
Abstract
Cronobacter sakazakii has been implicated in causing serious infections in neonates due to consumption of contaminated infant powdered milk. The zoonotic potential of the organism was not clear due to scarce evidence about the role of food animals in the transmission of infection. C. sakazakii was identified in infant powdered milk (n = 100), infant stool (n = 100), and dairy animal feces (n = 100) with the percentages of 1%, 2%, and 4%, respectively. The outer membrane protein A (ompA) gene was characterized in all isolates of different origin, while gene encoding for zinc-metaloprotease (zpx) was only identified in isolates from animal feces. Genotyping of C. sakazakii isolates using enterobacterial repetitive intergenic consensus polymerase chain reaction revealed heterogenicity. The survival and thermotolerance of one potentially virulent C. sakazakii isolate of animal origin were examined at different temperatures. The isolate could survive with a stationary number at refrigeration temperature and the number increased significantly at room temperature after 24 h. The isolate showed thermoresistance when subjected to temperature range from 54°C to 64°C with D values ranged from 13.79 and 4.64 min and z value of 14.42. To the best of our knowledge, this is the first report of C. sakazakii isolation from buffalo feces in Egypt.
Introduction
G
Infant milk powder was implicated as the main source of Cronobacter resulting in 50–80% of infections (van Acker et al. 2001, Ye et al. 2010). However, various food and environmental samples have also been documented as sources of infection (Iversen and Forsythe 2004, Kandhai et al. 2004, 2010, Friedemann 2007).
Severe meningitis, meningoencephalitis, necrotizing enterocolitis, and sepsis caused by C. sakazakii have been reported in neonates consuming infant powdered milk (Caubilla-Barron et al. 2007). Moreover, surviving patients were reported to suffer severe neurological signs including hydrocephalus and developmental delay (Lai 2001). Infection in elderly immunocompromised individuals consuming contaminated milk powder have also been reported (Lai 2001, van Acker et al. 2001). Virulence associated genes in Cronobacter spp. have a role in the potential of these bacteria to produce pathogenic effect in infected persons. The outer membrane protein A associated gene (ompA) and the gene encoding for zinc-metaloprotease (zpx) are both considered important virulence factors (Mohan Nair and Venkitanarayanan 2006).
Various genotyping techniques such as random amplified polymorphic DNA (RAPD) and enterobacterial repetitive intergenic consensus polymerase chain reaction (ERIC-PCR) have been developed to trace the sources of C. sakazakii infection. ERIC-PCR has been developed as a rapid, easy, inexpensive, and potential method to study the epidemiology and genetic diversity of different Cronobacter species (Ye et al. 2008, 2010).
Although the number of isolated Cronobacter cells from dry infant powdered milk is low, reconstitution of milk formula is considered a risk that provides an opportunity to the existing organisms to grow, especially when prepared under unhygienic conditions (Al-Holy et al. 2009). Moreover, Cronobacter spp. are capable of rapid multiplication if kept for prolonged time in bottle heaters or at room temperature. Thermotolerance of Cronobacter species was reported to be higher than other Enterobacteriaceae, which may be contributed to their survival in dried products (Nazarowec-White and Farber 1997). However, variable thermal resistance of Cronobacter spp. has been reported (Al-Holy et al. 2009).
The main objective of this study is to investigate the occurrence of Cronobacter spp. in different samples including raw milk, milk powder, infant powdered milk, animal feces, and infant stool. Characterization of virulence associated genes and genotyping of the isolates were carried out. In addition, the survival and thermoresistance of C. sakazakii in reconstituted infant powdered milk were also investigated.
Materials and Methods
Sampling
One hundred raw milk samples were collected from individual lactating cows (n = 66) and buffaloes (n = 34) from five dairy farms (three cattle and two buffalo farms) at different localities in Sharkia Governorate, Egypt. Fecal samples (n = 100) were collected from the rectum of animals from which milk samples were obtained. The farms and animals were selected with respect to agreement of the farm owners to participate. Stool swabs (n = 100) were also collected from diarrheic (n = 32) and apparently healthy neonates (n = 68) at infant incubators at different hospitals (four hospitals). Stool samples were collected from the neonates with respect to the agreement of their parents and physicians. Twenty milk powder samples of different manufacturers and 100 infant powdered milk samples were purchased from different supermarkets and pharmacies within the same study area.
