
Abstract
The purpose of this study is to investigate the relationships between habitat structure and Babesia spp. occurrence in rodents in mainland Southeast Asia. Of 1439 rodents and insectivores investigated, the protist Babesia was found in only 81 individuals (5.6% of the micromammals investigated) with Babesia microti U.S. type the more prevalent (4.1%), followed by the B. microti Kobe type (1.2%), and by the very rare Babesia BiCM002 (0.04%). We used georeferenced data of rodents analyzed for Babesia infection and land cover maps produced for the seven study sites in Thailand, Cambodia, and Lao PDR, where they were collected. Rodents infected by Babesia were more likely to be found in settlements close to forested areas, which may represent risky place for spillover of Babesia species to humans.
Introduction
B
Human babesiosis is a neglected disease due to its low incidence, although its occurrence is underestimated in Asia. Babesiosis was reported in Japan (Tsuji et al. 2001, Wei et al. 2001, Zamoto-Niikura et al. 2012) with Babesia microti, Korea (Kim et al. 2007) with Babesia sp. KO1, Taiwan (Shih et al. 1997) with B. microti-like, China (Zhou et al. 2014, Fang et al. 2015, Jiang et al. 2016, Man et al. 2016) with B. microti-like, B. venatorum, and Babesia sp. XXB/HangZhou, and the area of China-Myanmar border (Zhou et al. 2013, 2015) with Babesia sp.
Rodents serve as reservoirs of several important pathogens of medical importance (Meerburg et al. 2009, Blasdell et al. 2015, Bordes et al. 2015a), including Babesia (Saito-Ito et al. 2004). B. microti and B. microti-like are the predominant babesial parasites found in rodent hosts and their associated ticks (Rar et al. 2016). However, the role of environmental factors in the disease transmission is still poorly investigated.
In Asia, Babesia in rodent hosts was documented in Japan by field and molecular surveys. B. microti is a genetically complex group found from field rodents, which is composed of three lineages as U.S. and Kobe and Hobetsu types (Tsuji et al. 2001, Saito-Ito et al. 2004) from four lineages, including B. microti-like (Zamoto et al. 2004, Saito-Ito et al. 2007), almost from Apodemus rodents and small mammals. B. microti in rodents was reported from investigation of captured rodents from Kinmen Island of Taiwan only in Rattus losea (Chao et al. 2017). There is no documentary report of these from Southeast Asia, except a publication from more than 10 years ago according to Babesia sp. BiCM002 from Bandicota indica rodents in Chiangmai province of Thailand (Dantrakool et al. 2004).
Southeast Asia, a hotspot of biodiversity at risk, is experiencing large-scale human modification of habitats (Sodhi et al. 2004, Wilcove et al. 2013). The ongoing conversion of natural habitats to either agricultural or urban land may lead to new risks of zoonotic diseases (Bordes et al. 2017). Disturbed landscape often benefits more generalist species, and particularly rodent reservoirs of zoonotic diseases, which are more tolerant and adaptable to ecological change, including conversion to agriculture (Morand et al. 2015).
The purpose of this study is to investigate the relationships between habitat structure and Babesia spp. occurrence in rodents in Southeast Asia. This study follows several published studies investigating rodent-borne diseases in the framework of the CERoPath project (Blasdell et al. 2015), such as hantavirus (Blasdell et al. 2012), Bartonella species (Jiyipong et al. 2015), or Trypanosoma species (Pumhom et al. 2015). For this, we used the land covers developed for Southeast Asian localities investigated in this study (Bordes et al. 2015b, Morand et al. 2015). Specifically, we hypothesized that heterogeneous habitats with high forest cover may favor the transmission ecology of Babesia among rodents and their ticks, and their potential transmission to humans.
Materials and Methods
Ethical statement
Rodent species included in the study are neither on the CITES list nor the Red List (IUCN). Animals were treated in accordance with the guidelines of the American Society of Mammalogists and within the European Union legislation guidelines (Directive 86/609/EEC). Each trapping campaign was validated by the national, regional, and local health authorities. Approval notices for trapping and investigation of rodents were provided by the Ethics Committee of Mahidol University, Bangkok, Thailand, Number 0517.1116/661.
