
Abstract
In recent years, the number of human rabies cases in China has decreased annually. However, some western provinces with no human cases for more than 10 years have begun to report rabies cases, and all of the rabies lineages that circulated in western China were found in Inner Mongolia as well. In this study, we generated a phylogenetic tree with all the Inner Mongolia rabies strains available in GenBank and our laboratory, as well as strains from western China and representative viruses from neighboring countries, based on the N gene sequence. Furthermore, the possible relationships underlying the spread of the virus within Inner Mongolia and neighboring regions were analyzed. Three of six rabies lineages of China (China I–VI) were shown to exist in Inner Mongolia, and a spatial cluster analysis supported that the China I lineage, the dominant cluster of China, likely spread to Ningxia from Inner Mongolia. Wild raccoon dog rabies (China IV/Arctic-like-2) may have spread to Inner Mongolia from Russia and likely continued to spread to Qinghai and Tibet. The red fox lineage (China III/Cosmopolitan), which likely spread from Russia and Mongolia, has been shown to circulate in Inner Mongolia and was a serious threat to Xinjiang, which is adjacent to Inner Mongolia. Thus, Inner Mongolia likely became a location where national and international rabies viruses collected and developed into a potential portal for the spread of rabies to western China. To effectively control the spread of rabies in China, both prevention and control of dog and wild animal rabies in Inner Mongolia should be a top priority.
Introduction
R
In 2002–2004, more than 1000 human cases were reported annually, and rabies became the leading cause of death among the 37 notifiable infectious diseases in China (Song et al. 2014, Zhou et al. 2016). For this reason, a national rabies surveillance program was implemented in 2005 to investigate the outbreak in terms of vaccination coverage, PEP treatment, and geographical and social composition. A number of preliminary control measures were implemented, such as raising public awareness and increasing the availability of normative PEP (Song et al. 2014). As a result, the number of annual fatalities decreased, with 644 cases reported in 2016 (Song et al. 2014, Zhou et al. 2016, Taxitiemuer et al. 2017). Although the number of annual fatalities has fallen each year since 2007, gradual geographic spread of rabies has been reported (Yin et al. 2012, Yu et al. 2012). Especially in western China, new cases have been recorded in areas that had been free of rabies for many years (Tao et al. 2015, Zhou et al. 2016), thus presenting a new problem that needs to be addressed.
The ongoing rabies surveillance program in China has provided a comprehensive overview of the current outbreak, and current surveillance analysis has shown that rabies viruses of western China, including Ningxia, Qinghai, Tibet, and Xinjiang, demonstrate a close relationship with those of Inner Mongolia (Feng et al. 2014, Tao et al. 2015, Liu et al. 2016), which is characterized by various rabies lineages (Shao et al. 2011, Yin et al. 2014).
Thus, this study summarized the sequences of all rabies strains of Inner Mongolia available in GenBank and our laboratory and constructed a phylogenetic tree with strains in neighboring regions based on the N gene sequence of the rabies virus. Furthermore, the possible source and direction of spread of rabies viruses of Inner Mongolia were inferred, and the significance of Inner Mongolia to rabies control was emphasized.
Materials and Methods
Ethics statement
The program for the collection of cattle brain specimens was approved by the Ethics Committee of the National Institute of Viral Disease Control and Prevention, Chinese Center for Disease Control and Prevention (China CDC), which is the national referral center for rabies diagnosis. Cattle brain specimens were obtained following suspected rabies deaths.
Epidemiological data
Data on cases of human rabies from different provinces of Mainland China were collected from the Chinese Notifiable Disease Reporting System (NDRS) of the China CDC. The reporting methods and the manner in which the cases were determined to be associated with rabies were the same as described previously (Yu et al. 2012, Song et al. 2014, Zhou et al. 2016).
