
Abstract
Human lice, Pediculus humanus, are obligate blood-sucking parasites. Phylogenetically, they belong to several mitochondrial clades exhibiting some geographic differences. Currently, the body louse is the only recognized disease vector, with the head louse being proposed as an additional vector. In this article, we study the genetic diversity of head and body lice collected from Bobigny, a town located close to Paris (France), and look for louse-borne pathogens. By amplifying and sequencing the cytb gene, we confirmed the presence of clades A and B in France. Besides, by amplifying and sequencing both cytb and cox1 gene, we reported, for the first time, the presence of clade E, which has thus far only been found in lice from West Africa. DNA from Bartonella quintana was detected in 16.7% of body lice from homeless individuals, but in none of the head lice collected from 47 families. Acinetobacter DNA was detected in 11.5% of head lice belonging to all three clades and 29.1% of body lice. Six species of Acinetobacter were identified, including two potential new ones. Acinetobacter baumannii was the most prevalent, followed by Candidatus Acinetobacter Bobigny-1, Acinetobacter calcoaceticus, Acinetobacter nosocomialis, Acinetobacter junii, and Candidatus Acinetobacter Bobigny-2. Body lice were found to be infected only with A. baumannii. These findings show for the first time, the presence of clade E head lice in France. This study is also the first to report the presence of DNAs of several species of Acinetobacter in human head lice in France.
Introduction
S
Mitochondrial genes (cytb and COI) appear to separate lice into five divergent mitochondrial clades (A, B, C, D, and E) exhibiting some geographic differences (Ashfaq et al. 2015, Drali et al. 2015, Amanzougaghene et al. 2016b). Head lice encompass all clades, while body lice belong only to clades A and D (Ashfaq et al. 2015, Drali et al. 2015, Amanzougaghene et al. 2016b). The clade A is the most common and is found around the world, while clade D is only found in the Republic of Congo and Congo Brazaville (Light et al. 2008a, Ashfaq et al. 2015, Drali et al. 2015, Amanzougaghene et al. 2016a). Clade B is found in America, Europe, Australia, and North and South Africa, and was most recently reported in Israel, on head lice remains dated ∼2000 years old (Light et al. 2008a, Amanzougaghene et al. 2016b). Clade C has been found in Ethiopia, the Republic of Congo, and in Asia (Amanzougaghene et al. 2016a). Last, clade E consists of head lice from West Africa (Senegal and Mali) (Amanzougaghene et al. 2016b).
Body lice are potentially more harmful than head lice, because of their role as a vector of at least three pathogenic bacteria that have killed millions of people: Rickettsia prowazekii (the causative agent of epidemic typhus), Bartonella quintana (trench fever), and Borrelia recurrentis (relapsing fever) (Raoult and Roux 1999, Veracx and Raoult 2012). Natural and experimental observations have been made that body lice can also transmit Yersinia pestis, the causative agent of plague, and that they may be the pandemic vectors of this agent (Blanc and Baltazard 1942, Houhamdi et al. 2006). Some other widespread pathogenic bacteria, such as Serratia marcescens, Acinetobacter baumannii, and Acinetobacter lwoffii, have been detected in human body lice, with the assumption that lice may probably also transmit these agents to humans (La Scola et al. 2001, Houhamdi and Raoult 2006).
Although body lice are currently assumed to be more potent vectors of pathogens, the vector potential of head lice is not yet fully understood. Studies have demonstrated that the immune reactions of head lice to different pathogens are stronger than those of body lice, which obviously may carry a broad spectrum of pathogens (Previte et al. 2014, Kim et al. 2017). In laboratory-reared lice, it has been demonstrated that head lice can support a persistent load of B. quintana for several days following acquisition in a bloodmeal (Previte et al. 2014). An experimental infection with R. prowazekii has also shown that head lice can be readily infected and disseminate these pathogens in their feces, showing that these lice might be a vector of pathogens under optimal epidemiologic conditions (Robinson et al. 2003). Indeed, a substantial number of studies have reported body louse-borne pathogens on head lice collected from different parts of the world. This is the case of B. quintana, B. recurrentis, and Y. pestis DNA, found in head lice belonging to different mitochondrial clades (Angelakis et al. 2011a, 2011b, Boutellis et al. 2012, 2013, Drali et al. 2015, Amanzougaghene et al. 2016a). Several Acinetobacter species have also been detected in human head lice (Sunantaraporn et al. 2015, Amanzougaghene et al. 2016a).
