
Abstract
Outbreaks of acute encephalitis syndrome (AES) with high fatality and disability, are reported every year in the Gorakhpur region of Uttar Pradesh, India, with the etiology of >60% of the cases being attributed to scrub typhus. In the present study, the prevalence of Orientia tsutsugamushi, the etiological agent of scrub typhus, was investigated among animal hosts and their ectoparasitic trombiculid mites prevalent in AES-reported areas of Gorakhpur. A total of 154 rodents/shrews were collected using 777 Sherman traps set in 12 study villages, and the overall trap rate was 19.8%. In total, 2726 trombiculid mites belonging to 12 species were collected from 154 rodents/shrews trapped. The shrew mouse Suncus murinus was the predominant animal species (78.6%) collected. The principal vector mite Leptotrombidium deliense was the predominant species (82.7%), and its index was 14.6 per animal. Of 114 rodent/shrew sera samples screened through the Weil–Felix test, 57% were positive for antibodies against O. tsutsugamushi. Of 128 blood samples tested by polymerase chain reaction (PCR), one rodent sample was positive for the gene encoding 56 kDa protein and 25 for 60 kDa. Among 2726 mite samples tested as 315 pools through nested PCR, seven pools were positive for 56 kDa gene. Phylogenetic analysis confirmed circulation of Gilliam, Karp, and TA678 serotypes of O. tsutsugamushi in Gorakhpur. The study clearly demonstrated natural infection of O. tsutsugamushi in both small-animal hosts and vector mites in the AES-reporting villages of Gorakhpur, which confirms transmission of the scrub typhus pathogen in this region. The high infestation rate of L. deliense with O. tsutsugamushi infection indicates that the people living in the rural villages of Gorakhpur are at risk of infection with scrub typhus, which might lead to AES.
Introduction
O
Scrub typhus is the most commonly occurring rickettsial infection in India, and the disease is caused by a Gram-negative bacterium, Orientia tsutsugamushi (Family: Rickettsiaceae). The infection is transmitted among small animals such as rodents and shrews and to humans by some species of larval mites (chiggers) belonging to the family Trombiculidae. The disease is known to occur in diverse ecological settings such as mountainous regions, rainforests, semi-arid deserts, seashores, river banks, terrain undergoing secondary growth, grasslands, and alpine subarctic terrains in the Himalayan regions (Kelly et al. 2002, Sharma et al. 2010). In a recent study, the scrub typhus pathogen was detected among small animals collected from the endemic areas of Puducherry and the adjoining areas of Tamil Nadu, India (Candasamy et al. 2016). In humans, the disease is generally an acute febrile illness involving multiple organs. Encephalitis is a late manifestation of the disease, with mortality in untreated patients (Saifudheen et al. 2012, Griffith et al. 2014). The disease is often undiagnosed mainly due to the fact that the clinical manifestations are nonspecific (Vivekanandan et al. 2010). Though the etiology of majority of AES cases in Gorakhpur was attributed to scrub typhus based on clinical and laboratory findings on human samples, evidence of the presence of the pathogen in animal hosts and vector mites are confirmatory to its transmission to humans. Therefore, this study was carried out to assess the prevalence of O. tsutsugamushi in small-animal hosts and their ecto-parasitic trombiculid mites collected from the villages of Gorakhpur district reported with human cases of AES.
Materials and Methods
Study area
Gorakhpur is a large district, located in the northeastern part of Uttar Pradesh between 26°46′N latitude and 83°22′E longitude. The district covers an area of 7483.8 km2 and is situated on the river basins of Rapti and Rohini. It is one of the 250 most backward districts in India. It has a population of 4,436,275 (2011 census), with a sex ratio of 944 females to 1000 males and a literacy rate of 73.25%. About 81.2% of the population in the district live in rural villages, and their main occupation (85.7%) is agriculture.
