
Abstract
Chagas disease is a lingering Public Health problem in Latin America with ∼5.7 million people infected with Trypanosoma cruzi. Transmission is still taking place in most countries of the Americas, including the United States. Dogs are frequently infected with T. cruzi and its high infection prevalence is associated with increased risk of Chagas disease in humans. The city of Mérida in the Yucatan peninsula is endemic for Chagas disease and canines are frequently infected with T. cruzi. The objective of this study was to evaluate the performance of a qualitative point of care (POC) molecular test (RPA-LF, recombinase polymerase amplification-lateral flow) developed in our laboratory for identifying infected dogs. We used retrospective samples of dogs that came for consultation because of cardiac alterations and proved to be infected with T. cruzi as determined by enzyme-linked immunosorbent assay (ELISA), Western blot, and quantitative PCR (qPCR). The analytical sensitivity indicated that RPA-LF amplified T. cruzi DNA in samples containing almost equal to one to two parasites per reaction. Serial twofold dilutions of T. cruzi epimastigotes showed that the test had 95% (19/20) repeatability at concentrations of two parasites per reaction. The test showed no cross reactivity with human DNA or other protozoan parasites (Trypanosoma rangeli, Leishmania spp., and Plasmodium spp.). RPA-LF had the capacity to amplify all discrete typing units (DTUs I–VI) of T. cruzi that circulate in domestic or extradomestic environments. The RPA-LF had 93.2% (95% confidence interval 87.2–98.1) sensitivity and excellent agreement with qPCR used as gold standard (Cohen's Kappa test = 0.963). ELISA was positive in 96.6% (85/88) of dogs, which together with the molecular tests confirmed the frequent contact with infected triatomine bugs in the city of Mérida. These preliminary results on the diagnostic efficacy of the RPA-LF deserve further large-scale field testing of this POC test for T. cruzi infection in endemic areas.
Introduction
C
Chagas disease has extended from strictly rural to urbanized environments where new reservoir hosts are playing an important role in the transmission cycle (Gurtler and Cardinal 2015). Dogs are frequently infected with T. cruzi and its high infection prevalence is associated with an increased risk of Chagas disease in humans (Beard et al. 2003, Gurtler et al. 2007).
Furthermore, dogs are considered reservoirs and victims of T. cruzi since its infection could lead to cardiomyopathies and sudden deaths like humans (Caldas et al. 2013, Vitt et al. 2016). In Argentina, the probability of seropositive dogs to get infected with the vector Triatoma infestans was 12-fold higher compared to seropositive children and 100-fold higher than seropositive adults (Gurtler et al. 1996). The clinical evaluation of these dogs indicated that poor nutritional status and decreased cell-mediated immunity were positively correlated with infectivity to T. infestans (Petersen et al. 2001). On the other hand, the infectiousness increased when the animals were co-infected with Ancylostoma caninum and an unidentified trematode (probably Fasciola hepatica) (Enriquez et al. 2016). A serological survey in the city of Mérida, Mexico, also showed high serological prevalence (>75%) of T. cruzi infection among dogs suffering different cardiomyopathies (Ortega-Pacheco et al., unpublished).
It is clear that intervention on the canine reservoir of Chagas disease is a key component of surveillance and control programs. A short turnaround time between diagnosis and intervention, eliminating the need of delivering samples to central or specialized laboratories, should strengthen the efficacy of control strategies. The aim of this study was to evaluate the performance of a POC molecular test (recombinase polymerase amplification [RPA]) for identifying dogs infected with T. cruzi.
Materials and Methods
Study sites
All dogs were from Mérida, Yucatan, Mexico (19′30″ and 21′35″N latitude, and 87′30″ and 90′24″ W longitude). Housing in many neighborhoods favors triatomine infestation and sustains active T. cruzi transmission (Guzman-Tapia et al. 2007, Jimenez-Coello et al. 2010).
Dog samples
We evaluated 88 naturally infected dogs brought for consultation at the Faculty of Veterinary Medicine and Animal Science of the Autonomous University of Yucatan (UADY), Mérida. The reason for consultation was the existence of cardiomyopathies.
Blood samples were obtained by venipuncture of the cephalic vein. Three milliliters of whole blood was collected in PAX-gene (cat. no. 761125; BD-Qiagen) to preserve DNA until purification. An additional 3 mL sample without anticoagulant was obtained using sterile Vacutainer tubes and centrifuged at 2000 g for 10 min to collect serum. Both, purified DNA and sera were stored at −20°C until use (Jimenez-Coello et al. 2015).
