
Abstract
Tick-borne encephalitis (TBE) is a zoonotic flaviviral infection that is a growing public health concern in European countries. The aims of this research were to detect and characterize tick-borne encephalitis virus (TBEV) in Ixodes ricinus ticks at presumed natural foci in Serbia, and to determine seroprevalence of TBEV IgG antibodies in humans and animals. A total of 500 I. ricinus ticks were examined for the presence of TBEV by real-time RT-PCR, and conventional nested PCR and sequencing. To determine TBEV seroprevalence, 267 human sera samples were collected, as were 200 sera samples from different animal species. All sera samples were examined by ELISA for the presence of anti-TBEV antibodies. To exclude cross-reactivity, all sera samples were tested for anti-West Nile virus (WNV) antibodies and all human sera samples were also tested for anti-Usutu virus antibodies by ELISA. Results of this preliminary study indicated TBEV activity in Serbia at two microfoci. Several decades after the previous documentation of TBEV in Serbia, we have demonstrated the presence of TBEV in I. ricinus questing nymphs (prevalence 2% and 6.6% at the two different localities) and anti-TBEV antibodies in humans (seroprevalence 0.37%). Moreover, we show for the first time TBEV seroprevalence in several animal species in Serbia, including dogs (seroprevalence 17.5%), horses (5%), wild boars (12.5%), cattle (2.5%), and roe deer (2.5%). None of the goats tested was positive for anti-TBEV IgG antibodies. TBEV isolate from I. ricinus tick in this study belonged to the Western European subtype. To understand the true public health concern in Serbia, detailed epidemiological, clinical, virological, and acarological research are required. This is important for implementation of effective control measures to reduce the incidence of TBE in Serbia.
Introduction
T
The first TBEV in the former Yugoslavia was isolated in Slovenia in 1953 from the blood of infected human patients, and in 1954, TBEV was found in Ixodes ricinus ticks (Vesenjak-Zmijanac et al. 1955, Kmet and Likar 1956). Later, a number of TBE foci were registered in the western part of the former Yugoslavia, whereas in Serbia, such foci were not found. However, results from seroepidemiological studies of TBEV infections in humans conducted between 1962 and 1969 and the isolation of virus from Ixodid ticks from the area of Pešter, Raška in 1972, which was identified as the Western European TBEV subtype, clearly showed the existence of active TBEV foci in Serbia (Bordoski et al. 1972).
Although the first seroepidemiological studies of TBEV infections in humans were conducted nearly fifty years ago, there have been no such studies in Serbia. Süss wrote that a few TBE cases were reported in an area near Belgrade, but there is no published information concerning these cases (Süss 2011). In Serbia, from 2004 through 2015, 16 cases of human TBE infections were reported to the Institute of Public Health of Serbia as required by mandatory reporting policies (Institute of Public Health of Serbia, 2004–2015). Thus, the significance of TBE in Serbia and its regions requires evaluation due to the positive preliminary serological findings in individuals living in the Autonomous Province of Vojvodina (Hrnjakovic Cvjetkovic et al. 2016).
In Western Europe, TBEV is transmitted primarily by I. ricinus ticks (Gritsun et al. 2003b), which are the dominant naturally occurring tick species in Serbia (Savic et al. 2010, Jurisic et al. 2010). According to previous studies in Serbia, the following pathogens were registered in I. ricinus ticks: Borrelia burgdorferi s.s., Borrelia lusitaniae, Borrelia afzelii, Borrelia valaisiana, Borrelia garinii, Borrelia miyamotoi, Anaplasma phagocytophilum, Anaplasma ovis, Coxiella burnetii, Candidatus Neoehrlichia mikurensis, Rickettsia monacensis, Rickettsia helvetica, Francisella tularensis subsp. tularensis, Hepatozoon canis, Babesia venatorum, and Babesia microti (Milutinovic et al. 2008, Tomanovic et al. 2013, Potkonjak et al. 2016a, b). However, there are no new data concerning the presence of TBEV in I. ricinus ticks.