The climate of the district is moderate, with four seasons: hot summer (March to mid-June), southwest monsoon (mid-June to September), post monsoon (October to mid-November), and the winter season (mid-November to February). The mean minimum and mean maximum temperatures recorded during summer are 25°C and 40°C, respectively, and the average temperature is 32°C. During winter, the temperature ranges from 9°C to 32°C, and this season provides a pleasant atmosphere. During the rainy season, ∼87% of rainfall occurs due to the southwest monsoon, and the average annual rainfall in the district is 1393 mm.
A cross-sectional survey of trombiculid mites was carried out in 12 villages selected randomly from areas reported with recent cases of AES in Gorakhpur district, Uttar Pradesh, India (Fig. 1). A survey was conducted in six villages during July 2015 and in other six villages during October 2015, as the outbreak of AES coincides with the monsoon (mid-June to September) and post-monsoon (October to mid-November) periods.
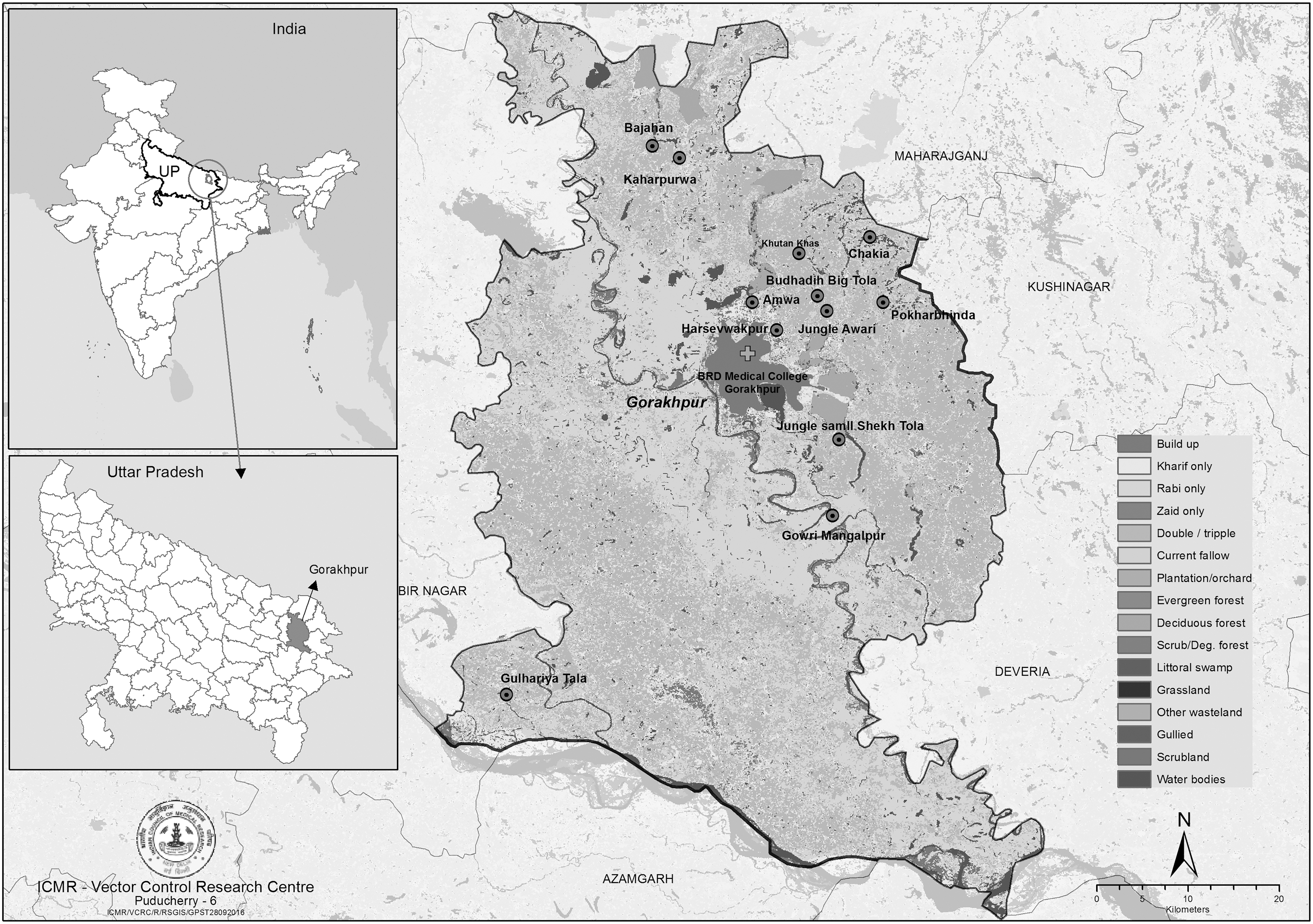
Gorakhpur district of Uttar Pradesh showing location of study villages.