Sequence data of the rabies viruses of Inner Mongolia obtained from GenBank
Sequence data of street rabies strains of Inner Mongolia (as of June 1, 2017) were verified in GenBank, and only isolates with full background information (isolation time, host, and location) were selected for analysis. Only the N gene sequence was available for all of the 24 candidate strains of Inner Mongolia (Supplementary Table S1; Supplementary Data are available online at
Specimen detection and sequencing
As the reference laboratory for national rabies surveillance in China, our laboratory has submitted and analyzed sequences of more than 200 rabies samples in patients and animals since 2005 (Wang et al. 2008, Tao et al. 2009, 2013, 2015, Lang et al. 2012, Yu et al. 2012, Guo et al. 2013), including three samples from Inner Mongolia (Supplementary Table S1) (Yin et al. 2014). In this study, we continued to detect and sequence newly collected samples from Inner Mongolia for subsequent analysis. Two cattle brain samples collected in 2015 from Bayannur, Inner Mongolia, were detected using a direct immunofluorescence assay, as described previously (Tao et al. 2009). Both samples were positive; these were named CNM1501C and CNM1502C.
Total RNA extraction and cDNA synthesis were performed as described previously (Tao et al. 2009, Lang et al. 2012), and the complete sequence of the N gene was amplified and sequenced using two prime pairs, as described previously (Tao et al. 2009, 2015). The two N gene sequences (1353 bp) were submitted to GenBank with the accession numbers MF741703 and MF741704.
Dataset of the N sequence rabies viruses
To gain perspective regarding the Inner Mongolia sequences, the different lineages of China, and rabies strains of neighboring provinces and countries, a complete dataset of rabies N gene sequences (n = 65, 1353 nt) was created, which included 26 sequences from Inner Mongolia (including 2 new sequences), 27 representative sequences from the six lineages of China (including strains from Ningxia, Gansu, Qinghai, Tibet, and Xinjiang), 11 sequences from neighboring countries, and 1 bat sequence (United States) as the outgroup (Supplementary Table S1).
Phylogenetic analysis
The sequences were aligned using Clustal X software (version 2.1) (Thompson et al. 1997); phylogenetic reconstruction was performed using the MEGA 7 software package with both the neighbor-joining (NJ) method and the Maximum-Likelihood (ML) method (Kumar et al. 2016). The Tamura 3-Parameter (T92) model (Tamura 1992), inferred by MEGA 7, was applied to build the ML tree. The topology support of the NJ tree and the ML tree was tested with 1000 and 100 bootstrap replicates, respectively.
Spatial cluster analysis
In recent years, rabies has spread to western China, and our previous study found that the emergence of the disease in Ningxia and Gansu was influenced by neighboring provinces (Inner Mongolia and Shaanxi) as reflected in the geographical distribution of human cases and phylogenetic analysis (Tao et al. 2015).
To further explore the possible relationship between Inner Mongolia and neighboring provinces, Local Indicators of Spatial Association (LISA) analysis was used to evaluate the spatial clusters of the reported rabies cases at the city level in four provinces (Inner Mongolia, Ningxia, Gansu, and Shaanxi). Since more than one human cases were first reported (up to nine) in Gansu in 2013 (Tao et al. 2015), the morbidities for each city of the four provinces in 2013 were downloaded from the NDRS of the China CDC (Supplementary Table S2). By calculating the Local Moran's I coefficient, which typically ranges from −1 to 1, the spatial correlations between the data at the level of a local area unit and the average of neighboring values in the surrounding units are revealed on LISA cluster maps (Anselin 1995, Li et al. 2016).
After standardization of Local Moran's I coefficient, the standardized Z-score was used to assess the significance of the observed spatial correlations. With LISA, it is possible to identify five classes of spatial grouping: High–High (zone with high risk surrounded by zones with high risk), Low–Low (zone with low risk surrounded by zones with low risk) (both cases for Z-score >1.96), Low–High (zone with low risk surrounded by zones with high risk), High–Low (zone with high risk surrounded by zones with low risk) (both cases for Z-score <−1.96), and nonsignificant correlations among risks assigned to each vicinity zone (−1.96 < Z < 1.96) (Parra-Amaya et al. 2016). The spatial statistical analysis module of ArcGIS software (version 9.3; ESRI, Redlands, CA) was used to perform the LISA analysis (ArcGIS 2018).