Informed consent for participation in the study was obtained from the parents of the neonates involved in this study. The study was approved by the Research Ethics Committee, Faculty of Veterinary Medicine, Zagazig University, Egypt.
Isolation and identification of Cronobacter species
Pre-enrichment of the samples was performed in buffered peptone water at 37°C for 24 h (ISO 2006). A portion (0.1 mL) of the culture was then transferred to 10 mL of Cronobacter screening broth (CSB 38948, Sigma) and incubated at 42°C for 24 h, and then examined for changing of color to yellow (Iversen et al. 2008). A loopful from the enriched CSB culture was streaked directly onto Cronobacter species isolation chromogenic agar (CSIA; Sigma, 14703; Hichrome CISA) and incubated at 44°C for 24 h.
Molecular identification of Cronobacter species
The DNA from biochemically suspected Cronobacter isolates was extracted using QIAamp DNA Mini Kit (Cat. no. 51304; Qiagen) according to the manufacturer's instructions. Amplification of 16S rRNA specific for Cronobacter spp. was carried out using conventional PCR (Hassan et al. 2007). Probe-based real-time PCR (rtPCR) was utilized for the identification of C. sakazakii by the amplification of cgcA gene (Hu et al. 2016). The virulence associated genes ompA (Mohan Nair and Venkitanarayanan 2006) and zpx (Kothary et al. 2007) were also identified by PCR. A positive control of C. sakazakii was kindly provided by the Biotechnology Unit, Reference laboratory for Veterinary Quality Control on Poultry Production, Animal Health Research Institute, Dokki, Giza, Egypt.
For further confirmation of C. sakazakii isolates identified by 16S rRNA PCR from buffalo fecal samples, sequencing of the amplified products was carried out. QIAquick Gel Extraction Kits (Qiagen, S. A. Courtaboeuf, France) were used for amplicon extraction from gel according to the manufacturer's guidelines. The purified products were sequenced with Big dye Terminator V3.1 Cycle Sequencing Kits (Perkin-Elmer, Foster City, CA) as described by the manufacturer. DNASTAR software (Lasergene version 7.2; DNASTAR, Madison, WI) was used to analyze two sequences that were then submitted to the GenBank, which provided the two accession numbers of KY210879 and KY210880. Identity of the two isolates with other C. sakazakii isolates on the GenBank was determined.
ERIC-PCR fingerprinting
Two primer sets with the sequences ERIC-DG111-F 5′-ATG TAA GCT CCT GGG GAT TCA C-3′and ERIC-DG112-R 5′-AAG TAA GTG ACT GGG GTG AGC G-3′ were used to amplify repetitive sequences contained in the chromosomal DNA of C. sakazakii isolates using multiple amplification profile in triplicate (Versalovic et al., 1991). Depending on the presence or absence of each band, a binary code was used to transform ERIC-PCR fingerprinting data. Unweighted pair group method with arithmetic average and sequential hierarchical and nested clustering routine were used to generate a dendrogram using SPSS, Inc. version 22 (IBM Corp. 2013, Armonk, NY). The Simpson's index of diversity was used to measure the discriminatory power of ERIC-PCR (Hunter, 1990). Discriminatory value of more than 0.9 indicates good differentiation.