In this study, we examined the genetic diversity of head and body lice collected from Bobigny, a town located 3 km north of Paris (France) (48°54′38″N 2°26′23″E), to look for louse-borne pathogens in these lice.
Materials and Methods
Ethical clearance
The protocol was reviewed and approved by the Comité de Protection des Personnes (institutional review board) of the CPP-Ile-de-France X (2017-02) Ethics Committee. Informed consent was obtained from all patients.
Study area and lice sampling
Between September 2015 and December 2016, head lice were collected from patients attending the Avicenne Hospital in Bobigny. One hundred forty-one patients belonging to 47 families were enrolled in this study, and 5 head lice were randomly selected per parasitized family (Supplementary Table S1; Supplementary Data are available online at
DNA extraction
Before DNA isolation and to avoid external contamination, the surface of each louse was decontaminated as described previously (La Scola et al. 2001). DNA was extracted using the QIAamp DNA tissue extraction kit (Qiagen, Hilden, Germany) in an EZ1 apparatus following the manufacturer's instructions.
Genotypic status of lice
Identification of louse mitochondrial clade by qPCR assays
To identify the mitochondrial clades of the lice, all DNA samples were analyzed using clade-specific quantitative real-time PCR (qPCR) assays that targeted a portion of cytochrome b (cytb) gene specific to clades A, D, B, and C, as previously described (Amanzougaghene et al. 2016a). It is important to note that, when the design of the qPCR specific to clade C was performed, clade E was classified as subclade within clade C; therefore, this qPCR detects both clades C and E (Amanzougaghene et al. 2016a). To discriminate between them, we performed another qPCR essay, specific only to clade E (Amanzougaghene et al. 2017). We used lice with known clades as positive controls and master mixtures as a negative control for each test.
PCR amplifications and sequencing
For phylogenetic study, 79 (79/235) head lice and 5 (5/24) body lice were randomly selected and subjected to standard PCR, targeting a 347-bp fragment of the cytb gene as previously described (Li et al. 2010). To confirm the presence of clade E, sixteen lice already identified as clade E by the previous PCR were subjected to another standard PCR, targeting another mitochondrial gene, cytochrome oxidase subunit 1 (cox1), as previously described (Drali et al. 2016).
PCR amplification was performed in a Peltier PTC-200 model thermal cycler (MJ Research, Inc., Watertown, USA). The reactions were carried out using the Hotstar Taq polymerase (Qiagen), in accordance with the manufacturer's instructions. Purification of PCR products was performed using NucleoFast 96 PCR plates (Macherey-Nagel EURL, Hoerdt, France) as per the manufacturer's instructions. The amplicons were sequenced using the Big Dye Terminator Cycle Sequencing Kit (Perkin Elmer Applied Biosystems, Foster City, CA) with an ABI automated sequencer (Applied Biosystems). The electropherograms were assembled and edited using ChromasPro software (ChromasPro 1.7; Technelysium Pty Ltd., Tewantin, Australia).
Molecular screening for the presence of pathogen DNA
The qPCRs were performed to screen all lice samples, using previously reported primers and probes, for Acinetobacter spp., Borrelia spp., B. quintana, Acinetobacter spp., R. prowazekii, Y. pestis, Coxiella burnetii, and Anaplasma spp. (Mediannikov et al. 2010, Nguyen-Hieu et al. 2010, Angelakis et al. 2011a, Bouvresse et al. 2011, Parola et al. 2011, Dahmani et al. 2017). All B. quintana-positive samples were confirmed by a second specific qPCR targeting the fabF3 gene (Angelakis et al. 2011a). All sequences of primers and probes used for qPCRs and conventional PCRs in this study are shown in Table 1.
All qPCRs were performed using a CFX96 Real-Time system (Bio-Rad, Marnes-la-Coquette, France) and the Eurogentec Master Mix Probe PCR kit (Eurogentec, Liège, Belgium). We included the DNA of the target bacteria as positive controls and master mixtures as negative control for each test.