Results
Rabies epidemics in Inner Mongolia and western provinces of China
Rabies is a zoonotic disease, and rabies virus can affect almost all terrestrial mammals, including humans (Rupprecht et al. 2017). Since the establishment of the People's Republic of China in 1949, cases of rabies in humans have been reported annually (Zhou et al. 2016). In 1996, the lowest values for the number of cases of rabies in humans and the total area of epidemic regions in China were at their lowest, and rabies was limited to only a few southern provinces (Song et al. 2014, Zhou et al. 2016). No cases were reported in Inner Mongolia in 1998, and two or fewer cases were reported from 1999 to 2006 (Tao et al. 2013). Subsequently, a sudden increase in the number of cases was reported in 2007. After 2013 (14 cases), the number of cases began to decline (Table 1). Almost no human cases were reported in the five western provinces of China (Xinjiang, Gansu, Ningxia, Qinghai, and Tibet) before 2011 (Table 1) (Tao et al. 2013). Ningxia and Gansu began to report cases in 2011 and 2013, respectively (Tao et al. 2015), and the first human cases in Qinghai and Tibet were reported in 2012 and 2015, respectively (Table 1).
The case numbers greater the “0” are in bold.
Animal rabies surveillance in China is lacking (Song et al. 2014) and needs to be supported by laboratory tests. Although detailed information regarding animal cases of rabies is unavailable, the animal strains confirmed by detection and sequencing can, to a certain degree, reflect the situation. The earliest report of animal strains found in Inner Mongolia was concerning raccoon dog rabies in 2007 (Shao et al. 2011). The first rabies strain identified in Xinjiang was in a sheep bitten by a fox in 2013 (Feng et al. 2014). Gansu, Qinghai, and Tibet reported their first rabies strains from dogs in 2012 and 2013 (Tao et al. 2015), and Ningxia reported its first animal rabies strain (from a camel) in 2015 (Liu et al. 2016).
During this epidemic period, beginning in 1996 (Tao et al. 2013), Inner Mongolia was considered an epidemic region until about 2007, and the five western provinces began to report human or animal cases ∼4 years later. It must be admitted that this information is based on the limited animal surveillance data available.
Lineage diversity of rabies viruses of Inner Mongolia
The rabies virus strains from Inner Mongolia were used to generate a phylogenetic tree with the representative strains of different rabies lineages from China (Fig. 1), which can clarify the lineages circulating in Inner Mongolia. Both the NJ tree (Supplementary Fig. S1) and the ML tree (Fig. 1) exhibited similar clustering patterns. The 26 strains of Inner Mongolia identified during 2007–2015 were divided into three lineages (China I, China III, and China IV), which represent half of the China lineages (Guo et al. 2013, Tao et al. 2013), and different lineage strains exhibited different relationships with the domestic and international strains (Fig. 1).