Survival of C. sakazakii in reconstituted infant powdered milk
Water used to prepare infant powder was boiled and then allowed to gradually cool, the appropriate amount of powder was added according to the manufacturer. Reconstituted infant powdered milk (Aptamil) in sterile distilled water was carried out according the manufacturer's guideline. C. sakazakii isolated from buffalo feces during this study and proven to harbor ompA and zpx associated virulence genes was used for the survival and thermotolerance experiment. The inoculum was prepared using surface plate method (Thatcher and Clark 1968). The isolate was inoculated into CSB and incubated at 42°C for 24 h. Serial dilution of the original broth culture was performed and a portion of 100 μL from each dilution was then aseptically plated onto CSIA and incubated at 44°C for 24 h. The dilution that had a microbial load of 105 CFU/mL was used for the inoculation of the prepared reconstituted infant powdered milk. Inoculated infant powdered milk tubes were then stored at room and refrigeration temperature. Examination was carried out in triplicate at 0 time (15 min after inoculation) and after incubation at both storage temperatures for 4, 8, 24, 48, 72, and 96 h. Control groups of noninoculated infant powdered milk were also included. For quantification of bacteria at each storage time, three samples were collected to determine C. sakazakii count (CFU/mL). Bacterial count in triplicate is given as mean values and standard deviations (SD).
The difference between the refrigeration and ambient temperature storage groups was estimated using two-way ANOVA test (Factorial design) and least significant difference. The test results were calculated by SPSS version 22 (IBM Corp. 2013, Armonk, NY). Data were presented as mean ± SD and significance was considered at p < 0.05.
Thermotolerance of C. sakazakii
Thermotolerance of C. sakazakii was examined according to Al-Holy, et al. (2009). The reconstituted milk was prepared and inoculated with C. sakazakii as described in the survival experiment. Heat treatment was conducted in a water bath at 54°C, 56°C, 58°C, 60°C, and 64°C. At each heat treatment, 1 mL from the inoculated infant powdered milk was taken in triplicate at 0, 3, 6, 9, 12, 15, and 18 min. Quantification of the colonies was then performed using overlay method (Al-Holy et al. 2008). The decimal reduction time (D-value), the time required at certain temperature to reduce a specific microbial population by a factor of 10,and the z-value (number of degrees of temperature change necessary to change the D-value by a factor of 10) were determined as previously described (Al-Holy et al. 2009).
Results
The results in Table 1 show that out of 420 examined samples, 12 (2.9%, 95% CI: 1.6–5.1) were confirmed by PCR amplification of 16S rRNA sequence specific for Cronobacter species. Molecular identification of C. sakazakii using specific rtPCR targeting cgcA gene showed overall, seven (1.7%, 95% CI: 0.7–3.6) samples were positive. Sequencing of the amplified 16S rRNA fragment was done for two isolates of animal feces origin. Alignment of the two sequences showed 100% identity with C. sakazakii isolates on the GenBank.
Molecular amplification of ompA gene revealed that 8 out of 12 (66.7%, 95% CI: 35.4–88.7) Cronobacter isolates harbored ompA associated gene (2 isolates from infant powdered milk and 6 isolates from infant stool), while, only the 4 isolates of animal feces origin (33.3%, 95% CI: 11.3–64.6) were positive for zpx associated gene.
The banding patterns obtained by ERIC-PCR revealed multiple DNA fragments ranging in size between 101 and 3000 bp (Fig. 1). Six profiles were produced and they were referred to as E1 to E6 (Table 2). Simpson's index of diversity calculation revealed a high discriminatory index of ERIC-PCR (0.9524). The dendrogram analysis of seven C. sakazakii isolates of infant powdered milk, animal and human origins showed one cluster (contained three isolates of animal feces origin) and four single isolates (Table 2 and Fig. 2).