Trapping and identification of rodents and shrews
Rodents/shrews were trapped by Sherman live traps (7.6 cm × 8.9 cm × 22.9 cm) baited with fried coconut and onion pakoda. In each of the selected village, about 65 traps were set outdoors (peri-domestic areas) 1 h before sunset (5.00 pm) and retrieved the following morning (6.00 am). The trapped rodents were euthanized with thiopentone sodium (Pentothal) injection (60 mg/kg intravenously) and identified after recording their morphological characteristics (Agarwal 2000). The study was approved by the Institutional Animal Ethics Committee, and handling of field-trapped rodents and shrews was carried out as per the guidelines of Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA).
Collection and identification of trombiculid mites
The ecto-parasites, including chigger (larval) mites, were collected by combing the animals against the fur over a white enamel tray. In addition, the ears, limbs, and axillary regions of individual rodents were examined for mite attachment under a stereomicroscope. If found positive, the ear pinna, limbs, and skin of the axillary regions were cut, and the mites were removed using fine forceps and transferred to 75% ethanol vials. Using a stereomicroscope (5 × ), individual mites were placed on drops of 75% ethanol kept on microscopic slides. The abdomen of each mite was punctured with a specially designed fine needle (that cuts the margin of abdomen), and their internal tissue contents were transferred using fine-tipped forceps into 1.5 mL Eppendorf tubes with 75% ethanol for polymerase chain reaction (PCR) assays. The exoskeletons of the mites were mounted in Hoyer's medium and identified to species level following standard taxonomical keys (Fernandes and Kulkarni 2003), and the tissue samples belonging to same-mite species collected from a particular study site and the same species of rodent/shrew were pooled (samples from 10 mites) and subjected to PCR assays.
Detection of antibodies of rickettsial pathogens in animals by serological assay
One milliliter of blood sample was collected from each of the trapped rodents/shrews through heart puncture without anticoagulant and the sera separated. The Weil–Felix test for antibody titer was performed against three antigens: OX-19 (Rickettsia typhi), OX-2 (Rickettsia conorii), and OX-K (O. tsutsugamushi; Bhattacharya et al. 2009; Progen–Tulip diagnostics). Samples that reacted with OX-19, OX-2, and OX-K antigens were considered to be positive for antibodies against murine typhus, tick typhus, and scrub typhus pathogens, respectively. Against all the above antigens, matt formation at 1:160 dilution was taken as positive for the respective antibodies (Kamarasu et al. 2007). Each time, matt formation was compared to that of a negative control and a known positive control.
Detection of O. tsutsugamushi DNA in mite and animal blood samples by PCR assay
Extraction of DNA from mite tissue and rodent/shrew blood samples was carried out using GenElute Blood Genomic DNA Kit (Sigma–Aldrich). The extracted DNA was used as templates for amplification using specific DNA primers. All the rodent/shrew blood samples were processed for amplification of two different gene fragments, groEL and 56 kDa type-specific antigen (TSA) of O. tsutsugamushi, by conventional PCR (Lee et al. 2003) and nested PCR (Lee et al. 2011), respectively, while the mite samples were processed by nested PCR assay.