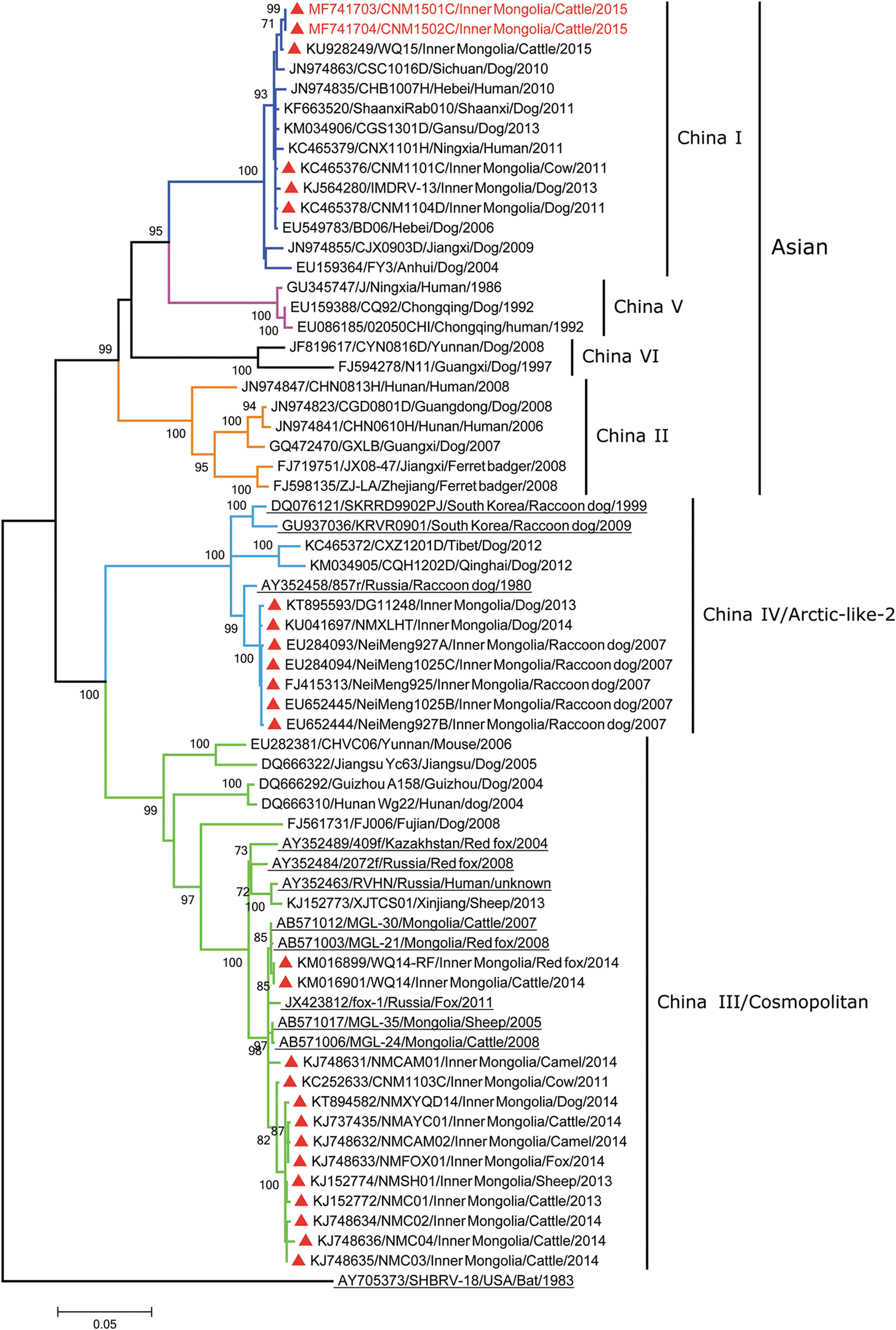
A maximum-likelihood phylogenetic tree of the rabies street strains of Inner Mongolia and related strains based on the N gene sequences. A scale indicating genetic distance is presented by the horizontal bar. Branches are colored to indicate the six different lineages circulating in China (dark blue, China I; orange, China II; green, China III; sky blue, China IV/Arctic-like-2; pink, China V; black, China VI). Taxa are in the format (ACCESSION NO/STRAIN/CHINA PROVINCE or COUNTRY/HOST/YEAR). The strains from Inner Mongolia are highlighted with red triangles, and the two strains newly sequenced in this study are highlighted with red taxa. The international strains are underlined.
The China I lineage is predominantly responsible for the rabies epidemic in China beginning in 1996 (Guo et al. 2013, Tao et al. 2013). This lineage is distributed throughout almost all the eastern and southern provinces, as well as some northern regions (Lang et al. 2012, Tao et al. 2013). Six dog or cattle strains of Inner Mongolia were included in this lineage, and three showed a close relationship with Ningxia, Hebei, Gansu, and Shaanxi strains (Fig. 1). In a previous study, the possible source of strains from Ningxia and Gansu was examined in detail (Tao et al. 2015). Following 8 years with no recorded cases in humans, Ningxia reported two cases of rabies in 2011, and a strain obtained from the saliva of one patient (CNX1101H) was determined to be most similar to the dog strain collected from Inner Mongolia the same year (CNM1104D) (Fig. 1) (Tao et al. 2015), suggesting that the emergence of rabies in Ningxia may have resulted from the spread of the rabies virus from neighboring Inner Mongolia (Tao, et al. 2015). The China I lineage has slowly spread to western China through neighboring high-incidence regions, which is also evident in the close relationship between the strains found in Gansu and Shaanxi provinces (Tao et al. 2015). In addition, the remaining three strains of Inner Mongolia are associated with strains from a more distant region (Sichuan) (Fig. 1) and are related to the current rabies viruses that have spread to western China.