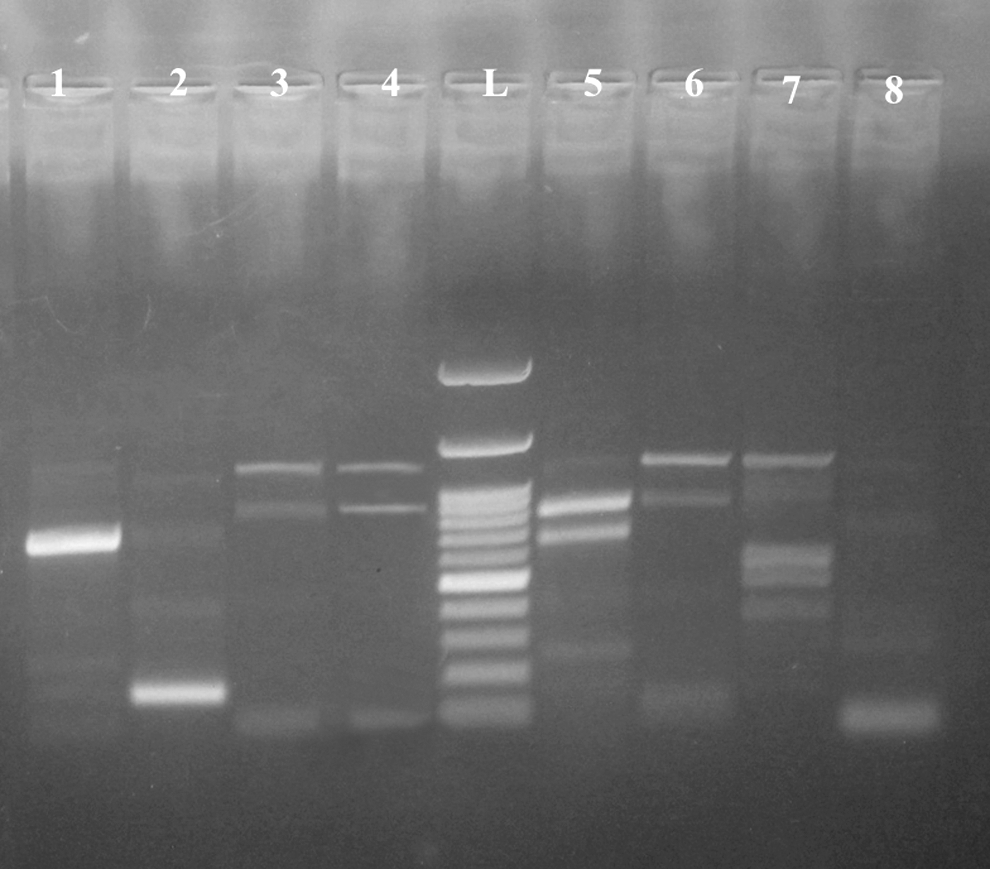
ERIC-PCR fingerprinting of C. sakazakii in 1.5% agarose: lane L, 100 bp DNA ladder; lane 1: infant stool (IS1), lane 2: infant stool (IS2), lanes 3–6, animal feces (AF3, AF4, AF5, AF6), lane 7 (infant milk powder; PIF) and lane 8: C. sakazakii positive control. ERIC-PCR, enterobacterial repetitive intergenic consensus polymerase chain reaction.

Dendrogram showing the relatedness of C. sakazakii isolates from PIF, animal feces and infant stool determined by ERIC-PCR fingerprinting using the SPSS computer software program (based on the presence or absence of each band at 12.5 linkage distance). PIF, powdered infant milk; AF, animal feces; IS, infant stool.
The concentration of C. sakazakii in infant powdered milk at refrigeration temperature (4°C) remained at the initial inoculum levels (5.176 CFU/mL) and did not decline or increase after incubation for 4, 8, 24, 48, 72, and 96 h (Table 3). At ambient temperature, there was no change in the number of colonies till 8 h postinoculation. A significant increase was observed after 24, 48, 72, and 96 h (p ≤ 0.05). The results in Table 4 show that D-values of C. sakazakii were 13.79, 9.13, 7.51, 5.74, and 4.64 min at 54°C, 56°C, 58°C, 60°C, and 64°C, respectively, while, the z-value was 17.42°C.
Means carrying different superscripts are significantly different at (p-value <0 .05), while means carrying same superscripts are not significantly different based on least significant difference.
