe et al. 2006, Diallo et al. 2008, Plutzer et al. 2008, Lobo et al. 2009, Yvonne et al. 2009, Daniels et al. 2016).
In Romania from 2010 to 2015, the National Institute for Public Health reported a high cumulative incidence (89.07 cases per 100,000 inhabitants) for G. duodenalis in human patients (RNIPH 2016). Moreover, few studies describe the molecular evidence of Cryptosporidium spp. in livestock (Imre et al. 2011, 2013, Vieira et al. 2015) and humans (Vieira et al. 2015) and G. duodenalis in pets and wildlife (Sommer et al., 2015, Györke et al. 2016), but data on the occurrence of these pathogens in aquatic environmental samples are scarce. The zoonotic Giardia and Cryptosporidium were detected in river water samples in the only available study (Imre et al. 2016), suggesting the recurrent exposure of human population to waterborne outbreaks in our country, Romania.
With these considerations and to obtain a better understanding of the complex puzzle of the molecular epidemiology of cryptosporidiosis and giardiasis in western Romania, the current study investigated the presence and human infective potential of G. duodenalis and Cryptosporidium spp. from treated wastewaters and other surface water sources, including brooks, irrigation channels, lakes, and ponds.
Materials and Methods
Study area description
The study was conducted in four western Romanian counties (Arad, Bihor, Timiș, and Caraș-Severin) (Fig. 1). The area has a temperate climate, occupies 32,509 km2, and has an approximate population of 2,073,000, of which ∼1,168,000 are classified as urban. Eleven wastewater treatment plants that collect mixed sewage with rainwater, primarily from the largest urban municipalities of the screened area, agreed to participate in the study. The selected plants, with different treatment capacities (min–max 700–120,000 m3/day), apply preliminary, primary, and secondary operational treatment procedures for the received wastewater and discharge their final effluents into the nearest rivers. The investigated brooks are tributaries of the western Romanian rivers, crossing primarily rural areas. These flowing water courses are typically used for irrigation of vegetable crops and for livestock. The screened region comprises an important network of irrigation canals, derived