
Abstract
Introduction:
Leptospirosis remains a major public health threat in Cambodia. In this study, we aimed at facilitating the development of preventive strategies against leptospirosis in Cambodia by conducting molecular epidemiological surveys of Leptospira infection among wild rodents.
Materials and Methods:
One hundred sixty-three wild rodents were captured in the capital Phnom Penh and surrounding areas, and their kidneys and bladders were collected for analysis. Identification of wild rodent species was determined by using the cytochrome c oxidase I gene. TaqMan PCR of the flagellin B gene (flaB) was performed to detect Leptospira, and species of the isolates were identified by flaB sequencing analysis.
Results:
The species and respective number of rodents collected were as follows: Rattus norvegicus, 80 (49.1%); Rattus argentiventer, 53 (32.5%); Rattus exulans, 6 (3.7%); Rattus indica, 15 (9.2%); Maxomys surifer, 3 (1.8%); and Rattus sp., 6 (3.7%). Leptospira was detected in 20 out of the 163 rodents (12.3%) and was categorized as either Leptospira interrogans or Leptospira noguchii. R. norvegicus had the highest prevalence of Leptospira (17.5%), and R. argentiventer and Rattus sp. showed infection rates of 9.4% and 16.7%, respectively.
Conclusions:
Based on these results, wild rodents living in urban areas of Cambodia were found to be important reservoirs of highly virulent Leptospira. The flaB sequence results of our study provided information regarding the prevalence of Leptospira species, which was dependent on the rodent species. This study is the first study on leptospirosis in wild rodents in the urban areas of Cambodia, where there is limited information on leptospirosis.
Introduction
L
Rodents are the most important mammalian reservoirs of Leptospira (Adler and de la Pena Moctezuma, 2010). They maintain Leptospira in their kidneys and shed the organisms in their urine, which may contaminate water and soil in the environment. Infection of humans and other animals with Leptospira occurs mainly through exposure of skin and mucous membranes to contaminated water and soil or direct contact with the urine of carrier animals (Adler and de la Pena Moctezuma, 2010).
Epidemiological surveys are necessary to elucidate the distributions of Leptospira in certain geographic locations. Determination of the prevalence of Leptospira and identification of infecting serovars among reservoir animals are important for establishing prevention strategies against leptospirosis. However, no reports have described the infection rates of Leptospira among wild rodents in Phnom Penh, the capital city of Cambodia, and surrounding areas, despite the fact that Phnom Penh city is the most populous area in Cambodia (National Institute of Statistics 2008). Accordingly, in this study, we aimed at elucidating the prevalence of Leptospira infection, determining the infecting Leptospira species in wild rodents, and identifying the reservoir species of Leptospira in Phnom Penh to contribute to the development of preventive health strategies against Leptospirosis.
Materials and Methods
Collection of wild rodents and organ samples
Trapping of wild rodents was conducted from September 2013 to July 2014 in Phnom Penh and surrounding areas (Fig. 1). Wire-mesh traps with bait were left overnight at the market or public facilities in Phnom Penh city and in rice fields and households in Kandal and Kampong Speu province. Captured wild rodents were euthanized by using halothane as anesthesia. Kidneys and bladders were aseptically collected for DNA extraction.