Discussion
Identification of sources contaminated with Cronobacter spp. is important to elucidate the epidemiology of the disease. Out of 420 examined samples, 12 (2.9%) were confirmed by PCR as Cronobacter species, of the examined infant powdered milk samples; 2% were positive for Cronobacter species and only 1% was identified as C. sakazakii (Table 1). Nearly similar results of Cronobacter isolation were previously reported in infant powdered milk (Iversen and Forsythe 2004, Hoque et al. 2010, Heperkan et al. 2017). In Egypt, higher percentages of 13.3% (El-Sharoud et al. 2009) and 24% (El-Gamal et al. 2013) were reported. Infant milk powder are usually pasteurized during manufacturing; therefore, the final product could be contaminated from factories' environment, handling and adding heat sensitive nutrients after pasteurization (Kandhai et al. 2004).
In this study, none of the milk powder or raw milk samples were positive for the presence of Cronobacter spp. (Table 1). Likewise, different studies reported the same findings (Jaradat et al. 2009, Mozrová et al. 2014). However, the role of milk powder and raw milk as vehicles for transmission cannot be ruled out.
To understand the potential zoonotic role of animals in maintaining and disseminating Cronobacter species (especially C. sakazakii) in the environment, 100 fecal samples taken from 66 lactating cows and 34 buffaloes were examined. Six (6%) of the examined animal fecal samples (six buffalo feces) were positive for Cronobacter spp., of which 4% were confirmed as C. sakazakii by PCR (Table 1). Isolation of C. sakazakii from goat feces, pig feces, and chicken droppings has been recently reported (Mahindroo et al. 2016). Previous studies have reported that food production animals have no role in the carriage of Cronobacter species (Molloy et al. 2009, Mozrová et al. 2014). However, other studies documented the isolation of Cronobacter spp. from animals with no apparent signs of infection (Hamilton et al. 2003). The current results indicated that the presence of C. sakazakii in animal feces (buffalo feces) poses a major risk for contamination of the environment, thus highlighting the zoonotic potential of this pathogen. Moreover, the previous isolation of Cronobacter from a range of meat products such as sausage, camel, pork, and chicken indicated also the risk posed by these bacteria to public health (Friedemann 2007, Molloy et al. 2009). To the best of our knowledge, this is the first report of C. sakazakii isolation from buffalo feces in Egypt.
Isolation of Cronobacter spp. from different clinical samples such as cerebral spinal fluid, skin, respiratory tract, blood, wounds, stool, and urine has been reported (Lai 2001). Out of the examined infant stool samples, 4% were positive for Cronobacter spp. and C. sakazakii was identified in two diarrheic cases (6.3%) (Table 1). Nearly similar results were reported previously (Kandhai et al. 2010, Liu et al. 2013).
Cronobacter spp. have many virulence factors that are related to Cronobacter penetration and pathogenicity to the host cells. The outer membrane protein A (ompA) is a virulence factor in Cronobacter spp. responsible for fibronectin-binding of the bacteria to human brain microvascular endothelial cells, basolateral invasion, and interaction with enterocytes, because the bacterium cannot bind to intestinal cells when ompA is not expressed (Mohan Nair and Venkitanarayanan 2006, Nair et al. 2009, Kim et al. 2010, Emami et al. 2011). The expression of ompA is essential for the onset of meningitis and resistance to blood and serum killing factors (Mittal et al. 2009). The absence of ompA expression reduced the invasion by a percentage of 87% (Nair et al. 2009). The presence of zpx encoding gene in different bacteria is associated with their ability to cross the blood-brain barrier due to degradation of extracellular matrix protein such as intravenous collagen leading to destruction of endothelial cells causing leakage of blood components to the surrounding tissue (Mohan Nair and Venkitanarayanan 2007). Furthermore, it may be responsible solely or in combination with enterotoxins for the necrosis and extensive cellular destruction in neonates with necrotizing enterocolitis (Nazarowec-White and Farber 1997, Pagotto et al. 2003).