Rodents
Rodents were trapped in the Thai provinces of Buriram (14.89 N; 103.01 E), Loei (17.39 N; 101.77 E) and Nan (19.15 N; 100.83 E), the Laotian provinces of Champasak (15.12 N; 105.80 E) and Luang Prabang (19.62 N; 102.05 E), and the Cambodian provinces of Mondolkiri (12.04 N; 106.68 E) and Preah Sihanouk (10.71 N; 103.86 E) in 2008 and 2009 within the framework of the CERoPath project (

Geographic locations of the seven trapping localities in mainland Southeast Asia (see Materials and Methods section). The trapping localities in three countries on mainland are composed of Nan, Loei and Buriram provinces in Thailand
Rodent species were identified in the field using morphological criteria, but were confirmed using molecular methods, if needed, using a mitochondrial gene for barcoding of some rodent species (Galan et al. 2012). Complete data on the animals used as reference specimens for the barcoding assignment are available on the “Barcoding Tool/RodentSEA” section of the CERoPath project website (
Environmental indices and land use
The methodology was described in Morand et al. (2015) and Bordes et al. (2015a, 2015b). For each locality, recent (years 2007–2008) high spatial resolution (2.5 meters in panchromatic mode and 10 meters in multispectral mode) SPOT 5 satellite images were acquired (© CNES 2009, distributed by Astrium Services/Spot Image S.A., France). SPOT-Digital Elevation Model (DEM) with a spatial resolution of 20 meters together with SRTM (Shuttle Radar Topography Mission) was also acquired. For each locality, the SPOT scene was classified into different land cover types using an object-based approach. The land cover maps and the DEM were integrated into a GIS to compute landscape metrics for each trapping site. To describe the landscape surrounding the trapping location of each individual rodent, landscape metrics were calculated within a 100-m radius. These metrics included the following: cover of agriculture on steep land (Agri steep), cover of agriculture on flat land (Agri flat), cover of forest (Forest), cover of human settlement (Settl), a proxy of habitat diversity (patch density) and habitat fragmentation (edge density), and all distances between each rodent trapped and each land cover type (agriculture on steep land, agriculture on flat land, forest, and human settlement).
Babesia
Babesia species were investigated in rodent blood samples by the nested PCR approach for partial sequence of 18 s rRNA gene with two couples of primers by the following steps: packed red cells were subjected for extraction of genomic DNA using DNA extraction kit (DNeasy® Blood & Tissue Kit; QIAGEN, Beckman Instrument, Inc.). The nested PCR reactions were performed as recommended protocols in LCBM-EA4558 research unit (Laboratoire de Biologie Cellulaire et Moleculaire; UFR Sciences Pharmaceutiques et Biologiques, Université de Montpellier, France). 100 ng/μL of genomic DNA was used for detection of the Babesia and their relative species by nested polymerase chain reaction (nested PCR) with two couples of the specific primer sets for piroplasm 18 s rRNA gene by (1) UpPIRO-A and DownPIRO-B (forward: 5′-GACGGTAGGGTATTGGCCTAC-3′ and reverse: 5′-GACGACCTCCAATCTCTAGTC-3′) and (2) PIRO-A and PIRO-B primer set as previously described by Omeda et al. (forward: 5′- AATTACCCAATCCTGACACAGGG-3′ and reverse: 5′-TTAAATACGAATGCCCCCAAC-3′) (Olmeda et al. 1997) with the specific thermal cycling profiles (denaturation 94°C for 45 s, 50°C for 45 s, and 72°C for 2 min; 35 cycles). The suspected band for Babesia and their relative species were targeted by 400–450 base pairs fragment on 1.5% agarose gels. The positive samples by nested PCR were the subjects for 18 s rRNA gene sequencing; amplified PCR fragments were cloned in a TOPO® TA Vector system (TOPO TA cloning kit for sequencing; Invitrogen, Thermo Fisher Scientific, Inc., MA) and transformed into E. coli-competent cells. Plasmid extraction was performed by using the plasmid DNA purification kit, NucleoSpin® Plasmid EasyPure (MACHEREY-NAGEL GmbH & Co. KG, Germany) featured for specially treated silica membrane binding The plasmid DNA was sent for Sanger DNA sequencing by outsource (GENEWIZ Europe, Takeley, United Kingdom). DNA sequenced 400–450 base pair fragments were analyzed for Babesia species and types by comparing with sequence databases based on the statistical significance by the Basic Local Alignment Search Tool; BLAST (National Center for Biotechnology Information; NCBI).
Statistical analysis
We modeled the probability of Babesia infection as a function of several environmental indices with a logistic regression (GLMM with logit function) using package lme4 implemented in R. Cases in each logistic regression were infected or noninfected individual rodents.