Animals infected with TBEV are generally asymptomatic, but many studies have demonstrated the presence of circulating antibodies to TBEV in animal populations in Europe (Mansfield et al. 2009). To our knowledge, seroepidemiological studies of TBEV infections in animals have not been conducted in Serbia.
The aims of this research were to detect and characterize TBEV in I. ricinus ticks at presumed natural foci in Serbia, and to determine seroprevalence of IgG antibodies to TBEV in humans and animals.
Materials and Methods
Tick collection and identification
Questing ticks were collected from the vegetation during 2014 and 2015, using the “Flag-hour” method (Maupin et al. 1991), at two localities (S1, S2) around the Fruška Gora mountain in the Autonomous Province of Vojvodina and 15 localities (S3–S17) in Belgrade, Serbia. Location of tick sampling areas is shown on a map created using ArcGis/ArcMap (Fig. 1 and Table 1). Tick species were identified according to morphological keys (Nosek and Sixl 1972, Walker et al. 2003, Estrada-Peña et al. 2004). A total of 500 I. ricinus ticks were collected (50 nymphs from the Autonomous Province of Vojvodina and 450 nymphs from the Belgrade area). The ticks were kept at −86°C until further analysis.
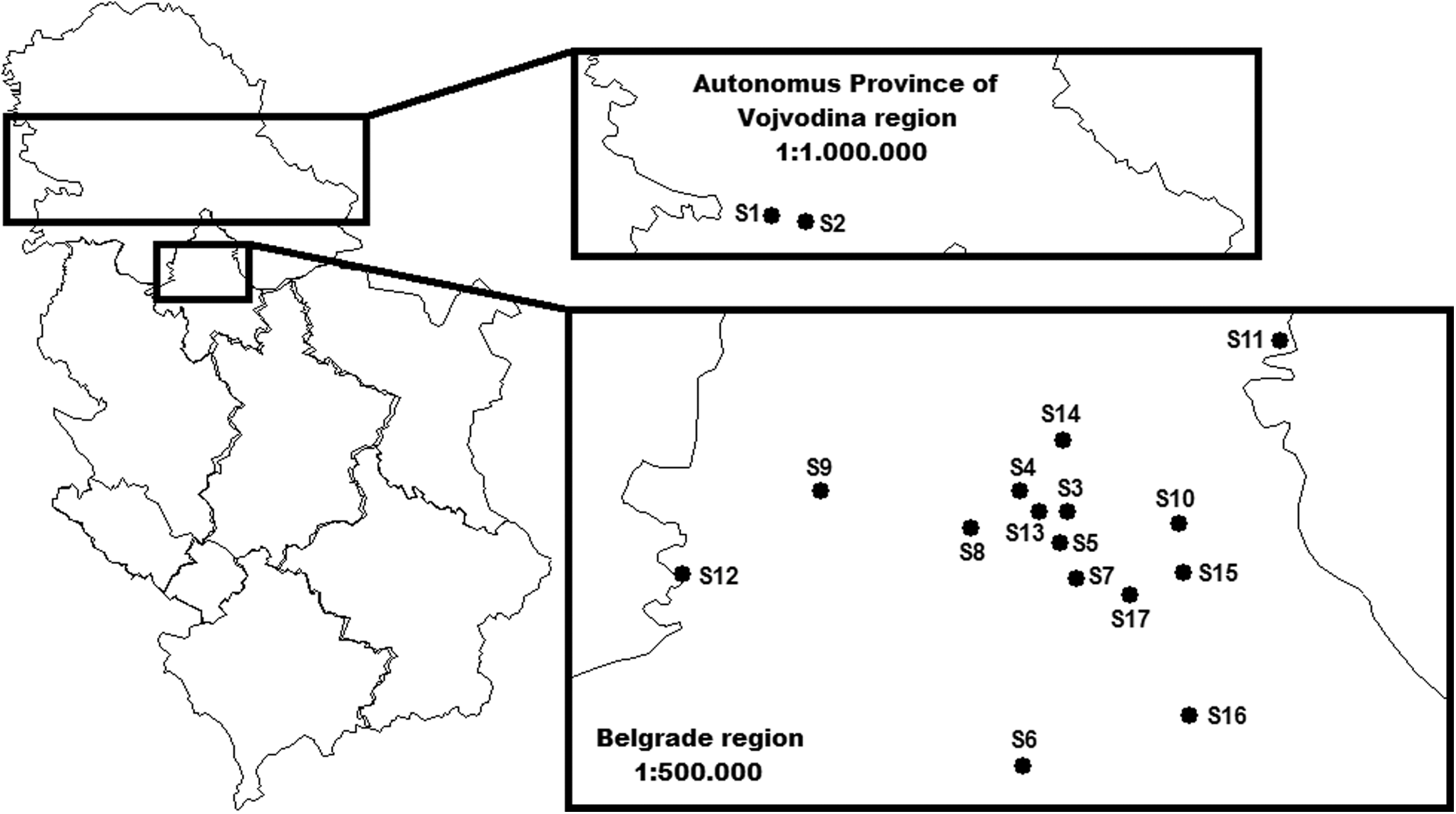
S1–S17, location of tick sampling areas in Serbia.
DD, decimal degrees; GPS, Global Positioning Systems.
Extraction of RNA from I. ricinus ticks
Individual I. ricinus ticks were placed in 2 mL microtubes and homogenized in 1 mL sterile phosphate-buffered saline for 5 min using a TissueLyser LT (Qiagen, Hilden, Germany) operating at 50 Hz. The homogenates were then centrifuged for 5 min at 2000 × g and 250 μL of supernatant was reserved for RNA extraction. Viral RNA was extracted using TRI Reagent® Solution (ThermoFisher Scientific, San Jose, CA) according to the manufacturer's instructions.