Molecular amplification of ompA gene revealed that 66.7% of Cronobacter isolates harbored ompA associated gene, while, only the four isolates of animal feces origin (33.3%) were positive for zpx associated gene. In accordance, the two genes were reported in Cronobacter isolates in several studies (Jaradat et al. 2009, Terragno et al. 2009, Cai et al. 2013). ERIC-PCR have been used previously for the genotyping of different pathogens, however, few studies have utilized ERIC-PCR for genotyping of C. sakazakii (Ye et al. 2008). ERIC-PCR was chosen for typing of the isolates due to its previously reported reproducibility, rapidity, reliability, and high discrimination index (Wilson et al. 2009).
Six profiles (E1 to E6) and high discriminatory index of ERIC-PCR (0.9524) were produced. In concordance, Ye et al. (2008) reported that 22 Cronobacter isolates produced 16 ERIC profiles with 0.933 discriminatory index. The production of six ERIC profiles and only one cluster indicated the genetic diversity of C. sakazakii of different sources. The obtained results strengthen the previously reported genetic diversity of Cronobacter isolates (Ye et al. 2008, Terragno et al. 2009, Miled-Bennour et al. 2010, Fakruddin et al. 2014). However, a study has reported genetic similarity between milk and patient isolates using molecular genotyping (van Acker et al. 2001). The results obtained indicated that ERIC-PCR can be used for discrimination and genotyping of C. sakazakii isolates of different sources. Moreover, the obtained isolates are heterogenic and genetically not related. Thus, further studies with a larger sampling size from different sources are required.
One of the C. sakazakii isolates of animal feces origin was selected for further investigation of its ability to survive and resist heating at different temperatures. The infection dose of Cronobacter spp. has been reported to be 103–105 CFU/mL, however, lower numbers are normally found in reconstituted infant powdered milk (Pagotto et al. 2003). Nonetheless, unhygienic practices during preparation and inappropriate storage temperature may enhance the growth and survival of such organism in infant powdered milk (Al-Holy et al. 2009). Reconstituted infant powdered milk is a nonsterile product that should be prepared with precautions. Consequently, heat treatment is essential before feeding infants to avoid infection with C. sakazakii (Al-Holy et al. 2009).
In this study, there was no significant change of C. sakazakii concentration in infant powdered milk at refrigeration temperature (4°C). Previous studies have demonstrated that C. sakazakii did not grow at 4°C (Nazarowec-White and Farber 1997, Richards et al. 2005, Ghassem et al. 2011). These findings support the significance of proper preservation of reconstituted infant powdered milk at refrigeration temperatures (4°C) for 24–48 h to prevent the growth of the micro-organism (Berseth et al. 2004, Al-Holy et al. 2009, Ghassem et al. 2011).
At ambient temperature, a significant increase was observed after 24, 48, 72, and 96 h (p ≤ 0.05). In accordance, previous reports confirmed that C. sakazakii was able to grow and multiply at room temperature (Pagotto et al. 2003, Ye et al. 2008, Wilson et al. 2009).
The results in Table 4 show that D-values of C. sakazakii were 13.79, 9.13, 7.51, 5.74, and 4.64 min at 54°C, 56°C, 58°C, 60°C, and 64°C, respectively, while, the z-value was 17.42°C. These results indicated that the examined C. sakazakii isolate of animal feces origin showed high heat resistance. Other studies have also reported the ability of Cronobacter isolates to resist heating (Iversen et al. 2004, Al-Holy et al. 2009, Fakruddin et al. 2014). In contrary, at 58°C the D-values of Cronobacter spp. ranged from 0.39 to 0.6 min indicating that these strains were not thermoresistant (Breeuwer et al. 2003).
Conclusion
The results of this study show that Cronobacter spp. are frequently found in infant powdered milk and stool of neonates. Furthermore, this study, to the best of our knowledge, is the first report of isolating C. sakazakii from buffalo feces indicating their role in environmental contamination. The isolate from animal feces has the ability to survive refrigeration and room temperature and shows high heat resistance. Public health education regarding sources of contamination with C. sakazakii is essential to decrease risk of infection.
Footnotes
Author Disclosure Statement
No competing financial interests exist.