To identify the species of Acinetobacter spp., all positive samples from qPCRs were subjected to standard PCR, targeting a portion of the rpoB gene as described previously (La Scola et al. 2006). Amplicons were prepared and sequenced using similar methods as described for the cytb gene for lice above.
Data analysis
For the head and body lice cytb and -cox1 sequences, unique haplotypes were defined using DnaSPv5.10 and compared with the cytb and cox1 haplotypes as described previously (Amanzougaghene et al. 2016a, Drali et al. 2016). All sequences of Acinetobacter species were analyzed using BLAST (
Results
Lice clade and phylogenetic analysis
Of the 141 patients (47 families) infested by head lice, the majority were female (sex ratio M/F = 0.1) aged between 2 and 53 years. No body lice were found.
In total, 235 head lice collected from the 47 families and 24 body lice collected from 2 homeless persons were analyzed using qPCRs to determine their clade. The result showed that 82 lice (31.8%, [82/258]) belonged to clade A, 42 (16.3%, [42/258]) to clade B, and 134 (51.9%, [134/258]) to clade E. All the body lice were clade A, while the head lice belonged to all three clades (A, B, and E).
The analysis of 84 cytb sequences yielded 44 variable positions defining 18 different haplotypes. Two haplotypes belonged to the worldwide haplotypes, A5 (14 head lice sequences and 5 body lice sequences) and A17 (5 head lice sequences), within clade A. Three haplotypes, all from head lice, also belonging to clade A, were novel and are named here as A60–A62. Within clade B, two haplotypes were found, one belonged to