Serology for anti-T. cruzi IgG detection
Serum samples were evaluated using an enzyme-linked immunosorbent assay (ELISA; Chagatest-ELISA recombinant v.4.0 kit; Wiener Laboratories S.A.I.C.). This ELISA test detects antibodies to six recombinant proteins expressed in T. cruzi. The assay was carried out following the manufacturer's recommendations, except for the second antibody that was replaced with goat anti-dog IgG conjugated with horseradish peroxidase (HRP; sc-2433; Santa Cruz Biotechnology). Briefly, 96-well plates were coated with recombinant proteins, and then sequentially incubated with 20 μL of serum samples (1:80 dilution) in phosphate buffer (137 mM NaCl, 2.7 mM KCl, 4.3 mM Na2HPO4, and 1.4 mM KH2PO4, pH 7.4) and HRP-conjugated dog anti-IgG (1:5000 dilution). Color was developed with tetramethylbenzidine and hydrogen peroxide substrates, and the reaction was stopped by acidification of the reaction medium. The optical density (OD) was read at 450 nm in an xMark™ microplate absorbance spectrophotometer (Bio-Rad, Hercules, CA). The cutoff value of 0.217 was determined from the mean value of the negative control sera ±3 standard deviations (Medina 2002).
DNA extraction
DNA was purified from whole blood samples according to Jalal et al. (2004). Subsequently, we used the protocol of the commercial kit, DNeasy Blood and tissue kit (69504; Qiagen, Germantown, MD), following the manufacturer instructions. Total DNA was examined for quality (OD260/OD280 ratio of 1.7–2.0) and quantity ([OD260 − OD320] × 50-μg/mL) using a DU® 800 ultraviolet/visible spectrophotometer.
Quantitative PCR
The quantitative PCR (qPCR) was performed with SsoAdvanced Universal SYBR Green Supermix (172-5271; Bio-Rad) and oligonucleotides (TCZ-F 5′-GATCTTGCCCACAMGGGTGC-3′ and TCZ-R 5′-CAAAGCAGCGGATAGTTCAGG-3′) as previously described (Schijman et al. 2011). Samples were evaluated by qPCR for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene to detect the potential presence of inhibitors in the reaction mix and to use equal amounts of DNA in all reactions.
Duplicate samples of total DNA (4 μL) were used as template for qPCR to estimate the T. cruzi loads. The standard curve was prepared using T. cruzi epimastigotes, clone CL Brener, TcVI as described by Bua et al. (2012). Briefly, 1 mL of blood from a seronegative dog was spiked with 107 parasites/mL and the total DNA was extracted. Tenfold serial dilutions of the extracted DNA, corresponding to 106–0.1 parasite/mL (corresponding to 1 × 103 to 1 × 10−4 parasite equivalents per assay), were used for running the qPCR (Cencig et al. 2011). Total DNA from healthy dogs was used as negative controls. All experiments included the parasite standard curve, DNA from negative controls, and no-template DNA control. The parasite loads were determined by linear regression analysis of the cycle threshold (Ct) values of the test samples against Ct values of the standard curve based on known amounts of T. cruzi DNA. The results are presented as Ct values and T. cruzi per mL. A melting curve analysis was performed in all qPCR-positive samples to confirm the presence of only one peak in each sample.
RPA and lateral flow reading
Design of primers and probe
The primer sets we designed for T. cruzi are 30–32 nucleotides long and target conserved sequences identified by computational alignment of T. cruzi satellite sequences reported in GenBank. Primers were designed with 40–60% GC content, few direct/inverted repeats, and absence of long homopolymer tracts. We focused on conserved regions and to a lesser extent on regions with moderate variability, obtaining a 146 base pair (bp) RPA amplicon in agarose gels. To enable detection by lateral flow (LF; as described below), the reverse primer was biotinylated at the 5′ end. We designed a 45 bp conserved internal probe (Biosearch Technologies, Petaluma, CA) that included FAM (5′-carboxy fluorescein amidite) at the 5′ end, an internal dSpacer, and a SpacerC3 in the 3′ end, as suggested by the manufacturer (TwistDx). Therefore, the primer set and probe used in this work were as follows: Fw03 5′ GCTGCACTCGGCGGATCGTTTTCGAG 3'; Rev 05 5′ GTTTGGTGTCCAGTGTGTGAACACGCAAACA 3′; and Probe-FAM-5′-GCACCACACGTTGTGGTCTAAATTTTTGTTTCGAATTATGAATGG-3′.