The initial model (Model 1) included the explanatory variables relating distance for each individual rodent trapped to the main habitats, shorter distance from forest (Dist Forest), shorter distance from human settlement (Dist Settl), shorter distance from agriculture on steep land (Dist Agri steep), and shorter distance from agriculture on flat land (Dist Agri flat), and the environmental indices related to the habitat structure, and calculated for a buffer of 100 meters for each individual rodent trapped: cover of agriculture on steep land (Agri steep), cover of agriculture on flat land (Agri flat), cover of forest (Forest), cover of human settlement (Settl), patch density, edge density, and SHDI. No interactions were added among the independent variables. We included locality and rodent species as random variables. We evaluated support for competing models investigating the relationship between Babesia occurrence and all explanatory variables of interest. We used likelihood-based methods (Burnham and Anderson, 2002) to quantify the alternative models and estimate their parameters. We used AIC adjusted for sample size (AICc) to assess the relative information content of the models. We quantified the uncertainty that the “best” model would emerge as superior if different data were used with Akaike weights wr (Burnham and Anderson, 2002). Selection of the best competing models was made using package gmulti version 1.0.7 (Calcagno and de Mazancourt, 2010) implemented in R, which allows the exploration of all models using automated model selection and model averaging procedure. Following this fist model, we modeled the most prevalent Babesia species (see Results section).
Results
Babesia
Babesia species infected in rodents and small mammals in this study was investigated in rodent blood samples by the nested PCR approach of partial sequence of 18 s rRNA gene with two couples of primer, and then the suspected fragments were analyzed by sequencing. This Babesia detection has shown 5.6% infections in 17 from 27 rodent species by two Babesia species with two types inside as B. microti by U.S. type and Kobe type and Babesia species BiCM002 in differently spreading of Babesia species in each country (unpublished data; in preparing).
Of 1439 rodents tested for this analysis concerning seven localities, the protist Babesia was found in only 81 individuals (representing 5.6% of the micromammals investigated) from the following rodents: six Bandicota indica (B. microti U.S. type, B. microti Kobe type, and Babesia sp. BiCM002), 11 B. savilei (B. microti U.S. type and Babesia sp. BiCM002), one Berylmys bowersi (B. microti U.S. type), two Mus caroli (B. microti U.S. type and B. microti Kobe type), two M. cervicolor (B. microti U.S. type), five M. cookii (B. microti U.S. type and B. microti Kobe type), seven Maxomys surifer (B. microti U.S. type), two Niviventer fulvescens (B. microti U.S. type), two Rattus andamanensis (B. microti U.S. type and B. microti Kobe type), four Rattus argentiventer (B. microti U.S. type), 14 Rattus exulans (B. microti U.S. type), one Rattus nitidus (B. microti U.S. type), three Rattus norvegicus (B. microti U.S. type), six Rattus sakaratensis (B. microti U.S. type), and 13 Rattus tanezumi (B. microti U.S. type and B. microti Kobe type), one gymnure Hylomys suillus (B. microti U.S. type), and one insectivore Suncus murinus (B. microti U.S. type) (Table 1).
The parasite was found in all localities: Preah Sihanouk (B. microti U.S. type), Mondolkiri (B. microti U.S. type), Champasak (B. microti U.S. type), Luang Prabang (B. microti U.S. type and B. microti Kobe type), Buriram (B. microti U.S. type and Babesia sp. BiCM002), Loei (B. microti Kobe type), Nan (B. microti U.S. type and B. microti Kobe type) (Table 1).
Habitat characteristics and Babesia infection
We performed the statistical model on only 1254 individuals, for which we have enough accurate localization, for total Babesia infections (68 individuals). The top models among all potential candidate models that fit the entire dataset with variables related to habitat structure and distance to each habitat using logit function are presented in Table 2.
Models are ranked from lowest to highest supported according to corrected Akaike information criteria (AIC). Only localities with at least one individual infected rodent were kept for each analyzed dataset. The initial model included the following explanatory variables: cover of agriculture on steep land (Agri steep), cover of agriculture on flat land (Agri Flat), cover of forest (Forest), cover of human settlement (Settl), patch density (density), slope, distance to agriculture on flat land (DistFlat), distance to agriculture on steep land (DistSteep), distance to forest (DistForest), distance to human settlement (DistSettl), and rodent weight (K is the number of estimated parameters, AICc the selection criterion, and wr the Akaike weights).
Results of GLM model-averaged importance of surrounding habitat characteristics explaining the infection of rodents by all Babesia type showed that only the explanatory variables rodent weight, distance to forest, and distance to settlement were found in 100% of the top best models (Fig. 2A). Hence, the first best top model selected using AICc values demonstrated that large rodents (p = 0.024) are likely infected when they lived close to human settlement (p = 0.062) and close to forest (p = 0.06) (Tables 2 and 3). However, rodent weight was the only significant variable at the level of 5% (Table 3).