Real-time RT-PCR
Prepared samples were tested for the presence of TBEV RNA by in-house TaqMan-based one-step RT-qPCR to amplify part of the TBEV E gene using primers and probes that were described previously (Gaumann et al. 2010).
Briefly, one-step RT-qPCR was conducted using the commercial kit RNA UltraSense™ One-Step qRT-PCR System (Life Technologies Corporation, Carlsbad, CA) with primers (forward TBEE-F6: 5′-GGCTTGTGAGGCAAAAAAGAA-3′ and reverse TBEE-R2: 5′- TCCCGTGTGTGGTTCGACTT-3′) and probes (TBEE-P4: 5′-FAM-AAGCCACAGGACATGTGTACGACGCC-BHQ-1-3′) that target the region of the TBEV genome encoding the E protein. Each reaction contained 15 μL of reaction mix containing 1 × RNA UltraSense reaction mix, 20 μM of each primer, 10 μM of TBEE-P4 probe, 1 × ROX reference dye, and 1 μL of RNA UltraSense enzyme mix. Five microliters of nucleic acid extract sample was added to yield a final reaction volume of 20 μL. The thermocycling conditions were 15 min at 50°C and 2 min at 95°C, followed by 50 cycles of 15 s at 95°C and 40 s at 60°C.
Conventional nested PCR
For additional confirmation of TBEV and subsequent sequencing, conventional gel-based nested PCR was performed as described previously (Schrader and Süss 1999, Frimmel et al. 2014). For the first RT-PCR amplification step, a commercial OneStep RT-PCR kit (Qiagen GmbH, Hilden, Germany) was used with primers that amplified a fragment of the 5′NCR and partially covered the gene encoding the TBEV capsid protein (Pp1: 5′-GCG TTT GCT TCG GAC AGC ATT AGC-3′ and Pm1: 5′-GCG TCT TCG TTG CGG TCT CTT TCG-3′). Briefly, the reaction was performed in a total volume of 25 μL that included 12 μL nuclease-free water, 5 μL 5 × PCR buffer, 1 μL dNTP mix (containing 10 mM of each dNTP), 1 μL of a 20 μM solution of each primer, 1 μL of one step RT-PCR enzyme mix, and 4 μL of RNA template.