RPA reaction and LF reading
The amplification mixture per reaction comprised the following: (1) forward primer (0.89 mM), (2) biotinylated reverse primer (0.89 mM), (3) FAM-labeled probe (0.22 mM), (4) magnesium acetate (1.25 μL), and (5) the rehydrated cocktail (14.5 μL; Twist amp nfo RPA kit; TwistDx). Template DNA (5–25 ng/μL) of parasite-spiked samples or clinical samples was immediately added to the mixture and subjected to amplification at 40°C for 30 min using a dry bath. The RPA product was diluted at 2% in the dipstick assay buffer and 100 μL was placed in a 1.5 mL microtube. The bottom tip of the LF strip (Ustar Biotechnologies, Hangzhou, China) was then immersed in the sample, making the amplification product run upwards by capillarity. Parasite amplification was confirmed with the naked eye after 5 min by the appearance of the test band in the lower part of the strip. The positive test band is produced when anti-biotin antibodies immobilize the amplified DNA, which contains the biotinylated reverse primers. The gold particles in the strip, which are covered with mouse anti-FAM antibodies, bind to the FAM-labeled probe making the test band visible. The reaction was validated by the appearance of the control band in the upper part of the strip. This band appears upon the immobilization of excess free gold particles (which are covered with mouse antibodies) by means of anti-mouse antibodies. Positive and negative controls were included in each round of RPA-LF.
Sensitivity and specificity of RPA-LF
The sensitivity of the test was established using normal dog blood spiked with serial dilutions (106–0.1 parasite/mL) of T. cruzi epimastigotes of clone CL Brenner, TcVI, as template. To determine the specificity, the RPA-LF was run using Trypanosoma rangeli, Leishmania mexicana, Leishmania braziliensis, Leishmania infantum, Plasmodium falciparum, and Plasmodium vivax together with the six T. cruzi discrete typing units (DTUs I–VI). In silico analysis showed that the RPA primers did not align with Trypanosoma caninum. This recently described that nonpathogenic trypanosome was isolated from dog skin only in Brazil (Madeira et al. 2014).