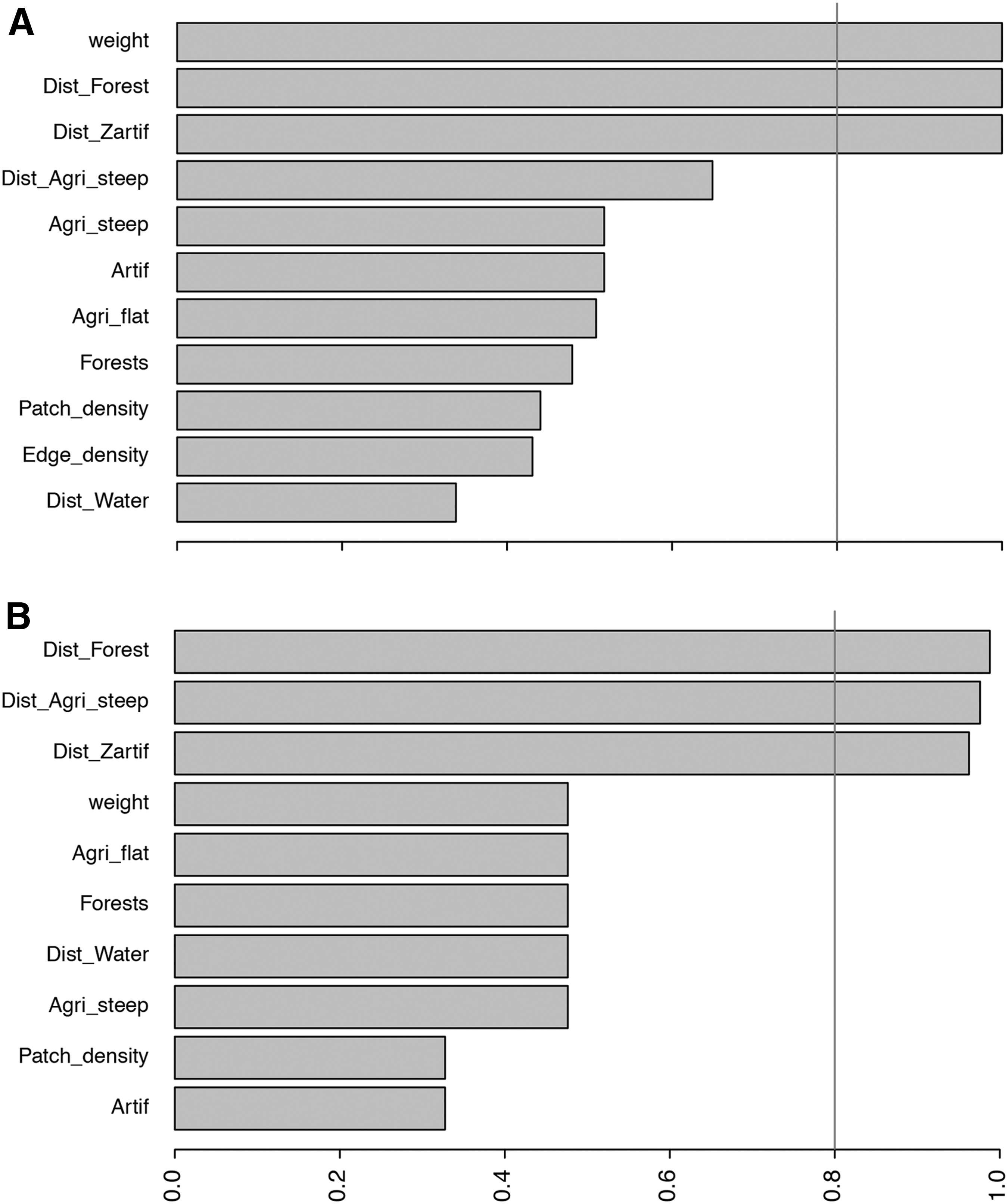
Results of GLMM (using logit function, and with locality and rodent species as random factors) model-averaged importance of surrounding habitat characteristics explaining the infection of rodents by
Investigating B. microti U.S. type using the same modeling approach, performed on 47 infected rodents for a total of 1264 individuals, showed that the distance to forest, distance to agriculture on steep land, and distance to settlement were found in more than 90% of the top best models (Fig. 2A). The first best top model selected using AICc values demonstrated that rodents are likely infected when they lived far to agriculture on steep land (p = 0.088) and close to forest (p = 0.11) (Tables 2 and 3). However, none of these variables was significant at the level of 5% (Table 3).