The RT-PCR protocol comprised reverse transcription at 50°C for 30 min followed by an initial denaturation at 95°C for 15 min. After this step, a PCR consisting of 40 repeated cycles of denaturation at 94°C for 30 s, annealing at 65°C for 60 s, and elongation at 72°C for 60 s was carried out. The reaction was finalized with extension at 72°C for 10 min.
The second PCR amplification was carried out with 2 μL of the first amplification product using the HotStarTaq Master Mix Kit (Qiagen GmbH) according to the manufacturer's instructions. Briefly, the amplification reaction was conducted in a volume of 25 μL containing 12.5 μL of master mix and 25 pmol of both internal primers (Pp2: 5′-TCG GAC AGC ATT AGC AGC GGT TGG-3′ and Pm2: 5′-TGC GGT CTC TTT CGA CAC TCG TCG-3′) that amplify a 178 bp fragment of the 5′NCR/Capsid (Core) gene from the TBEV genome. Amplification conditions were as follows: 95°C for 15 min, 35 cycles of 95°C for 30 s, 65°C for 30 s, and 72°C for 30 s, and a final extension at 72°C for 10 min.
Sequencing and phylogenetic analysis
All PCR products were analyzed by gel electrophoresis in 1.5% agarose and ethidium bromide staining. Specific bands were excised and purified using a QIAquick Gel Extraction Kit (Qiagen GmbH) according to the manufacturer's instructions. Regarding the amount of DNA in the PCR products, sequencing of selected PCR amplicons was performed in both directions using a Big Dye Terminator v3.1 Kit (Applied Biosystems, Foster City, CA) according to the manufacturer's instructions and both sets of primers (external and internal pairs of primers) in a 3130 Genetic Analyzer (Applied Biosystems).
BLAST (
Collection of human sera samples and flavivirus serology
Between January 1, 2014, and October 31, 2015, 267 sera samples from patients were collected at the Clinical Centre of Vojvodina, Clinic for Infectious Diseases, a tertiary care university hospital in Novi Sad, Vojvodina, Serbia. The study protocol received approval from an institutional review board and the study subjects provided informed consent. None of the patients had obtained a vaccination against TBE, yellow fever, or dengue fever. All sera samples were tested using a commercial indirect ELISA kit (Euroimmun, Lübeck, Germany) for anti-TBEV IgG antibodies, and were also tested with a commercially available indirect ELISA kit (Euroimmun) for anti-West Nile virus (WNV) IgG antibodies and anti-Usutu virus (USUV) IgG antibodies to exclude cross-reactivity within the sera samples that were positive for anti-TBEV IgG antibodies.
Collection of animal sera samples and flavivirus serology
Between January 1, 2014, and October 31, 2015, 200 sera samples from various animals (40 dogs, 20 horses, 40 wild boars, 40 cattle, 40 roe deer, and 20 goats) were collected at the Scientific Veterinary Institute “Novi Sad”, Novi Sad, Serbia. All sera samples were tested for anti-TBEV IgG antibodies with a commercial indirect ELISA kit for all species (ProgenBiotechnikGmbH, Heidelberg, Germany), and were also tested with a commercial blocking ELISA kit (INGEZIM WEST NILE COMPAC, Ingenasa, Madrid, Spain; for blood sera samples from horses) and a commercial competitive ELISA kit (ID Screen West Nile Competition Multi-species, IDvet, Montpellier, France; for blood sera samples from dogs, wild boars, cattle, roe deer, and goats) for anti-WNV IgG antibodies, as well as to exclude cross-reactivity of sera samples that were positive for anti-TBEV IgG antibodies.