The China III lineage corresponds to the international “Cosmopolitan” lineage (Bourhy et al. 2008, Xue et al. 2013, Troupin et al. 2016) and has been reported in some areas of China, including Yunnan, Jiangsu, and Hunan (Guo et al. 2013, Tao et al. 2013). However, in recent years (2011–2014), this lineage was found only in Inner Mongolia and Xinjiang and was identified in local livestock, dogs, and red foxes (Fig. 1 and Supplementary Table S1). These sheep, cattle, and camels were almost all bitten by foxes (Feng et al. 2014, Liu et al. 2016), and their strains, as well as the local fox strains, demonstrated a close relationship with the fox strains of Russia and Mongolia, although the strains from Xinjiang and Inner Mongolia were divided into two branches (Feng et al. 2014).
The China IV lineage corresponds to another international lineage “Arctic-like-2” (Kuzmin et al. 2008, Pant et al. 2013, Tao et al. 2013), which is also a wild animal rabies lineage. Five strains from domestic raccoon dogs on a farm in Inner Mongolia (Shao et al. 2011), likely transmitted by a local wild raccoon dog in 2007 (Shao, et al. 2011), and two local dog strains during 2013–2014 demonstrated a close evolutionary relationship with raccoon dog strains of Russia (Fig. 1). In addition to Inner Mongolia, Qinghai and Tibet reported this same lineage (Tao et al. 2015), although these strains were not significantly close to those of Inner Mongolia (Fig. 1).
Spatial clusters of rabies cases in Inner Mongolia and neighboring western provinces of China
It was inferred based on the geographical distribution of human cases and phylogenetic analysis (Tao et al. 2015) that rabies could have spread from Inner Mongolia to Ningxia and from Shaanxi to Gansu. This was also supported by the “LISA” analysis (Supplementary Table S3 and Fig. 2). The red areas were identified as hot spots (High–High), where significant spatial clusters of a high rabies incidence exist. A significant spatial cluster appeared near the border between Inner Mongolia and Ningxia, which suggests that the China I lineage likely spread to Ningxia through Inner Mongolia (Fig. 2B). The distribution of rabies hot spots in Gansu and Shaanxi suggests that the spread of the China I lineage from neighboring regions with high incidence to regions with low incidence is common.

Geographical location of Inner Mongolia, with neighboring provinces and countries
Discussion
WHO claims that the elimination of human rabies cases transmitted by dogs is feasible through mass vaccination of domestic dog populations (WHO 2017). However, this is difficult to carry out for China, given its vast territory and unbalanced regions. Although much has been done to raise public awareness and increase widespread availability of normative PEP (Song et al. 2014), which has decreased the incidence of rabies in humans (Song et al. 2014, Zhou et al. 2016), the rabies virus continues to spread to northern and western China and has reached nearly all provinces of Mainland China (Tao et al. 2015).
In recent years, hot spots characterized by rabies emergence were almost all located in western China (Guo et al. 2013, Tao et al. 2013, 2015, Feng et al. 2014, Liu et al. 2014), and Inner Mongolia was the only location that demonstrated a close relationship with all of the epidemic strains identified in these hot spots.
Inner Mongolia is a wide frontier province of northern China that shares borders with Mongolia and Russia. Unlike other border areas in China, such as Tibet, which is blocked by the Himalayan Mountains, Inner Mongolia connects with neighboring provinces and bordering countries across the Mongolian plateau through steppe and forest ecosystems (Fig. 2). At the same time, there are many other carnivorous hosts in addition to dogs, including foxes, raccoon dogs, and wolves (Feng et al. 2014, Wang et al. 2014), which are beneficial for the propagation and dispersion of the rabies virus.