the B36 haplotype, the most widespread and prevalent in the B haplogroup, the second was novel and is referred to here as B39. The remaining 11 haplotypes belonged to clade E, all were novel, and they are named here as E56–E67 (Table 2). The phylogenetic position of these haplotypes is shown in Figure 1.
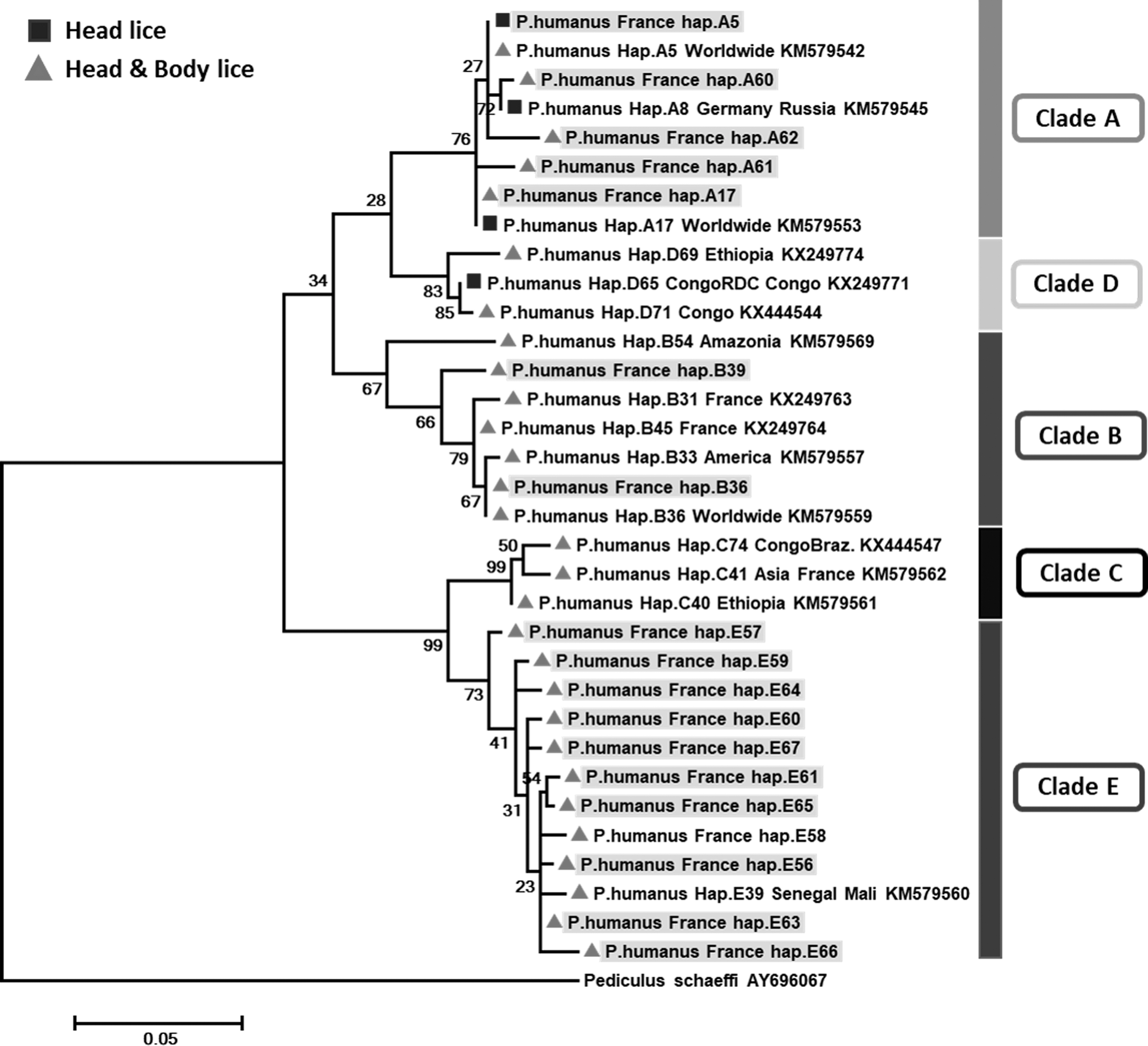
Phylogenetic tree showing the relationship of haplotypes identified in this study with other Pediculus humanus haplotypes. The cytb sequences were aligned using CLUSTALW and phylogenetic inferences were conducted in MEGA 6 using the maximum likelihood method, based on the Kimura 2-parameter. Statistical support for internal branches of the trees was evaluated by bootstrapping with 500 iterations. There was a total of 270 positions in the final dataset. The scale bar represents a 5% nucleotide sequence divergence.
The analysis of -cox1 sequences from 16 head lice belonged to clade E, yielded 6 variable positions defining 4 different haplotypes; all were novel named here as E40–E43 (Table 3). Phylogenetic tree (Fig. 2) showed that all these haplotypes clustered with haplotypes from Mali within clade E, confirming the identity of this clade.
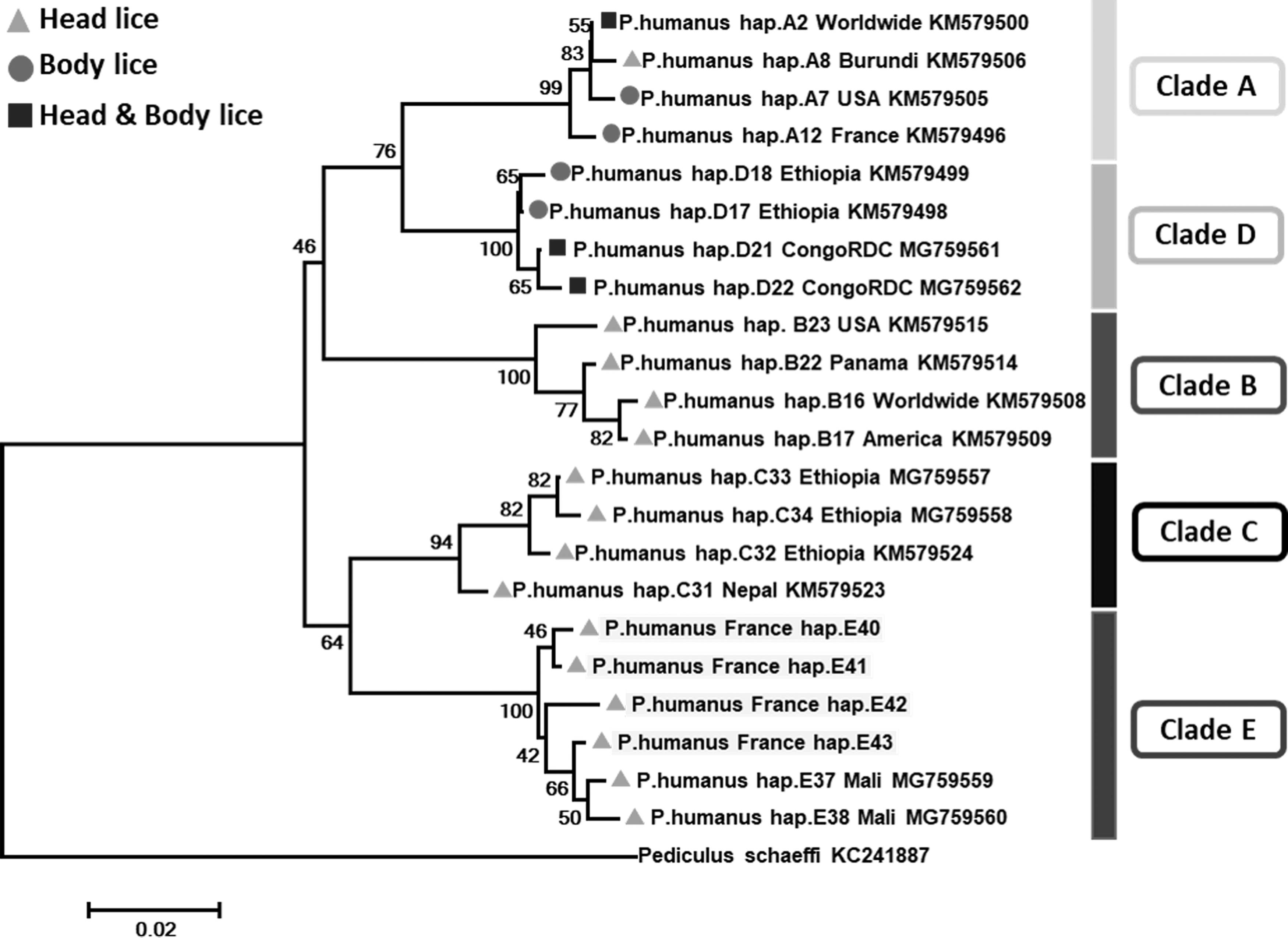
Phylogenetic tree showing the relationship of haplotypes identified in this study with other P. humanus haplotypes. The Cox1 sequences were aligned using CLUSTALW and phylogenetic inferences were conducted in MEGA 6 using the maximum likelihood method based on the Kimura 2-parameter. Statistical support for internal branches of the trees was evaluated by bootstrapping with 500 iterations. There was a total of 283 positions in the final dataset. The scale bar represents a 2% nucleotide sequence divergence.