Results and Discussion
Presence of TBEV in I. ricinus ticks
The presence of the Western European TBEV subtype was detected in I. ricinus ticks using real-time RT-PCR and nested PCR methods, and was also characterized at a molecular level by sequencing. The TBEV Western European subtype (NCBI GenBank accession number MF497317) was detected in 1 of 50 (2%) I. ricinus ticks from one locality near the Fruška Gora mountain in the Autonomous Province of Vojvodina (Andrevlje, S1 on Fig. 1). In I. ricinus ticks from one Belgrade locality (Manastirska šuma-Rakovica, S7 on Fig. 1), 30 out of 450 (6.6%) ticks were positive for TBEV.
More than 14 species of ticks can be infected by TBEV, but I. ricinus ticks are the principal vector of the Western European subtype (Dumpis et al. 1999). The TBEV prevalence of infected ticks varies between regions (Valarcher et al. 2015) and in Europe, the prevalence in I. ricinus ticks varies between 0.1% and 5% in unfed ticks and increases with stage development (Süss 2011, Pettersson et al. 2014).
In Croatia, TBE virus was detected in adult I. ricinus and Ixodes hexagonus ticks with minimal viral prevalence of 1.6% (Jemersic et al. 2014). In Hungary, no TBEV in I. ricinus ticks was detected between 2006 and 2008 (Egyed et al. 2012). One year later, in the southeastern region of Hungary, I. ricinus was the only species to test positive for TBEV and there was minimal prevalence (0.08%) in unfed nymphs (Pinter et al. 2013). In Slovenia, the overall infection rate in I. ricinus nymphs was 0.37% (Durmisi et al. 2011), whereas in Czech Republic, a very low overall TBEV prevalence of 0.096% in I. ricinus nymphs was registered (Daniel et al. 2016). In Poland, the minimal infection rate of ticks with TBEV in the total area examined was 0.96% (Biernat et al. 2014).
The minimal field infection rate was 4.4 virus-containing ticks out of 1000 collected ticks from Austria (Labuda et al. 1993), whereas later reports from Austria reported a prevalence of TBEV in ticks that ranged between 0% and 3% (Duscher et al. 2015a). In Germany, the TBEV prevalence rate in I. ricinus was 0.1% (Mehlhorn et al. 2016). Thus, the prevalence of TBEV in I. ricinus ticks in our research is higher than that for other reports from European countries. These fluctuations in TBEV tick infection prevalence are presumably due, in part, to the varying densities of reservoir-competent versus reservoir-incompetent tick hosts (Pettersson et al. 2014).
Our detection of TBEV-infected I. ricinus questing nymphs is the first positive result in Serbia since 1972 year and indicates that two localities (Andrevlje, Manastirska šuma-Rakovica) can be considered TBEV-positive microfoci. Both TBEV-positive microfoci are in forested areas where rodents are present. Climatic and ecological conditions, as well as cofeeding ticks on small mammal hosts, are known to determine TBEV transmission efficiency. Moreover, TBEV is known to occur in discrete foci of variable sizes (Andreassen et al. 2012). Indeed, these microfoci can sometimes be very small (Rieille et al. 2017). TBEV detection in questing ticks is not a sensitive indicator in risk assessment of TBE in humans; so other surveillance methods, such as wildlife or farm animal investigations, as was done in this study, have been suggested to gauge the seroprevalence of TBEV (Stefanoff et al. 2013).
Phylogenetic analysis of Serbian TBEV isolates
From all selected PCR amplicons for sequencing, a PCR amplicon of one TBEV isolate was successfully sequenced to generate a high-quality sequence for phylogenetic analysis.