Detection of gene encoding 56 kDa protein, which amplify 483 bp segments, was carried out by nested PCR following the method described by Lee et al. (2011). Primers used for the reaction were: F′-5′-TCAAGCTTATTGCTGAGTGCAATGTCTGC-3′; R′-5′-AGGGATCCCTGCTGCTGTGCTTGCTGCG-3′ for the first round, and F′-5′-GATCA AGCTTCTCAGCCTACTATAATGCCC-3′; R′-5′-CTAGGGATCCCGACAGAGCA CTATTAGGC-3′ for the second round reaction (nested PCR). The PCR amplification (Qiagen PCR Core Kit) was carried out in an Eppendorff thermal cycler, and the initial reaction mixture of 50 μL consisted of each of 10 pmol of forward and reverse primers, 5 μL of buffer, deoxynucleotide triphosphates (dNTPs), and 2 μL of extracted DNA and Taq Polymerase. The cycling conditions included an initial denaturation of 94°C for 5 min, followed by 31 cycles of 94°C for 50 s, 55°C for 2 min, and 72°C for 2 min, and a final extension of 5 min. The second reaction (nested PCR) consisted of 1 μL of the first PCR product, 20 pmol each of forward and reverse primers, 5 μL buffer, dNTPs, and 1 IU of Taq polymerase made up to 50 μL with deionized water. The annealing temperature set for the reaction was 62.5°C. Other conditions were the same as that of the first PCR. The presence of diagnostic amplicons were visualized on a gel documentation system (GelDocIT Imaging System, UVP) after electrophoresis on 1.5% agarose gel containing 0.5 μg/mL ethidium bromide.
The primers used for groEL gene PCR were F-5′-TTGCTGATGATGTAGACGGA-3′ and R-5′-TGTTCACAACGAGAATTAACTT-3′ for the amplification of a 307 bp segment (Lee et al. 2003). The PCR reaction mixture contained Go Tag green Master Mix (Promega), 10 pmol each of forward and reverse primers, and 4 μL of the DNA in a final volume of 30 μL reaction. Amplified 56 kDa and groEL gene fragments were sequenced in an ABI automated Genetic analyzer 3130 XL (Applied Biosystems). The DNA sequences generated were aligned using Clustal W and were analyzed using the phylogenetic software MEGA v7.0 using the maximum likelihood method to determine their serotypes and evolutionary relationships. Bootstrap analysis of 1000 replications was performed for the generation of a statistically valid analysis.
Data analysis
The trap success rate and chigger infestation index (average number of chiggers per animal) were estimated. The difference in the chigger indexes between villages was tested using one-way analysis of variance. The statistical analysis was carried out using STATA SE v9.0 (StataCorp, College Station, TX).
Results
Species diversity of small animals collected
A total of 154 rodents/shrews were trapped using 777 Sherman traps set in 12 study villages. The trap rate ranged from 9% to 39.3% in different villages, and the overall trap rate was 19.8%. Suncus murinus Linnaeus (Order: Soricomorpha) constituted 78.6% of the total animals trapped. The species of rodents collected were Mus booduga Gray (16.2%) and Rattus sp. (5.2%). Of 154 rodents/shrews screened, 83.4% of them were infested with trombiculid mites.
Species diversity of trombiculid mites
A total of 2726 mites belonging to 7 genera and 12 species of trombiculids were recovered from 154 rodents/shrews trapped (Table 1). Leptotrombidium deliense Walch was the predominant species (82.7%) followed by Schoengastiella ligula Radford (7.5%) and Walchia gujaratensis (Fernandes and Kulkarni 2003; 6.5%; Table 1). The proportion of L. deliense ranged from 9.9% in Gulhariya Tala village to 99.4% in Jungleshekh Tola village.
Figures in parentheses denotes percentage.
Chigger infestation index
The shrew species S. murinus contributed the maximum number (95%) of trombiculid mites, with the remaining contributed by the rodent species (Table 2). The overall chigger (all-mite species) index for S. murinus was 21.3 per animal, and it was 15.1 and 1.0 for Rattus sp. and M. booduga, respectively. The chigger (all-mite species) index estimated for different villages ranged from 1.2 to 80.1, and the overall index was 17.7 per animal. The estimated L. deliense index ranged from 0.6 to 79.3 in different villages surveyed, and the overall L. deliense index was 14.6 per animal. The Chigger index and L. deliense index varied significantly between the villages (chigger index: F = 3.13, p-value <0.001; L. deliense index: F = 3.86, p-value <0.001).