Discussion
The goal of this study was to investigate the relationship between the landscape structure of several sites in Southeast Asia and the prevalence of Babesia spp. in rodents and some insectivores. Our results showed that almost all micromammals can be infected by several types of Babesia, in all types of habitat (forest, agriculture, and settlement) and all localities (Cambodia, Lao PDR, and Thailand) investigated in this study. However, our results also showed the very low prevalence of Babesia in these micromammals (5.6% in overall), with the U.S. type more prevalent (4.1%), followed by the Kobe type (1.2%), and by the very rare BICM002 (0.04%). Our study shows a very low prevalence of Babesia in rodent reservoirs in Thailand in comparison to the precedent study of Dantrakool et al. (2004), in which a prevalence of 36% of infection was observed, although the species diversity and numbers of investigated rodents were very low (30 B. indica and 17 R. exulans). We also confirmed the presence of BICM002 in Thailand (Dantrakool et al. 2004).
Several studies revealed that the prevalence of Babesia in small mammals may vary considerably according to the geographical region investigated. In Europe, the prevalence of B. microti in Croatia was observed between 6.6% and 14.0% (Beck et al. 2011, Tadin et al. 2012), but only 1.9% in Slovakia (Blaňarová et al. 2016). Babesia microti Munich type was found with a low prevalence of 0.4% in France (Jouglin et al. 2017), but with a higher prevalence of 40% in Finland (Kallio et al. 2014). In USA, only 1% of woodrats were found infected by B. microti in Texas (Charles et al. 2012), while the prevalence reached 35% in rodents in Florida (Clark et al. 2012). In Japan, the prevalence of B. microti Kobe type, B. microti Hobetsu types, U.S. type, and B. microti like varied from 5.4% to 50% (Tsuji et al. 2001, Okabayashi et al. 2002, Saito-Ito et al. 2004, Zamoto et al. 2004, Tabara et al. 2007). Similarly, in China, the prevalence of B. microti Kobe and Otsu (Hobetsu) lineages was observed ranging from 4.3% to 41.9% (Saito-Ito et al. 2008, Chen et al. 2017). In Taiwan, the prevalence of B. microti Kobe and Otsu (Hobetsu) lineages was observed ranging from 5.1% to 9.9% (Saito-Ito et al. 2008, Chao et al. 2017). The prevalence observed in our study is not exceptionally low, but remains difficult to explain without further investigation of the environmental factors, such as temperature and rainfall, which may play a significant role in the population dynamics of ticks and Babesia transmission.
Such low prevalence of infection was already observed for other bacteria and protists transmitted by arthropods in Southeast Asia (reviewed in Herbreteau et al. 2012), and in our studies investigating Bartonella species (Jiyipong et al. 2012, 2015) or Trypanosoma species (Pumhom et al. 2015) among the same rodent species in similar Southeast Asian environments.
This observed prevalence allowed us to explore the relationship between the parasite occurrence in the micromammals and the structure of landscape only for the whole Babesia type and for the U.S. type. Rodents living close to settlement and close to forest were likely infected by a Babesia parasite.
Our results showed that large body-sized rodents were more likely to be infected than small body sized, controlling for species identity. Infection is likely to increase with long-living rodents and with large home range, which are often correlated with body size (Morand and Bordes, 2015). Moreover, large body size animals offer a large target for ectoparasites (Morand, 2015), favoring the transmission of parasites by ticks.
Rodents living close to forest and far from agriculture land were more likely infected by B. microti U.S. type, although these variables were not significant at the risk of 5%. The low support values (weight w) can be explained by this low number of infected individuals.
There are other limitations of our study. We did not investigate the presence of Babesia in ticks. However, in our field works, ticks are scarcely observed. All these ticks, mostly larvae and nymphs, have been collected for further studies (identification and pathogen screenings). The trapping method and the behavior of ticks can explain this low observation. As the period and season for each field trip were different, we could not investigate how seasonality and climate factors might affect the dynamics of ticks, rodents, and their Babesia parasites.
Our study is the first to explore, to our knowledge, the diversity of rodent reservoirs of Babesia in relationship to habitat in mainland Southeast Asia. Our study has some implications for public health as it highlights that particular habitats, such as settlements close to forested areas, are likely place for spillover of Babesia species to humans.
Footnotes
Acknowledgments
This study was funded by the French ANR Biodiversity, grant ANR 07 BDIV 012 CERoPath project (“Community Ecology of Rodents and their Pathogens in a changing environment”) (
Author Disclosure Statement
No competing financial interests exist.