from the main western rivers, which collect water excess during rainfall events and frequently receive illegal livestock or human wastewater. The water from these channels is used for recreational purposes, particularly during summer, and for irrigation of agricultural lands. The targeted natural and artificial lakes, located mostly in mountain regions, were selected based on their use for recreational activities, particularly during summer, with facilities for bathing, swimming, or boating. In addition, most of these lakes are an important domestic and drinking water supply for the surrounding human settlements. The selected counties contain a series of natural and human-constructed ponds with shallow water, designed for habitat diversification, fish breeding, watering of green areas, bathing during summer, and as water sources for grazing animals.
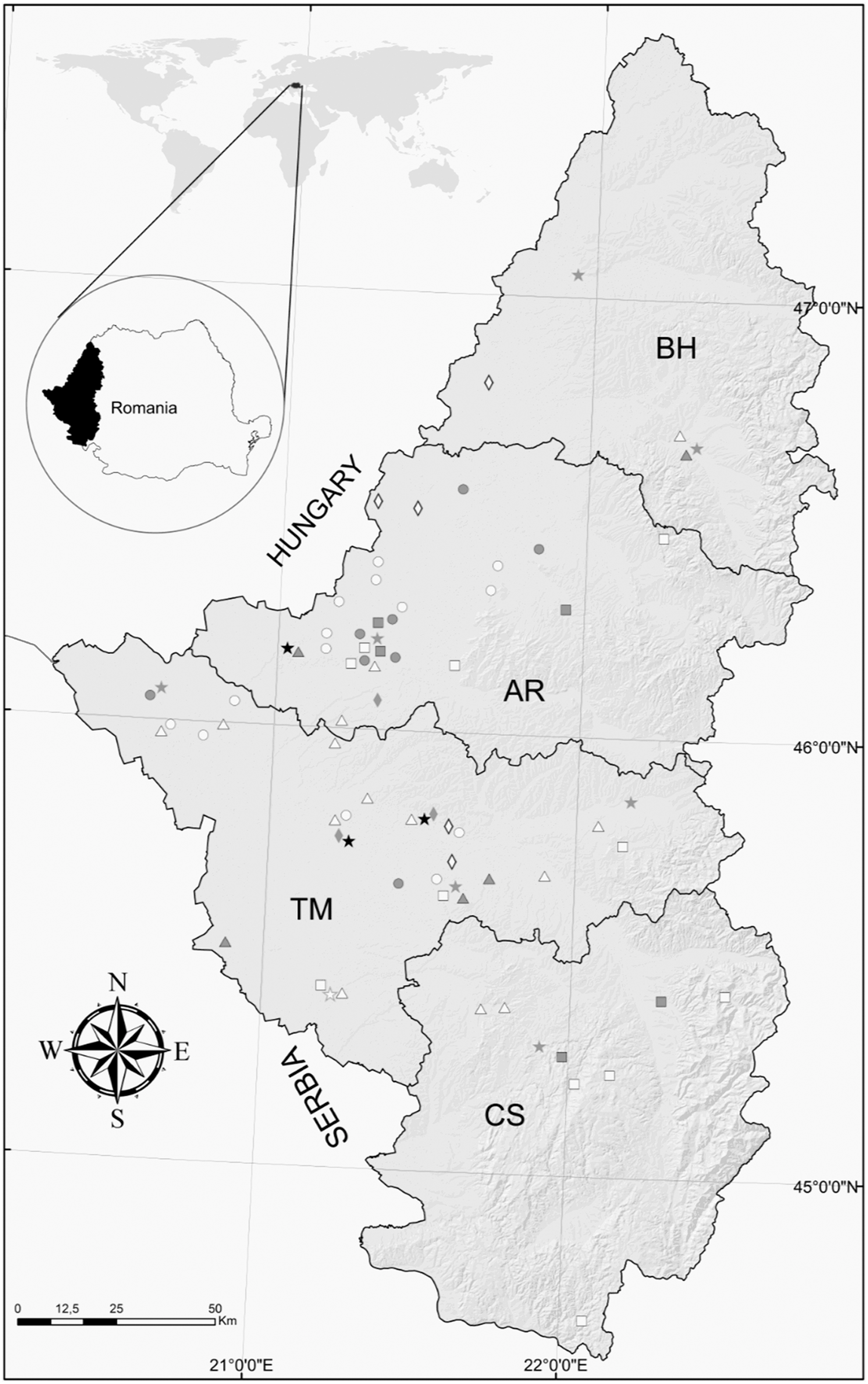
Map of western Romania indicating the approximate positions of the sampling sites, according to the type of water source tested (star, sewage; triangle, brook; rhombus, irrigation channel; square, lake; and circle, pond) in each county (AR-Arad; BH-Bihor; CS-Caraș-Severin; TM-Timiș). White-filled geometric symbols are negative results; gray-filled symbols are sites in which Giardia spp. were confirmed by microscopic and/or molecular analysis; black-filled geometric figures are sites in which mixed Giardia spp. and Cryptosporidium spp.-positive samples were identified microscopically and/or molecularly. Figure 1 can be viewed in greater detail online at
Sample collection and analyses
From January to November 2016, raw surface water samples (n = 76) were collected by microfiber filtration directly from the different water bodies using polyester microfiber filters (ARAD Hungária Kft., Miskolc, Hungary) with a pore size of 2 μm and then concentrated as detailed elsewhere (Plutzer et al. 2010). The tested samples included effluents of the wastewater treatment plants (n = 11) (average volume [AV] = 24.7 L, min–max 2.5–60 L), brooks (n = 19) [AV = 22.9 L, min–max 8–53 L], lakes (n = 16) [AV = 26.9 L, min–max 11–49 L], ponds (n = 22) [AV = 12.6 L, min–max 5–31 L], and collector channels (n = 8) [AV = 28.4 L, min–max 6–48.5 L]. The filtered water volume depended on water turbidity.