Prevalence of rickettsial pathogens in rodents/shrews
In the Weil–Felix test, 65/114 (57%) samples tested were positive for antibodies against O. tsutsugamushi. The number of positives for O. tsutsugamushi was greater in S. murinus than Rattus sp. The reactivity of 41 samples to Weil–Felix antigens was suggestive of the presence of antibodies to murine/tick typhus pathogens (Table 3).
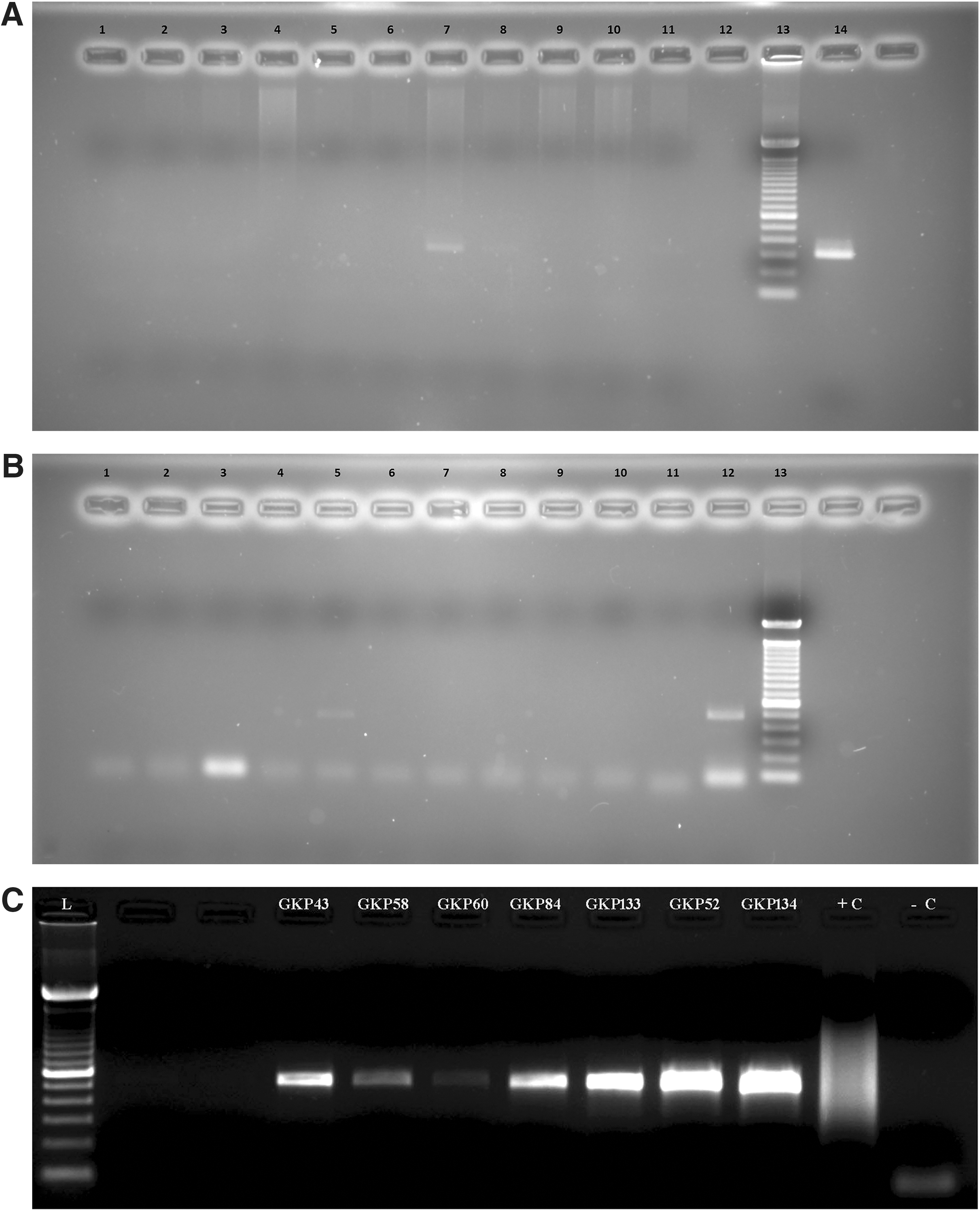
Of the 128 blood samples tested by PCR assay, 25 samples were positive for the groEL gene (307 bp; Fig. 2A) and one sample for the 56 kDa gene (483 bp; Fig. 2B). The identities of these genes were confirmed as that of O. tsutsugamushi by DNA sequencing and were submitted to GenBank (acc. nos.: KX130950 for 56 kDa gene and KX432184–KX432185 for groEL gene).