The phylogenetic analysis of the 5′NCR/capsid (Core) protein nucleotide sequence showed that a Serbian TBEV isolate from an I. ricinus tick from Fruška Gora mountain (TBEV/FruskaGora2015/Srb; NCBI GenBank accession number MF497317), which was detected in this study, belonged to the Western European subtype of TBEV (TBEV EU). This Serbian TBEV isolate branched together with the Kumlinge A52 strain (1952) from Finland, the Salem (2007) and Leila-BH95/15 (2015) strains from Germany, the Neudoerfl strain (1988) from Austria, the Toro-2003 (2003) and Tick/Swe/Habo/2011/1 (2011) strains from Sweden, the Skrivanek (2014) strain from Czech Republic, and the Ljubljana I (1992) strain from Slovenia, which shared 100% homology in a 175 nucleotide region of the 5′NCR/capsid protein sequence (Fig. 2).
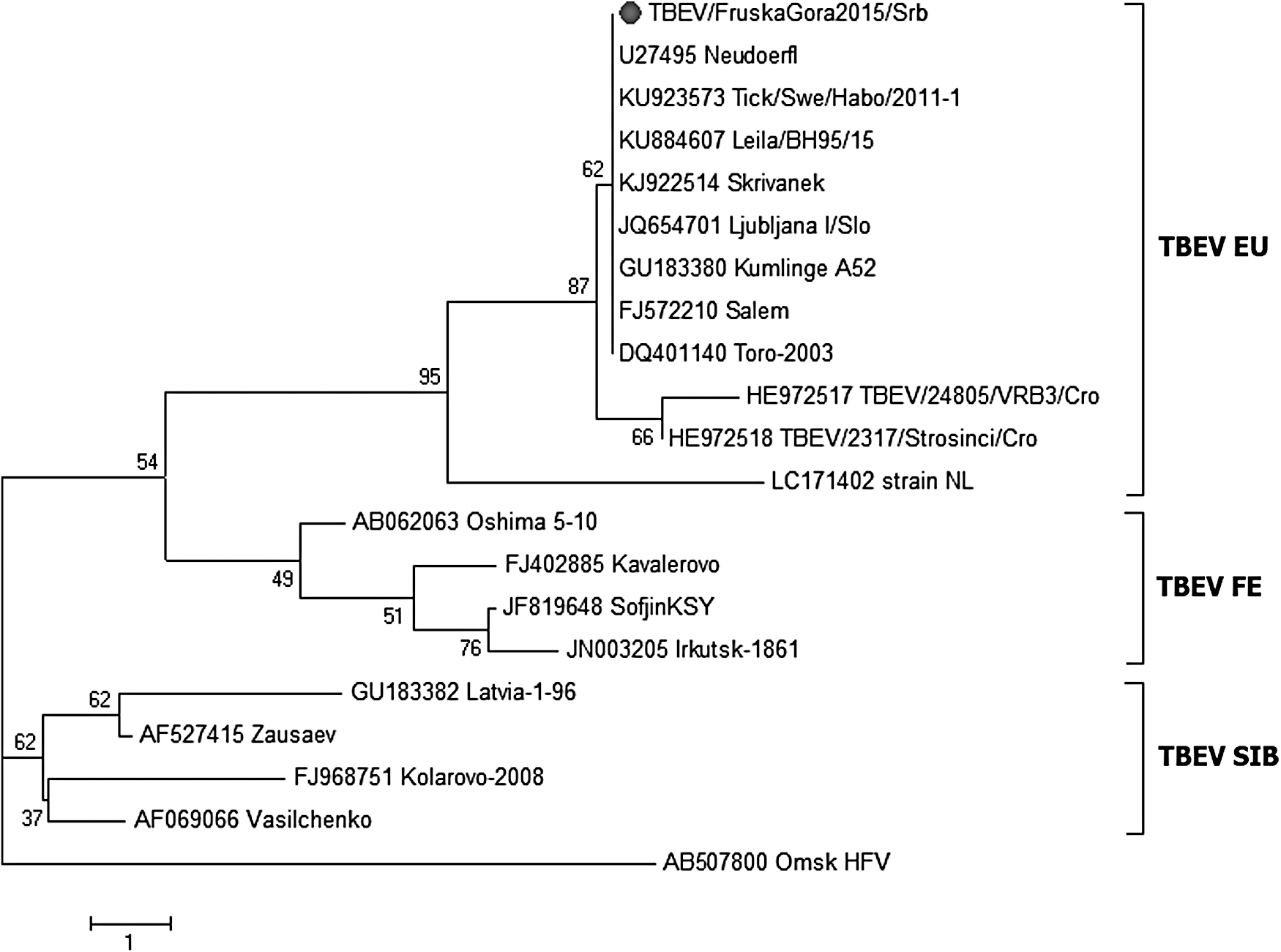
Phylogenetic tree constructed from 175 nucleotide (nt) sequence encoding the 5′UTR/Capsid (Core) from the Serbian TBEV/FruskaGora2015 isolate (NCBI GenBank® accession number MF497317) from an I. ricinus tick (marked with circle) and 19 nt sequences of TBEV strains obtained from NCBI GenBank. The tree was constructed using the neighbor-joining method based on bootstrapping of 1000 replicates (using MEGA version 6). The scale bar indicates the branch length based on the number of nucleotide substitutions per site and figures at branching points represent bootstrap values. Omsk hemorrhagic fever virus (Omsk HFV) was used for outgroup sequences. TBEV, tick-borne encephalitis virus; TBEV EU, Western European TBEV subtype strains; TBEV FE, Far Eastern TBEV subtype strains; TBEV SIB, Siberian TBEV subtype strains.
There was 99% sequence similarity between the Serbian TBEV isolate and TBEV/24805-VRB3/Cro (2011) and TBEV/2317-Strosinci/Cro (2012) strains from neighboring Croatia belonging to the same TBEV EU subtype, but the similarity to another TBEV EU subtype, strain NL (2015) from the Netherlands, was only 96% (Fig. 2).