Ethical statement
Blood samples from naturally infected dogs were collected with the consent of their owners after explaining the objectives of the study. The study was approved by the Bioethics Committee of the Campus de Ciencias Biológicas y Agropecuarias, Universidad Autónoma de Yucatán (CB-CCBA I-2017-003).
Results
ELISA was positive in 96.6% (85/88) of dogs, confirming the frequent contact with infected triatomine bugs in the city of Mérida. All the dogs included in the study were positive by qPCR. The three dogs that were serologically negative resulted positive by qPCR and RPA-LF (dogs No. 54, 58 and 74; Supplementary Table S1; Supplementary Data are available online at
The initial analytical sensitivity indicated that RPA-LF amplified T. cruzi DNA in samples containing 1–2 parasites per reaction, which corresponded to Ct values of 27 or 26 in the real-time PCR used as gold standard (Fig. 1). Serial two fold dilutions of T. cruzi epimastigotes showed that RPA-LF had 95% (19/20) repeatability at concentrations of two parasites per reaction.
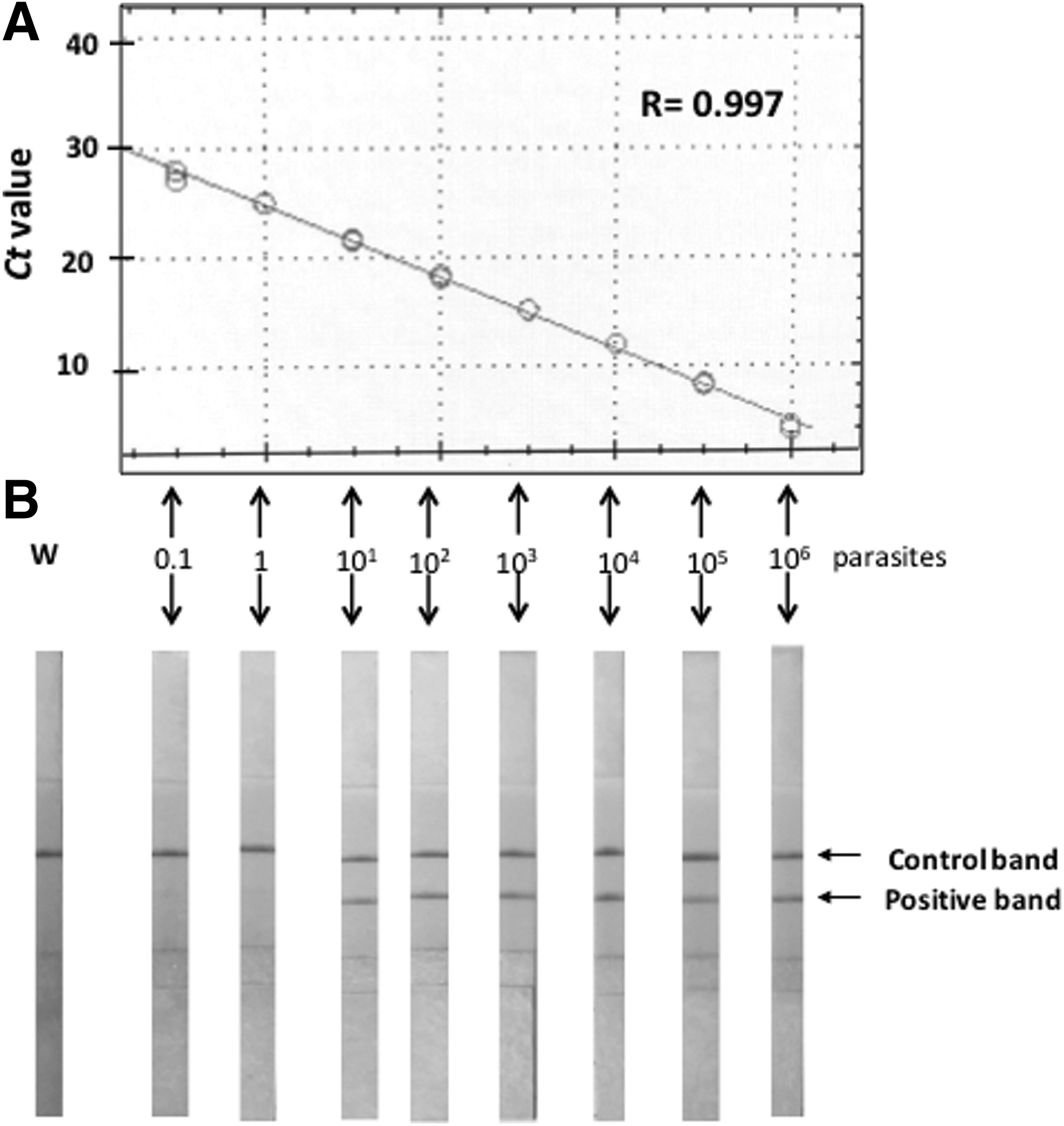
Analytical sensitivity of RPA-LF to detect Trypanosoma cruzi. Tenfold serial dilutions of parasite DNA were extracted with Qiagen DNeasy blood and tissue kit, and used to develop a standard curve upon amplification by qPCR (SYBRgreen; gold standard)
RPA-LF showed good specificity when run in the presence of human DNA or other protozoan parasites (Leishmania spp. and Plasmodium spp.) (Fig. 2A and Supplementary Fig. S1). There was no cross-reactivity with the closely related, but nonpathogenic T. rangeli, which infects different mammals, including dogs and humans. Of epidemiological relevance was the capacity of RPA-LF to amplify all DTU's (I–VI) of T. cruzi that circulate in domestic and extradomestic environments of different countries (Fig. 2A).