Molecular detection of bacterial pathogens
In this study, the qPCR investigation of all lice samples for Rickettsia spp., R. prowazekii, Borrelia spp., Y. pestis, C. burnetii, and Anaplasma spp. produced no positive results. However, we obtained positive results when testing for the presence of B. quintana and Acinetobacter spp.
B. quintana DNA was found in 4 of 24 (16.66%) body lice collected from the same homeless patient. The patient was a 35-year-old man originally from Pakistan, who was hospitalized with an unexplained fever at the time of the sample collection. No head lice were found to be infected with B. quintana.
Acinetobacter DNA was detected by qPCR targeting the rpoB gene in 27 of 235 head lice (11.5%) collected from 8 of 47 families and in 7 of 8 body lice collected from the homeless patient 2, which represented a total of 29.1% (7/24) of all body lice tested. Among these 7 body lice Acinetobacter-DNA positive, 3 lice were also co-infected with B. quintana. Conventional PCR and sequencing targeting a 350-bp fragment of the same gene was successful only in 23 of the 34 samples that were positive in qPCR. This may be due to the lower sensitivity of standard PCR compared to qPCR.
Based on a BLAST search, seven sequences were identified as A. baumannii, two sequences as Acinetobacter nosocomialis, one sequence as Acinetobacter junii, and three sequences as Acinetobacter calcoaceticus, sharing 99–100% identity with their corresponding reference Acinetobacter species.