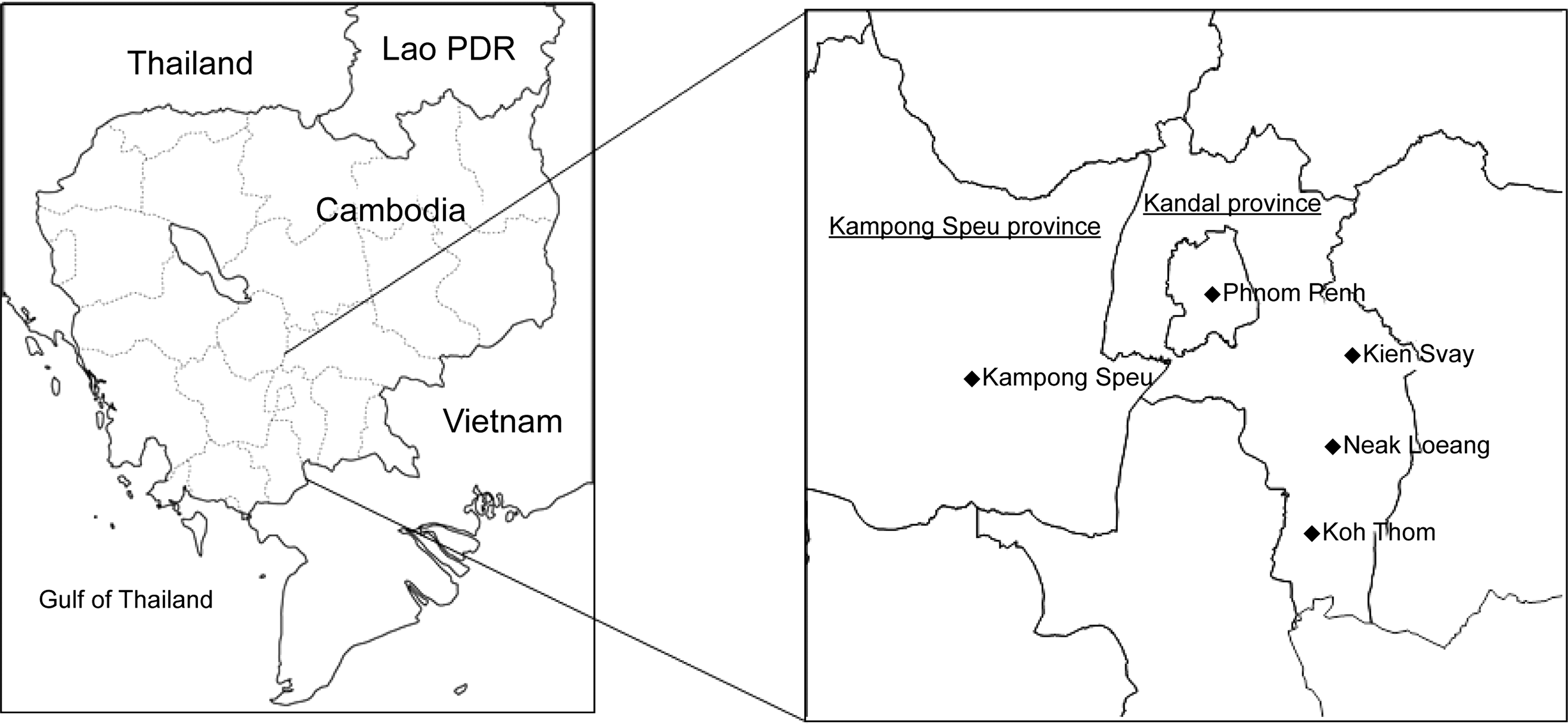
Sampling sites (⧫) in Cambodia.
DNA extraction from wild rodent tissues
DNA was extracted from the kidneys and bladder, using a QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol.
Identification of species of wild rodents by cytochrome c oxidase gene analysis
Cytochrome c oxidase I (COI) gene sequences of mitochondrial DNA were used to identify the species of trapped wild rodents according to the method described in Community Ecology of Rodents and their Pathogens in South-East Asia (CERoPath,
TaqMan PCR targeting Leptospira flaB
TaqMan PCR assays targeting the flagellin B gene (flaB) were performed to determine the presence of Leptospira DNA in the kidneys and urinary bladders of wild rodents. The TaqMan PCR assay was performed with a 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA) by using the following PCR mixture: Premix Ex Taq (Perfect Real Time; Takara, Otsu, Japan), forward primer L-flaB537 (5′-AGATGCTGCCCTTACGAAGA-3′), reverse primer L-flaB624R (5′-CATCAGACCCTTTGCGGTAT-3′), TaqMan probe L-flaB561F (5′-FAM-GAAGCAGAGAGCGGATATGG-MGB-3′), which harbored the fluorescent reporter dye 6-crboxy-fluorescein (FAM) at the 5′ end of the probe and the minor groove binder (MGB) at the 3′ end, and ROX dye II. Cycling conditions were as follows: 1 min at 95°C, followed by 42 cycles of 10 s at 95°C and 40 s at 60°C. Results were compared with the readings of positive control DNA, which was extracted from Leptospira interrogans serovar Losbanos strain LT 101-69 and reagent blank as a negative control.