The recovered water concentrates (∼8 mL) were subjected to Cryptosporidium and Giardia (oo)cyst detection and identification, following the U.S. Environmental Protection Agency Method 1623 (U.S. Environmental Protection Agency [U.S. EPA] 2005). Therefore, the pathogens were detected through immunomagnetic separation using a Dynabeads™ GC-Combo Kit (Thermo Fisher Scientific Baltics UAB, Vilnius, Lithuania), as per the manufacturer's instructions. Subsequently, half (50 μL) of the resulted final concentrated pellet was transferred to the wells of microscope slides and scanned using immunofluorescence microscopy (IF) at a magnification of 400× (Motic-031; Miotic Incorporation LTD., Wetzlar, Germany). The slides were scanned for the presence and the number of (oo)cysts after staining with fluorescently labeled (FITC) monoclonal antibodies (Merifluor®, Cryptosporidum/Giardia; Meridian Bioscience, Inc., Cincinnati, OH), according to the manufacturer's protocol.
All samples were subjected to molecular analysis. Genomic DNA extraction from the second half (50 μL) of the concentrated water pellet was performed using an Isolate Genomic DNA Kit (Bioline Reagents Limited®, London, United Kingdom), according the manufacturer's recommendations. Molecular detection of G. duodenalis was accomplished by nested PCR of a fragment of the glutamate dehydrogenase (gdh) gene (∼432 bp). The specific primers and cycling parameters were previously reported (Read et al. 2004).
Cryptosporidium spp. were identified by a nested PCR amplification of the small subunit (SSU) rRNA gene (∼830 bp) and a restriction fragment length polymorphism (RFLP) analysis of the final nested PCR products with VspI (Promega®, Madison, WI) and SspI (New England Biolabs®, Beverly, MA) restriction enzymes. Except for the volume of the PCR reaction (25 μL instead of 100 μL), the primers, annealing temperatures, and digestion conditions were adopted following those in previous publication (Xiao and Ryan 2008). For the subtyping, the C. parvum diagnosed isolates were nested-PCR amplified targeting the 60 kDa glycoprotein (GP60) gene as previously described (Xiao and Ryan 2008). Positive and negative controls were included in each amplification. The secondary PCR and RFLP products were visualized on Midori Green™ (Nippon Genetics®; Europe Gmbh) stained 2.2% agarose gel.
G. duodenalis genotyping and C. parvum subtyping were performed through bidirectional sequencing of the positive gdh and GP60 gene amplicons, respectively, by the Macrogen Europe® Company (Amsterdam, the Netherlands). Genotypes and subtypes were identified by comparison with the GenBank® retrieved reference sequences using the online version of BLAST software (available
The results were statistically interpreted using the nonparametric Pearson's Chi–square test (Excel 2007; Microsoft, Redmond, WA). A p value ≤0.05 was considered statistically significant.
Results
The results are summarized in Table 1.
N.S., could not be sequenced; RFLP, restriction fragment length polymorphism; SD, standard deviation; IFA, immunofluorescence assay.
A total of 31 (40.8%; 95% confidence interval [CI] 29.8–52.6%) of the 76 collected water samples were Giardia spp. positive. This pathogen was registered in all tested water types. Cryptosporidium spp. were detected in only 3 (4.0%; CI 1.0–11.8%) sewage water samples in association with G. duodenalis. The cumulative detection rate for G. duodenalis was significantly higher (p < 0.05) in the effluents of wastewater treatment plants (10/11; 90.1%) than that in brooks (5/19; 26.3%; p = 0.0022), irrigation channels (3/8; 37.5%; p = 0.0267), lakes (5/16; 31.2%; p = 0.0057), and ponds (8/22; 36.4%, p = 0.0065). Except for the water samples from irrigation channels, the frequency of detection rates were different between the two diagnostic techniques (IFA and PCR) (Tables 1 and 2), which indicated that certain of the microscopically positive Giardia water samples exhibited negative PCR results and vice versa (Table 2; kappa agreement = 0.538). No correlations were found between the occurrence of negative or positive results and the filtered water volumes. The highest mean concentration of Giardia cysts per liter was in the effluents of wastewater (1.96 ± 30.07), and the lowest concentration was in lake water (0.21 ± 0.18; Table 1).