For 5 of the 23 sequences, BLAST analysis showed a homology score of under 95%, meaning that these sequences are likely to correspond to new species, provisionally referred to here as Candidatus Acinetobacter Bobigny-1 and Candidatus Acinetobacter Bobigny-2. The most closely related species are Acinetobacter johnsonii (GenBank number CP010350) for Candidatus Acinetobacter Bobigny-1 with 94% similarity (314 of 334 base positions in common), and Acinetobacter venetianus (GenBank no. LSVC01000004) for Acinetobacter Bobigny-2 with 91.9% similarity (307 of 334 base positions in common). The phylogenetic position of these Acinetobacter are shown in Figure 3. Interestingly, Candidatus Acinetobacter Bobigny-1 was identified in four clade E head lice collected from the same family, while Candidatus Acinetobacter Bobigny-2 was found in one clade B head louse from another family.
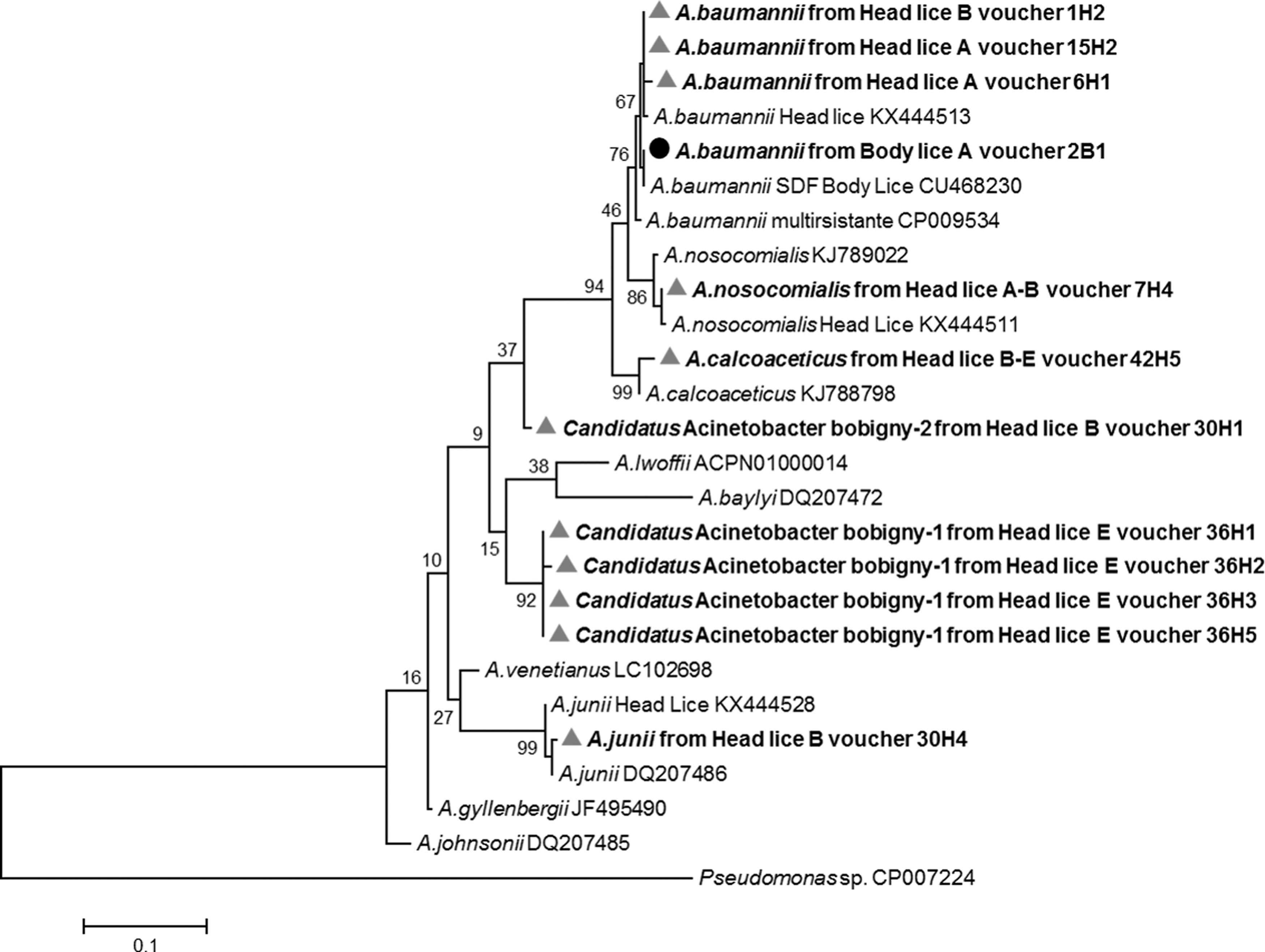
Phylogenetic tree highlighting the position of Acinetobacter species identified in head and body lice compared to another Acinetobacter available in the GenBank database. The rpoB sequences were aligned using CLUSTALW, and phylogenetic inferences were conducted in MEGA 6 using the maximum likelihood method based on the Kimura 3-parameter model for nucleotide sequences. The GenBank accession numbers are indicated at the end. Statistical support for internal branches of the trees was evaluated by bootstrapping with 1000 iterations. There was a total of 345 positions in the final dataset. The scale bar represents a 10% nucleotide sequence divergence.