Nested PCR targeting Leptospira flaB and sequence analysis
To confirm Leptospira infection among the trapped rodents, nested PCR targeting flaB and sequencing analyses were performed on TaqMan-PCR positive DNA. flaB was amplified by using the forward primer L-flab-F1 (5′-CTCACCGTTCTCTAAAGTTCAAC-3′) and reverse primer L-flab-R1 (5′-TGAATTCGGTTTCATATTTGCC-3′) under the following conditions: an initial cycle of 10 min at 94°C; 40 cycles of 94°C for 30 s, 60°C for 30 s, and 72°C for 1 min; and 10 min at 72°C amplifying a 790-base pair (bp) DNA fragment. For the nested PCR, 1 μL of the first PCR product was added to a 19-μL second round PCR mixture. Amplification was performed by using the primers L-flab-F2 (5′-TGTGCACAAGACGATGAAAGC-3′) and L-flab-R2 (5′-AACATTGCCGTACCACTCTG-3′) under the following conditions: an initial cycle of 10 min at 94°C and 40 cycles of 94°C for 30 s, 58°C for 30 s, and 72°C for 1 min, amplifying a 734-bp DNA fragment. A final extension was performed for 10 min at 72°C. PCR amplification was confirmed by electrophoresis on 1.5% agarose gels. The nested PCR amplicons purified with Microcon-PCR purification columns (Millipore, Bedford, MA) were subjected to DNA cycle-sequencing analysis, using nested PCR primers as sequencing primers, and a BigDye Terminator v3.1 cycle sequencing kit (Applied Biosystems) with an ABI 3130-Avant Genetic Analyzer (Applied Biosystems). Phylogenetic analysis was performed by the Clustal W algorithm by using sequence analysis software (MegAlign; DNASTAR, Inc., Madison, WI). A phylogenetic tree was constructed by the neighbor-joining method with 1000 bootstrap resampling.
The resulting DNA sequence data were deposited in and assigned to the DNA Data Bank of Japan (DDBJ) and confirmed with the BLAST algorithm (
Animal ethics
Animal experiments were carried out in accordance with the U.K. Animals Scientific Procedures. This study was reviewed and approved by the Ethic Committee of the Ministry of Health, Royal Government of Cambodia.
Statistics analysis
Statistical analysis was performed by chi-squared tests with SPSS ver. 20J (IBM; Armonk, NY). Differences with p values of less than 0.05 were considered significant.
Results
Identification of rodent species
A total of 163 wild rodents were trapped in the study sites. The COI gene in rodent DNA kidney was amplified, and amplicons from 41 rodents were initially sequenced and phylogenetically analyzed (Fig. 2).

Phylogenetic analysis of wild rodents based on the mitochondrial COI gene. The phylogenetic tree was constructed by aligning COI sequences by using the Clustal W algorithm, followed by the neighbor-joining method with 1000 bootstrap resampling. The sequences obtained in this study are indicated as serial numbers. Sequence accession numbers are indicated in parentheses. COI, cytochrome c oxidase I.
The first 41 samples were applied to COI sequences to establish RFLP patterns for the identification of rodent species. By this analysis, 122 rodent species were identified.