Compared to the strains Zausaev (2002) from a patient in Siberia and the Kolarovo-2008 (2008) strain from Russia, the Serbian TBEV isolate TBEV/FruskaGora2015/Srb showed 96% and 92% sequence similarity, respectively. There was 94% similarity between our Serbian isolate and Far Eastern TBEV subtype strains and 92% similarity with the strain Oshima_5_10 (2001) from Japan and the Irkutsk-1861 (2008) strain from the eastern part of Russia (Fig. 2).
Human sera samples and flavivirus serology
Anti-TBEV IgG antibodies were found in six human sera (6/267; 2.24%) samples, but in five of these samples, anti-WNV IgG antibodies and/or anti-USUV IgG antibodies were also found. Due to this cross-reactivity between members of the Flaviviridae family, these five sera samples were excluded from the overall TBEV seroprevalence results and further interpretation. As such, the actual seroprevalence in humans in our sample was 0.37%. According to preliminary serologic investigations conducted in Serbia from 2012, a low prevalence of anti-TBEV IgG was detected in the South Bačka District of the Vojvodina Province in the northern region of Serbia, and no seropositive individuals were detected in the Nišava district in the southern region of Serbia (Hrnjakovic Cvjetkovic et al. 2016).
This seroprevalence rate is lower than that seen for other areas of Europe. For example, the seroprevalence of TBEV in exposed forest workers in eastern France was 3.4% (Thorin et al. 2008), whereas in Poland, TBEV seroprevalence in nonvaccinated humans from endemic regions was as high as 81% (Siennicka et al. 2010). In Lithuania, TBEV seroprevalence in healthy individuals from different parts of the country was 2.96% (Juceviciene et al. 2002). Meanwhile, Oehme et al. (2002) reported that in southwest Germany, anti-TBEV antibody prevalence in forestry workers was up to 27% in areas with no reports of clinical cases. Again, although the seroprevalence obtained in this study is low relative to other European countries, this finding could reflect the lack of clinical cases in the investigation area.
Our results nonetheless confirm the presence of anti-TBEV antibodies in humans in Serbia, indicating that TBEV is likely circulating in the country.