(
Detection of O. tsutsugamushi DNA in mite samples
Mite (internal tissue) samples of the same species collected from a particular site and the same rodent/shrew species were pooled for PCR assays. A total of 2726 mite samples (12 species) pooled in 315 pools (10 mite samples per pool) were tested by nested PCR for O. tsutsugamushi DNA. Of these, seven pools of L. deliense were positive for 56 kDa gene (483 bp; Fig. 2C). All the O. tsutsugamushi infected L. deliense specimens were collected from S. murinus.
Phylogenetic analysis
Phylogenetic analysis of seven 56 kDa-positive samples revealed that three were genetically related to Karp, another three to TA678, and one to Gilliam serotype (Fig. 3). Village-wise analysis revealed that one Gilliam, two TA678, and three Karp strains were detected in mite vectors collected from Gowri Mangalpur village, and one TA678 strain was detected in Budhadih Tola village (Table 4). Phylogenetic analysis of eight randomly selected groEL positive samples revealed that seven samples were related to Karp-like strains and one sample to the Gilliam-type strain (Fig. 4). Analysis of village-wise distribution of the O. tsutsugamushi strains showed that Karp-like strain was found in five of the study villages and the Gilliam-type strain in one village (Table 4). Analysis of the 56 kDa sample showed a similarity with the Gilliam-type strain, and it was detected in a rodent sample collected from Chakia village (Table 4).
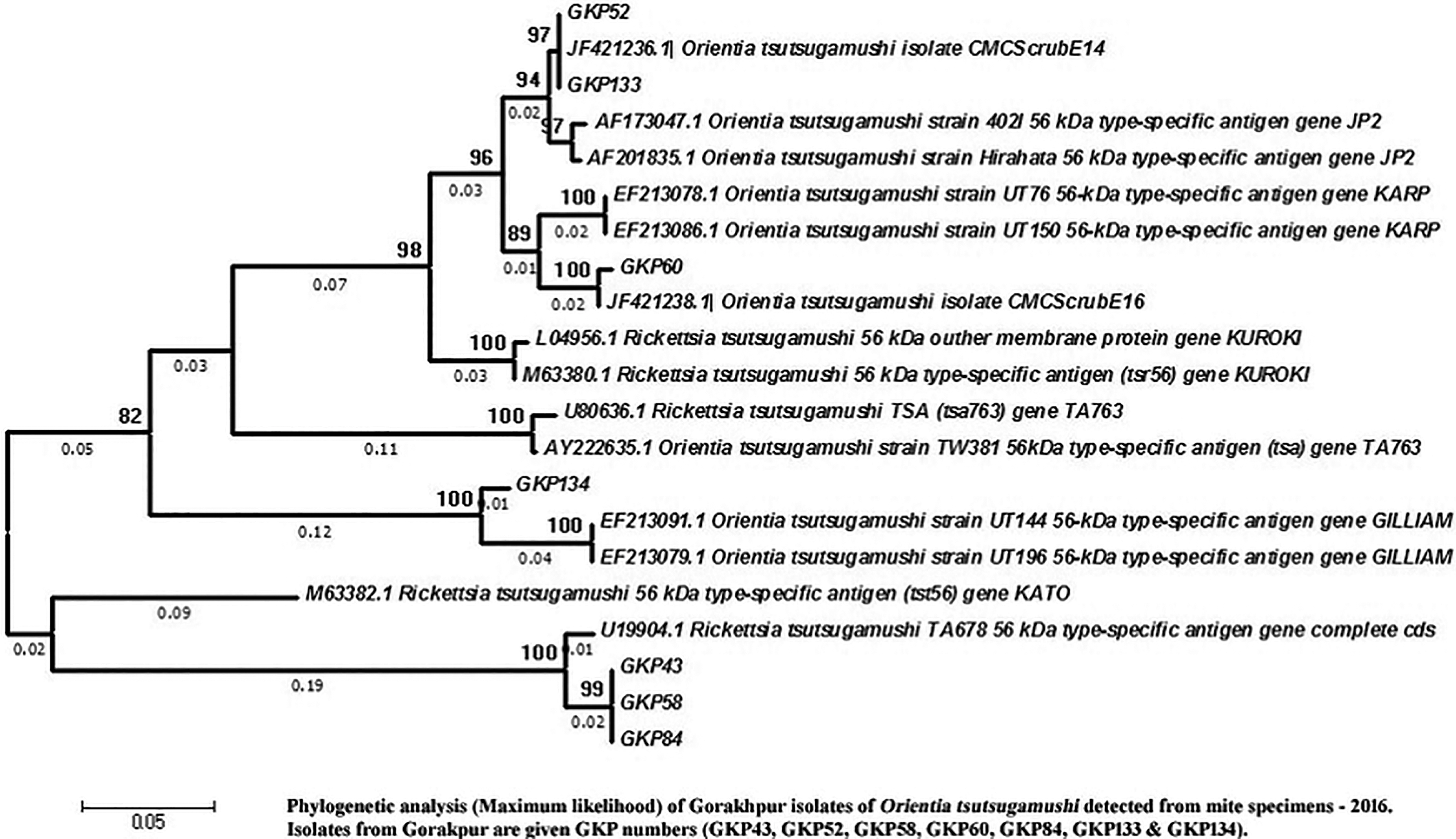
Phylogenetic analysis (maximum likelihood) of Gorakhpur isolates of O. tsutsugamushi detected from mite specimens 2015. Isolates from Gorakhpur are given as GKP numbers (GKP43, GKP52, GKP58, GKP60, GKP84, GKP133, and GKP134). Figure 3 can be viewed in greater detail online at

Maximum likelihood–based phylogeny of groEL gene of O. tsutsugamushi from rodent blood samples depict the clustering of Gorakhpur isolates with Karp- and Gilliam-related strains.
Discussion