From the COI sequences obtained in this study, 28 out of 41 were clustered under Rattus argentiventer (HM217484), 10 were Rattus norvegicus (HM217501), and two were Rattus exulans (HM217508). The sequence derived from rodent no. 15 was similar to that of an unknown Rattus species (HM217485). The remaining 122 amplicons were subjected to RFLP analysis of COI amplicons to identify the rodent species (Table 1).
Positive rate of details in same item was compared by chi-squared test.
COI amplicons (∼650 bp) were digested with MboI, and 2–6 fragments were generated. RFLP of these amplicons was compared with those of different rodent species, such as R. norvegicus, R. argentiventer, and R. exulans. These RFLPs were consistent with the expected fragment patterns based on COI sequences. From the results for the 163 rodents, the following species were identified: R. norvegicus, 80 (49.1% of the total number of rodents); R. argentiventer, 53 (32.5%); R. exulans, 6 (3.7%); Bandicota indica, 15 (9.2%); Maxomys surifer, 3 (1.8%); and Rattus sp., 6 (3.7%). R. norvegicus was found in all sites. However, R. exulans was trapped only in Kampong Speu province, and M. surifer was trapped only in the Neak Loeang district, Kandal province.
Prevalence of Leptospira in rodents
Kidneys (n = 139) and/or bladders (n = 163) of wild rodents were collected and examined for Leptospira infection. Thirteen of 139 kidneys (9.4%) and 8 of 163 bladders (4.9%) of rodents were positive for Leptospira, using TaqMan PCR targeting flaB (Table 2). One rodent (no. 11) was Leptospira positive in both kidneys and bladder.
Twenty out of 163 rodents (12.3%) were found to be infected with Leptospira. R. norvegicus had the highest prevalence of Leptospira (17.5%; Table 2).
R. argentiventer and Rattus sp. were also found to be infected (9.4% and 16.7%, respectively). However, Leptospira was not found in the other species of rodents. In terms of sex, Leptospira positivity tended to be higher in male rodents than in female rodents (19% and 8.6%, respectively; p < 0.079). The results also showed that there were no statistically significant differences in the distribution, rodent species, or season when the rodents were captured.