Inner Mongolia has likely been important in the spread of dog rabies to low-incidence regions such as western China. Like many rabies epidemic countries, ∼95% of Chinese rabies cases in humans result from virus transmission through the bites of infected dogs (Song et al. 2014, Zhou et al. 2016, WHO 2017). The China I lineage of rabies virus mainly corresponds to the rabies epidemic in humans and dogs, and it has spread to northern areas from the southern China over the past 20 years (Lang et al. 2012, Guo et al. 2013, Tao et al. 2013). Since 2011, Inner Mongolia has reported more than 10 human cases of rabies annually (Table 1), and Inner Mongolia has become a potential danger to regions of low incidence. Ningxia began to report human cases again in 2011, after being human rabies free for 8 years. This was likely the result of the spread of China I lineage strains from Inner Mongolia by dogs, a conclusion that is supported by epidemiology, as well as phylogenetic and LISA analyses (Figs. 1 and 2). Inner Mongolia is not the only place responsible for the spread of rabies virus to western China; China I lineage viruses likely spread from Shaanxi to Gansu (Figs. 1 and 2) (Tao et al. 2015). However, there are long border areas between Inner Mongolia and Gansu, and Gansu, Qinghai, Tibet, and Xinjiang are under threat from neighboring regions with a high incidence of rabies. Inner Mongolia is a potential doorstep for the spread of the China I lineage (dog rabies) to western China.
Moreover, wild animal rabies of Inner Mongolia probably is likely a threat to western China. Fox rabies (China III lineage) has been shown to circulate in both Inner Mongolia and neighboring Mongolia and Russia (Fig. 1) (Feng et al. 2014), where wild animals (e.g., red foxes and raccoon dogs) remain an epidemic threat (Botvinkin et al. 2008, Deviatkin et al. 2017). Rabies in Inner Mongolia likely spread from, or was at least deeply influenced by, Mongolia and Russia. According to limited sequence data, wild animal rabies of Xinjiang have been influenced by Russia and Kazakhstan (Fig. 1) (Feng et al. 2014), but the substantial risk from neighboring Inner Mongolia cannot be neglected.
In addition, with respect to the China IV lineage, strains from Inner Mongolia were also much closer to the Russia strain (Fig. 1), suggesting the possible source of raccoon dog rabies in Inner Mongolia. The rabies strains from Qinghai and Tibet and the raccoon dog strains of Inner Mongolia belong to the Arctic-like-2 lineage (Fig. 1), which circulates primarily in Russia and other East Asian countries (Kuzmin et al. 2008). A reasonable speculation is that Russian raccoon dog rabies slowly spread to the Qinghai–Tibet Plateau through Inner Mongolia and that the viruses found in Qinghai and Tibet experienced an independent evolution for a while, as they are not significantly close to the strains from Inner Mongolia and Russia (Fig. 1), although it will be necessary to collect additional samples to support this hypothesis. The raccoon dog rabies currently present in Inner Mongolia is also at risk for spread to western China.
Both dog and wild animal rabies were widely observed in Inner Mongolia and were likely a major threat to western China, which still exhibits a low incidence of rabies. Therefore, further and more comprehensive surveillance should be conducted to improving our understanding of the rabies situation in Inner Mongolia, and prevention and control of dog and wild animal rabies in Inner Mongolia is a top priority for the elimination of rabies in China. To this end, efforts in and collaboration among China, Mongolia, and Russia are essential.
Footnotes
Acknowledgments
This work was supported by the National Program on Key Research Project of China (2016YFD0500400), the National Science and Technology Major Project (2017ZX10104001), the National Natural Science Foundation of China (31500152), the National Key R&D Program of China (2017YFC1200503), and the National Science and Technology Major Project (2018ZX10201002). The authors thank Xiao-Long Li for assistance with data analysis.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.