PCR and sequencing of the G. duodenalis gdh gene was successful in 22 samples, which showed a different genotype distribution according to the tested water source (Table 1). Assemblage E was detected most frequently (12/22, 54.5%) and was recovered from all tested water types. Nine samples (9/22, 40.9%) contained assemblage A subassemblage II, with dominant occurrence in sewage water; and another isolate (4.5%) from pond water was identified as assemblage D (Table 1). Genotyping was unsuccessful for one G. duodenalis isolate because primer mismatches occurred within the gdh gene fragment.
The nested PCR-RFLP procedure targeting the SSU rRNA gene revealed a single Cryptosporidium species, which was the zoonotic C. parvum in the positive microscope samples (kappa agreement = 1). In subsequent GP60 subtyping, the isolates were identified as the IIaA15G2R1 (n = 2) and IIdA18G1 (n = 1) subtypes.
Discussion
The present study demonstrated the presence and the first evidence of G. duodenalis in wastewaters and other surface water sources, including brooks, lakes, ponds, and irrigation channels, in Romania. In addition, the first molecular characterization of wastewater origin zoonotic C. parvum was performed in this study.
Of the five types of water studied, the contamination level of wastewaters with Giardia spp. was the highest significantly (p < 0.05). This result is not surprising, because wastewater discharges (effluents) are major environmental pollutants for this protozoan worldwide (Nasser et al. 2017). Furthermore, the dominance of the human-derived subassemblage AII (66.6%) supports our assumption based on a previous survey targeting western Romanian rivers in the same region (Imre et al. 2016) that effluents of the wastewater treatment plants are the primary Giardia contaminants of the great water courses. The ruminant origin assemblage E might have originated from the wash water discharged by the highly condensed livestock farms of the area. Although the wastewater treatment plants receive a preponderance of human origin sewage, the human-adapted C. hominis was not detected. However, the identification of C. parvum (involved in the zoonotic transmission of cryptosporidiosis) demonstrated that the effluents of the treatment plants play an important role in the environmental dispersion of this zoonotic species. Data available on the molecular epidemiology of cryptosporidiosis in the screened region indicate the exclusive presence of this species in humans (Vieira et al. 2015), cattle (Imre et al. 2011, Vieira et al. 2015), and pigs (Vieira et al. 2015) as well as dominant occurrence in lambs (Imre et al. 2013), highlighting that these hosts can contribute to environmental pollution with oocysts. Of note, the IIaA15G2R1 C. parvum subtype was previously identified as the primary pathogen in cattle (Imre et al. 2011), whereas the IIdA18G1 subtype has never been identified in Romania.
In the monitored brooks that predominantly cross rural landscapes, both G. duodenalis subassemblage AII and assemblage E were recorded, which suggested that the cysts could originate from human and domestic ruminant infections, most likely discharged through the emptying of loaded latrines or livestock runoff from unhygienic backyards. Similar to our results, the subassemblage AII responsible for anthroponotic transmission has been recorded in brook waters of neighboring Hungary (Plutzer et al. 2008). Therefore, these contaminated brooks constitute a potential threat for public health, particularly when the nearest household wells and springs that are used for drinking water are under their influence; further studies are required to clarify this assumption (Hijnen et al. 2005).
Consistent with the results obtained for the water types discussed above, the Giardia assemblages in irrigation channels (Table 1) indicated human and ruminant origins of the cysts. The canals are likely polluted with the nonzoonotic assemblage E during heavy rainfall events and receive transmissible forms of the parasite in water excess with diluted slurry from fertilized agricultural lands or directly from animal excreta from the highly condensed ruminant livestock farms of the region. The subassemblage AII indicated a preponderance of human pollution sources, which likely include the frequently observed practice of using the canals as wastewater collection facilities for households that are not connected to the sewer system. The results of Diallo et al. (2008) in a study aimed at detecting waterborne pathogens in a canal network were similar. Therefore, the results highlight a potential public health threat, particularly during the summer season when the population uses the canal waters for different purposes. For example, both bathing and irrigation of vegetables grown for human consumption or salad crops eaten uncooked can result in accidental ingestion of the infective cysts (Diallo et al. 2008).