In India, endemic AES has been reported from 171 districts, distributed in 17 states, and around 375 million people are at risk (Potharaju 2012). Outbreaks of AES have been reported from Gorakhpur region since 1978. Every year, 1500–2000 cases with a 20–25% fatality rate occur (Murhekar et al. 2016, Shrivastava et al. 2016). Serological investigation of 365 AES cases reported during September–October 2015 revealed IgM scrub typhus positivity in 63% of the AES cases, and the results suggested that scrub typhus could be an important etiology of AES (Murhekar et al. 2016). However, evidence on the occurrence of O. tsutsugamushi, the causative agent of scrub typhus in small-animal hosts and chigger mite vectors, was lacking. This study provided information on the relative abundance of small-animal hosts, abundance of chigger mite vectors, and the prevalence of the scrub typhus pathogen, O. tsutsugamushi, in the animal and vector hosts.
The shrew mouse S. murinus was the predominant animal species collected, accounting for >75% of the live captured animals. This contributed to the maximum number (95.6%) of L. delicense, and the estimated chigger (L. deliense) index for this species was 17.8 per animal. In the present study, very few Rattus sp. were collected. The low occurrence of Rattus sp. observed in the study villages might be due to the trap placement (i.e., in peri-domestic areas). The estimated infestation rate of L. deliense for Rattus sp. was considerably higher next to S. murinus. Analysis of blood samples of rodents/shrews by PCR assays also confirmed that S. murinus was the animal species naturally infected with O. tsutsugamushi. Occurrence of O. tsutsugamushi infection in small animals has been reported from Southeast Asian countries (Frances et al. 1999). In India, natural infection of O. tsutsugamushi has been documented in rodent and shrew species collected from scrub typhus endemic areas of Puducherry and adjoining areas of Tamil Nadu (Candasamy et al. 2016). In the present study, O. tsutsugamushi infection could not be established in Rattus sp., as only three samples were screened by PCR assay.