The remaining 5 of 23 (21.7%) sequences, which were also rated, resembled Acinetobacter, but were of poor quality, which is assumed to be due to co-infection with several Acinetobacter species. The distribution of these species according to lice ecotypes and clades are presented in Table 4. The partial rpoB sequences obtained in this study were deposited in the GenBank under the accession no.: MF672011-MF672020.
Discussion
Louse infestation remains a social and public health concern around the world in the 21st century (Bonilla et al. 2013). In this study, the patients affected by pediculosis capitis were mainly female (sex ratio M/F = 0.1). During an epidemiological investigation in the same area, 70% (402/574) of infested children were girls and the median age was 8.9 years (Bouvresse et al. 2012).
The mtDNA analysis of the 235 head and 24 body lice showed the presence of three major haplogroups: A, B, and E. Haplogroup E was the most prevalent (51.9%, [134/258]), followed by haplogroups A (31.8%, [82/258]) and B (16.3%, [42/258]). All the body lice belonged to clade A. These data confirm the existence of clade A and B in France, as reported by others (Light et al. 2008a, Drali et al. 2015, Amanzougaghene et al. 2016b). Previous studies reported that clade E was limited to West African countries, namely Senegal and Mali (Ashfaq et al. 2015, Amanzougaghene et al. 2016b, 2017). This is the first report of clade E found in head lice in France, which may be explained by the intercontinental migration flow. Furthermore, clade C lice that were previously only observed in Africa and Asia have also recently been found in head lice collected in Paris (Drali et al., unpublished data). Our sampling failed to find this clade among the head lice tested, possibly due to the limited size and region of louse sampling.
B. quintana infection is the most common reemerging louse-borne disease in homeless populations living in poor hygiene conditions in developed countries, where it is responsible for a range of clinical manifestations in humans, including asymptomatic chronic bacteremia, endocarditis, and bacillary angiomatosis (Raoult and Roux 1999, Brouqui 2011, Badiaga and Brouqui 2012). In this study, we found B. quintana DNA in four of 24 body lice collected from one of the hospitalized homeless patients, consistent with the standard view that body lice are the major natural vector for this pathogen, which is among the most prevalent parasitic infestations in the homeless population (Raoult and Roux 1999, Badiaga et al. 2008). Epidemiologic studies of homeless populations have reported the prevalence of 7–22% for body louse infestations and 2–30% for B. quintana infections (Badiaga et al. 2008).
In recent decades, B. quintana DNA has also been detected in head lice collected from poor and homeless persons in the United States, Nepal, Senegal, Ethiopia, the Democratic Republic of the Congo, and France (Sasaki et al. 2006, Bonilla et al. 2009, Angelakis et al. 2011a, 2011b, Boutellis et al. 2012, Cutler et al. 2012, Drali et al. 2014, 2015). Conversely, all attempts to detect B. quintana in head lice collected from schoolchildren living in family households have failed (Fournier et al. 2002, Bouvresse et al. 2011, Sunantaraporn et al. 2015). Taken together, these results suggest that head lice probably can also transmit B. quintana if people live close together in poor sanitary conditions and if they lack medical treatment. This view is supported by our finding, as we were unable to detect the bacterium in any of the head lice collected from the 47 middle-class suburban families in the study.