Identification of Leptospira species
For identification of the Leptospira species infecting the trapped rodents, flaB sequences from 21 positive samples on TaqMan PCR were phylogenetically analyzed. Twenty-one samples were categorized into two groups: L. interrogans and Leptospira noguchii. In this study, these species were identified in R. norvegicus and R. argentiventer. Leptospira (B66 and B58) detected from R. argentiventer captured in Koh Thom district in Kandoal province were classified as L. noguchii. The remaining sequences were classified as L. interrogans. These were in R. norvegicus, R. argentiventer, and Rattus sp. (Fig. 3).
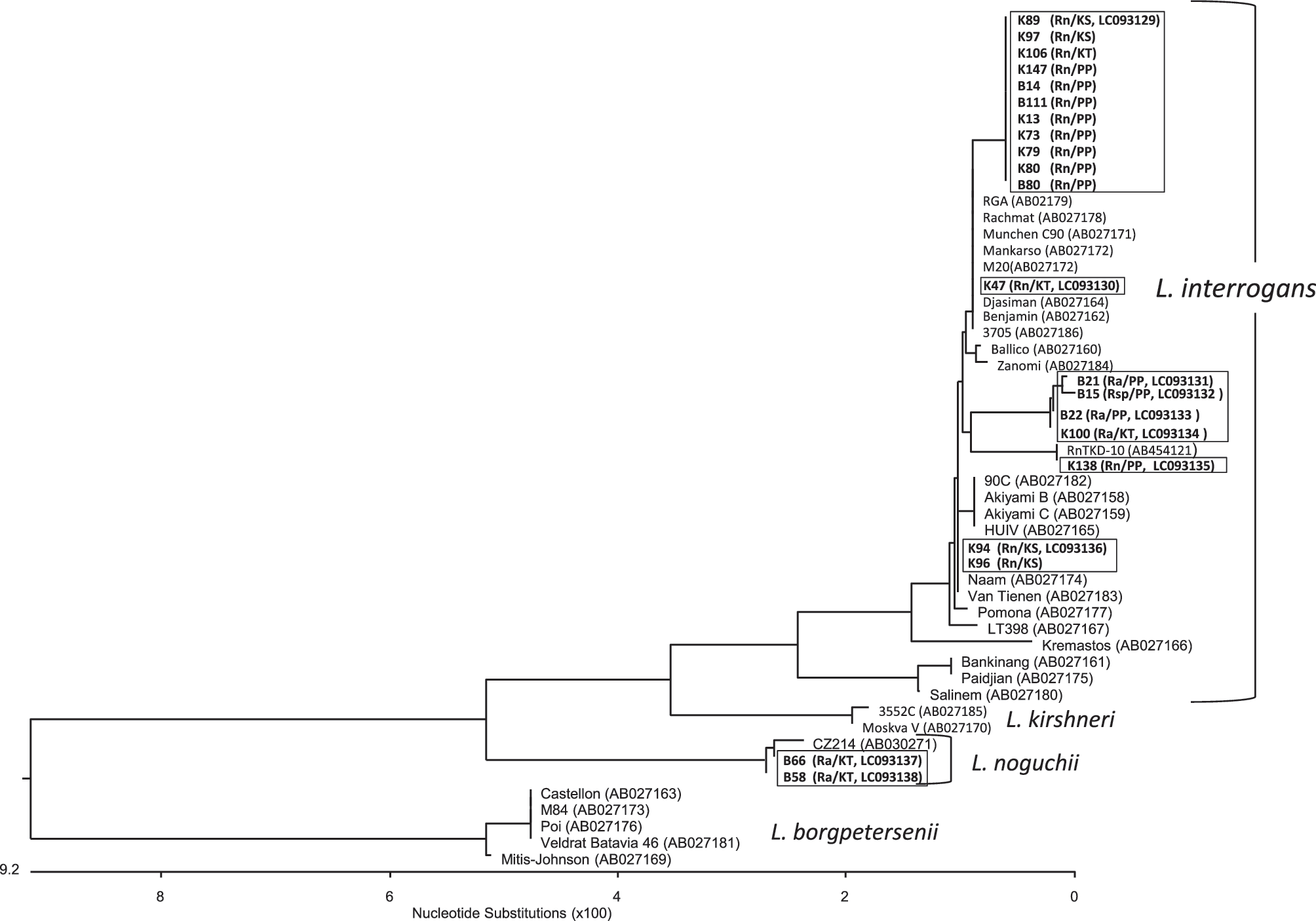
Phylogenetic tree based on flaB sequences of isolated Leptospira. The phylogenetic tree was constructed based on the alignment of flaB sequences using the Clustal W algorithm, followed by the neighbor-joining method with 1000 bootstrap resampling. The sequences from this study are highlighted in bold. Wild rodent species, trapping sites, and sequence accession numbers are indicated in parentheses. Rn, Rattus norvegicus; Ra, Rattus argentiventer; Rsp, unknown Rattus species; KS, Kien Svay; KT, Koh Thom; PP, Phnom Penh.
Discussion
In this study, we aimed at elucidating the status of Leptospira infection in wild rodents in the urban areas of Cambodia. Cambodia is located in the Indochina peninsula in southeast Asia, which experiences heavy rains and flooding.
Our results indicated that there was a high incidence of Leptospira infection in wild rodents, as determined by TaqMan PCR targeting flaB (12.3%). TaqMan PCR targeting flaB is more sensitive and effective for identification of Leptospira infection than the method using lipL32 or 16srRNA (Kawabata et al. 2001). The cause of positive results in the bladder sample, which might contain urine, and of negative results in the kidney sample in the same rat might relate to the selection of part of kidney tissue.