Animal sera samples and flavivirus serology
Anti-TBEV IgG antibodies were found in serum samples from 7 dogs (7/40; 17.5%), 1 horse (1/20; 5%), 5 wild boars (5/40; 12.5%), 1 cattle (1/40; 2.5%), and 1 roe deer (1/40; 2.5%). None of the goats tested (0/20) was positive for anti-TBEV IgG antibodies. Since WNV is highly prevalent in Serbia (Djuricic et al. 2013, Escribano-Romero et al. 2015), animal sera were also tested for the presence of anti-WNV IgG antibodies to exclude cross-reactivity. Negative results for WNV were obtained for all animal sera samples.
The results of other seroepidemiological studies confirmed the findings of anti-TBEV antibodies in different animal species. In Germany, results from an investigation using roe deer as sentinels would eliminate the inherent bias of risk maps based on human incidence (Kiffner et al. 2012). Similar to our findings for anti-TBEV antibodies in roe deer, the seroprevalence was 2% in roe deer from the Netherlands (Jahfari et al. 2017) and 2.4% in roe deer from Austria, as indicated by immunofluorescent antibody test (Duscher et al. 2015b). However, Tonteri et al. reported that none of the 17 roe deer tested in Finland was seropositive for TBEV (Tonteri et al. 2016).
Noticeably higher seroprevalence for roe deer was reported in Denmark (8.7% of roe deer were seropositive for TBEV by hemagglutination-inhibition test) (Skarphedinsson et al. 2005) and Germany (8/35; 22.86% TBEV seropositive roe deer by serum neutralization test). The German study also reported 10.26% (190/1851) TBEV-seropositive wild boars (Balling et al. 2014). In Norway, 51 out of 317 (16.1%) domestic dogs had TBEV-specific IgG in sera (Csango et al. 2004). Seroprevalence in sheep and goats varies greatly among individual flocks to range from 0% to 58%, with the highest rates measured near recognized foci for TBEV transmission (Imhoff et al. 2015). In Austria, transmission of TBEV to six humans and four domestic pigs through consumption of unpasteurized goat milk was evidence for a changing TBEV epidemiology in central Europe (Holzmann et al. 2009).
Sheep and goats are optimal sentinels for TBEV because they are rather sedentary and stay on meadows that are owned or leased by the flock owner or on meadows used on agreed terms of a contract, for example, for rural conservation (Klaus et al. 2012). In our research, all 20 goat sera samples were negative, although an evaluation of a larger sample of goats in Serbia is needed. Meanwhile, a study conducted in southwestern Germany in districts that had varying TBE incidence found seroprevalence in cattle ranging between 2% and 10% (Leutloffa et al. 2006). In horses in TBEV-endemic regions in southern Germany and Austria, seroprevalence rates of up to 26% were reported (Klaus et al. 2013, Rushton et al. 2013).
Cross-reactivity with related Flaviviridae must be taken into consideration, as was done in this study, particularly for TBEV serology in regions with a high prevalence of other Flaviviridae family members (Klaus et al. 2014). Overall, our results confirm the presence of anti-TBEV antibodies in several animal species in Serbia, which, together with the results for humans, indicate the circulation of TBEV in Serbia.
Conclusions
Results of this preliminary study indicate TBEV activity in Serbia at two microfoci. Several decades after earlier documentation of TBEV in Serbia, we have demonstrated the presence of TBEV in I. ricinus questing nymphs and anti-TBEV antibodies in humans and, for the first time in Serbia, in different animal species. As TBE is an increasing public health concern in other European countries, to understand the true public health concern in Serbia, detailed epidemiological, clinical, virological, and acarological research are required. This is important for implementation of effective control measures to reduce the incidence of TBE in Serbia.
Footnotes
Acknowledgments
This work was supported by the Ministry of Science and Technological Development of the Republic of Serbia, grant TR 31084.
Author Disclosure Statement
No competing financial interests exist.