L. deliense was the most abundant species among the 12 species of trombiculid mites collected. L. deliense is the established vector of the scrub typhus pathogen in many of the endemic countries, including India (Kalra 1952, Oaks et al. 1983, Lee et al. 2011, Rodkvamtook et al. 2013). The Chigger indexes, one of the major attributes of the scrub typhus transmission potential, varied greatly in different villages studied. The chigger (L. deliense) index recorded in the present study is significantly greater than that reported in Megalaya (Sharma 2013a), Kerala (Sharma 2013b), and Kolkata (Sharma and Kumar 2014). In Gulhariya tala village, where a single case of AES/scrub typhus was reported, the prevalence of L. deliense (9.9%) was low compared to other study villages. However, the estimated L. deliense index for this village was significantly higher than reported in other epidemiological settings.
Out of 12 species of trombiculid mites screened by nested PCR, natural infection of O. tsutsugamushi was found only in L. deliense sp., confirming that this species as the vector of O. tsutsugamushi in the study villages. All the O. tsutsugamushi–infected L. deliense specimens were collected from the shrew species S. murinus.
In this study, the higher positivity rate observed by groEL gene-specific PCR might be due to the high degree of nucleotide similarity in the groEL gene among different strains of Orientia, which ranges from 95.74% to 99.6% (Lee et al. 2003). On the contrary, the hyper-variable 56 kDa TSA gene has greater genetic variability among the strains of Orientia, which may require more copies of the same strain of Orientia for amplication. This is also corroborated by the sensitivity of nested PCR assay that varies from 62% to 90% (Paris et al. 2009). In the study area, phylogenetic analysis of nucleotide sequences of O. tsutsugamushi 56 kDa gene from mite samples showed the prevalence of Karp, Gilliam, and TA678 strains. Analysis of rodent samples showed the presence of Gilliam and Karp strains, as observed in mite samples. Phylogeny in human cases revealed the presence of Gilliam and Karp strains (NIV, Gorakhpur, unpublished data). Thus, the finding suggests that Gilliam, Karp, and TA678 are the common strains of O. tsutsugamushi circulating among humans, rats, and vectors in Gorakhpur region. The prevalence of Karp and Gilliam serotypes has been documented in Southern and Eastern India (Varghese et al. 2015), and the third serotype, TA678 antigenic type, has been reported from South Korea (Kang and Chang 1999) and Thailand (Kelly et al. 2009). Information on such local antigenic diversity of O. tsutsugamushi pathogen is essential for the development of region-specific vaccines and diagnostics (Chattopadhyay and Richards 2007).
The rural villages of Gorakhpur, with extensive agricultural land, provide a favorable environment for the breeding and survival of host animals and their ecto-parasitic vector mites that contribute to the transmission of the scrub typhus pathogen to humans living in their proximity. S. murinus, the shrew species, is known to live in close proximity to human dwellings, and the extent of domiciliation may be an important criterion in rural areas for the transmission of O. tsutsugamushi and human cases of scrub typhus manifesting as AES.
Conclusion
The present study clearly demonstrates the prevalence of O. tsutsugamushi infection in both the animal hosts and their ectoparasitic mites collected from AES endemic areas of Gorakhpur, which confirms transmission of the scrub typhus pathogen. The high infestation rate of mite vector L. deliense with natural infection of O. tsutsugamushi indicates that the people living in the rural villages of Gorakhpur are at high risk of acquiring the disease, which might lead to AES.
Footnotes
Acknowledgments
The technical assistance rendered by Mr. Sri Hari Murmu and M. Stalin, Vector Biology and Control division, and Mr. Kumaresan, Molecular biology division of the Centre, is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.