The prevalence rate was higher than those in Vietnam (5.8%) by real-time PCR targeting 16S rDNA (Loan et al. 2015), higher than those in Luang Probang (7.9%) and the Champasak district (1.4%) in Lao PDR by PCR targeting the LipL32 gene, and similar to those from the Loei district (17.8%) and Nan district (2.1%) in Thailand by PCR targeting the LipL32 gene (Cosson et al. 2014).
In Cambodia, Ivanova et al. (2012) previously reported that 11.1% of wild rodents were infected with Leptospira, on the basis of nested PCR targeting 16S rDNA. The prevalence of leptospirosis in the Mondulkiri district, which is a mountainous area, was 16.9%; whereas that in the Preah Sihanouk province, which is on the gulf coast, was 8.1% (Ivanova et al. 2012). Our study revealed a high Leptospira carriage rate in wild rodents in the urban areas of Cambodia.
Results of flaB sequence analysis showed that 19 out of the 21 sequences were L. interrogans, and the remaining two were L. noguchii. In a previous study of Leptospira infection among humans in Cambodia, L. interrogans was the most common species detected among febrile patients in the rural areas of Cambodia. Leptospira weilii was the next most common species, and L. noguchii was found to infect humans (Mueller et al. 2014). In another study, L. interrogans and Leptospira borgpetersenii were detected among wild rodents in rural areas of Cambodia (Cosson et al. 2014). Our results were similar to those of previous reports and indicated that L. interrogans was widely distributed among wild rodents. Further, wild rodents were found to be reservoirs of pathogenic Leptospira, which may contribute to human leptospirosis in Cambodia. Information about rodents (i.e., species of wild rodents, trapping site, and environmental characteristics) is critical for establishing preventive programs against leptospirosis. In our study, we aimed at elucidating the species of rodents that are possible reservoirs of Leptospira. For identification of rodent species, RFLP analysis of the COI gene was introduced in our study instead of morphological criteria. R. norvegicus and R. argentiventer were the most common rodent species trapped in this study, and both were found to harbor pathogenic Leptospira. Three other rodent species, that is, R. exulans, B. indica, and M. surifer, were also captured in this study. However, none of these species was infected with Leptospira.
In the study by Ivanova et al. (2012), a high prevalence (7/12 [58.3%] during the dry season and 7/36 [19.4%] during the rainy season) was observed in R. argentiventer, which was trapped only in the Preah Sihanouk province. R. exulans was also found in both areas, but it was not infected (Ivanova et al. 2012). These results were similar to those of our study; however, there was a discrepancy in the results of Leptospira infection of R. norvegicus. In the study by Ivanova et al. (2012), no R. norvegicus were infected in the dry or rainy seasons. In our study, R. norvegicus showed a high prevalence (17.5%), consistent with previous studies in Vietnam (18.3%), Thailand (5.6%), Malaysia (6.6%), and other countries in southeast Asia (Kositanont et al. 2003, Benacer et al. 2013, Loan et al. 2015). The results of our study suggested that R. norvegicus and R. argentiventer are potential reservoirs of L. interrogans in urban areas of Cambodia (Table 2). However, there is a limitation to identify all infection routes from animal hosts to humans in this study, and it would be recommended to examine other reservoir hosts, dogs, pigs, and cattle, in future studies.
Conclusions
In conclusion, wild rodents living in urban areas of Cambodia were found to be important reservoirs of highly virulent Leptospira. Leptospira are transmitted by rodent urine, which may contaminate water and soil in the environment. In addition, the flaB sequence results of our study provided information regarding the prevalence of Leptospira species and demonstrated that this prevalence depended on the rodent species. This study is the first study on leptospirosis in wild rodents in the urban areas of Cambodia, where limited information on leptospirosis is available. However, our study was limited by the sampling timings and small number of samples in each area. In future studies, we aim at evaluating larger numbers of samples and more diverse areas to obtain more information on the epidemiology of this disease.
Footnotes
Acknowledgment
This study was supported by a CIS Grant for Research and Education in 2013 and 2014.
Author Disclosure Statement
No competing financial interests exist.