In this study, we also assessed our collected lice for the presence of Acinetobacter species. More attention is now paid to extrahospital reservoirs of these ubiquitous opportunistic bacteria and their potential involvement in emerging human community-acquired infections, as pan drug-resistant strains are increasingly being identified worldwide (Eveillard et al. 2013).
Recent works have shown the Acinetobacter infection to be highly prevalent among body and head lice (La Scola and Raoult 2004, Bouvresse et al. 2011, Amanzougaghene et al. 2016a). However, it is still unknown how lice acquire these infections. Some authors argue that infections could occur after the ingestion of an infected blood meal from patients with ongoing bacteremia (La Scola and Raoult 2004) or possibly by passage through the human skin while feeding (Kempf et al. 2012). Furthermore, an experimental study showed that the human body louse, feeding on bacteremic rabbits, can acquire and support a persistent life-long infection with A. baumannii and A. lwoffii (Houhamdi and Raoult 2006). Another study compared two sequenced genomes of A. baumannii and showed that the A. baumannii homeless strain from the body louse had several hundred insertion sequence elements, which played a crucial role in its genome reduction compared to the human multidrug-resistant A. baumannii AYE strain, and which have also been shown to have low catabolic capacities, suggesting the specific adaptation of this strain to the louse environment (Vallenet et al. 2008).
Our results showed the presence of Acinetobacter DNA in 11.5% of head lice and 29.1% of body lice. Six species were identified, including two potential new ones. A. baumannii was the most prevalent, followed by Candidatus Acinetobacter Bobigny-1, A. calcoaceticus, A. nosocomialis, A. junii, and Candidatus Acinetobacter Bobigny-2. Body lice were found to be infected only by A. baumannii, while the head lice belonging to all three clades were found to be infected by at last one of all six species. As a result, it appears that Acinetobacter lice infection is not specific to a particular louse clade, but probably to louse ecotypes, as body lice were found to be infected only by A. baumannii. This hypothesis is supported by the detection of A. baumannii in 21% of body lice collected worldwide (La Scola and Raoult 2004). In contrast, studies performed on head lice collected from elementary school children in Thailand and from pygmy population in the Republic of Congo, showed the presence of the DNA of several Acinetobacter species, in 3.62% of head lice belonging to both clade A and C, and 37.3% of head lice belonging to clades A, D, and C (Sunantaraporn et al. 2015, Amanzougaghene et al. 2016a), respectively. In these two studies, the Acinetobacter species identified were A. junii, A. ursingii, A. baumannii, A. johnsonii, A. schandleri, A. lwoffii, A. nosocomialis, A. towneri, and A. radioresistens (Sunantaraporn et al. 2015, Amanzougaghene et al. 2016a). When comparing the panel of Acinetobacter species found in all these studies with our findings, A. ursingii, A. johnsonii, A. schandleri, A. lwoffii, A. towneri, and A. radioresistens were not identified in our head lice collection. Conversely, our sampling showed, for the first time, the presence of the DNA of A. calcoaceticus and two potentially new species in human head lice.
These studies together demonstrate the widespread infection of human lice with several species of Acinetobacter, suggesting that lice could be a preferential host for these bacteria. However, it remains to be determined whether these Acinetobacter strains present in lice are the same as those that are responsible for human infections.
In conclusion, we confirmed that head lice from Bobigny (France) belong to haplogroups A and B, and reported, for the first time, the presence of haplogroup E, which is specific to West Africa, reflecting the heterogeneous communities found in the studied area. We detected B. quintana only in body lice from homeless individuals, but not from head lice collected from the 47 middle-class suburban families. Several Acinetobacter species were also detected, including two potentially new ones, indicating that lice could be a source for Acinetobacter spp. infections in humans.
Footnotes
Acknowledgments
We gratefully thank IHU Fondation Méditerranée Infection for supporting the study.
Funding Source
This study was supported by Méditerranée Infection and the National Research Agency under the program « Investissements d'avenir », reference ANR-10-IAHU-03.
Authors' Contributions
Conceived and designed the experiments: O.M., F.F., and A.I.. Collected samples: K.C., A.I., and S.B. Conducted the experiments: K.C., N.A., O.M., and F.F. Analyzed the data: K.C., N.A., and O.M., and F.F. Wrote the article: N.A. K.C., O.M., F.F. A.I., R.D., M.L., and D.R.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.