Similar to the findings of various authors (Coupe et al. 2006, Yvonne et al. 2009), a significant risk of exposure to infection with Giardia cysts was identified during recreational activities in the screened lake waters (e.g., swimming, scuba diving, and jumping) or when these water bodies served as the drinking water supply for the surrounding human settlements. The recorded subassemblage AII suggested that prevention measures be implemented to avoid contamination by human waste, most likely from houseboats and from the locations near the lake densely inhabited during weekend outings. In addition, the assemblage E in the tested lakes could result from direct defecation during watering of sheep flocks and/or pasture cows or from wildlife as previously demonstrated by Györke et al. (2016) in Romania.
The investigated ponds were the only surface water type that tested negative for the human-infective subassemblage AII. The recorded assemblages (E and D) might originate from several domestic and wildlife animal sources, with the most probable scenarios direct defecation by grazing livestock and domestic or wild canids during watering. Notably, except for the wastewaters, Cryptosporidium was not detected in any other tested surface water type, although large-scale occurrence has been previously demonstrated in different screened hosts (Imre et al. 2011, 2013, Vieira et al. 2015) as potential environmental contaminants of the monitored area. However, considering the limited number of the processed samples, further investigations enrolling a statistically representative sample size are necessary to clarify the really low Cryptosporidium occurrence of the environmental samples.
As has been previously observed by various authors (Plutzer et al. 2008, Lobo et al. 2009, Efstratiou et al. 2017), the current survey highlighted a discrepancy between IFA and PCR in the detection of Giardia spp. (Table 2). This disagreement can be derived from the fact that different aliquots of the final concentrated pellet have been used within the diagnostic tests for each processed sample. In addition, the possible presence of PCR inhibiting substances (e.g., calcium ions, urea, phenol, ethanol, tannic acid, and proteinases) (Schrader et al. 2012) in the analyzed samples can markedly influence the assay sensitivity, resulting in the occurrence of false negative results. Taking these considerations into account, the results suggest that these diagnostic techniques are complementary and that further harmonization and improvement in the sensitivity and specificity remain for the correct evaluation of the occurrence of these protozoa in aquatic environmental samples.
Conclusions
The results of this pioneering study provide baseline information on the occurrence and molecular characterization of Giardia and Cryptosporidium in treated wastewaters and different surface water sources, including brooks, irrigation channels, lakes, and ponds, in western Romania. The survey showed that, except for ponds, all tested water types harbored Giardia and/or Cryptosporidium (oo)cysts with human-infective potential, representing a potential threat to public health. In the lack of routine monitoring of these waterborne pathogens in any aquatic system in Romania, the obtained results highlighted that the implementation or at least improvement of measures that provide protection from human fecal contamination of natural surface water bodies are recommended. In addition, the increasing of the awareness of the authorities responsible for recreational and drinking water quality can contribute significantly to mitigation of the highest Giardia incidence rate reported by the Romanian public health laboratories within the countries of the Eastern European Union over the last 5 years (summarized unpublished data). Likewise, further epidemiological studies focused on the environmental origin of (oo)cysts and testing their infectivity and viability, in addition to their occurrence in potable water, are recommended to correct the assessed risks.
Footnotes
Acknowledgments
This work was supported by a grant from the Romanian National Authority for Scientific Research and Innovation, CNCS–UEFISCDI, and project number PN-II-RU-TE-2014-4-1300. The authors appreciate very much the indispensable contributions of Eng. Anca Deac, Dr. Florin Hora, Dr. Corina Badea, and Dr. Morar Doru during sample collection and processing. We thank the water authorities for their cooperation and technical assistance.
Author Disclosure Statement
No competing financial interests exist.