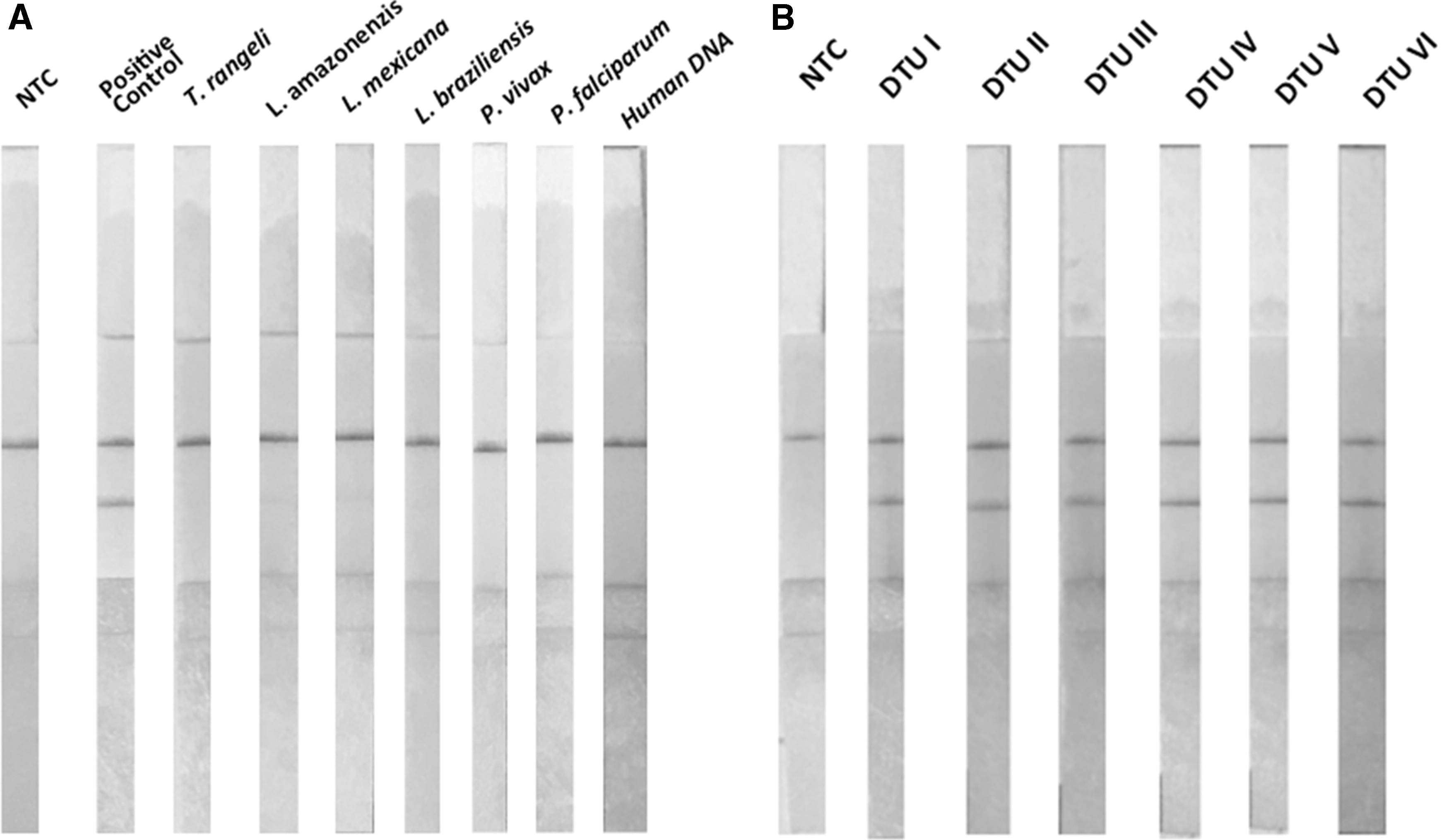
Specificity of RPA-LF and capacity to amplify all DTUs of T. cruzi. Species of Trypanosomatidae (Leishmania amazonensis, Leishmania mexicana, and Leishmania braziliensis), Plasmodiidae (Plasmodium vivax and Plasmodium falciparum), or human DNA were not amplified by the RPA-LF test
We determined the diagnostic efficacy of RPA-LF using DNA from retrospective blood samples obtained from 88 dogs inhabiting the city of Mérida that came for consultation to the Faculty of Veterinary Medicine and Animal Science (UADY), Mérida. Most of these infected dogs (confirmed by qPCR; Supplementary Table S1) presented cardiomyopathies, but not all of them were compatible with Chagas disease. The RPA-LF detected T. cruzi DNA in 82 of the 88 samples, reaching a sensitivity of 93.2% (95% confidence interval 87.2–98.1) and excellent agreement with qPCR (Cohen's Kappa test = 0.963). Four of the six dogs that resulted negative by RPA-LF had low parasite burden as indicated by the high Ct values (≥28.28) of qPCR (Supplementary Table S1). No RPA-LF false positive results were found when DNA samples from uninfected dogs or blood spiked with unrelated pathogens were included in the RPA-LF runs.
Discussion
We designed and evaluated an RPA-LF test for detecting T. cruzi DNA in dogs. The sensitivity was assessed using serial dilutions of T. cruzi spiked in dog blood. We determined the specificity of the test using DNA samples of other trypanosomatids and Plasmodium spp. The diagnostic applicability of RPA-LF was evaluated using blood samples of dogs naturally infected with T. cruzi.
Our primers targeted the kinetoplast DNA (kDNA) minicircles or satellite DNA since both parasite targets have been considered the basis for molecular diagnosis of Chagas disease in humans (Schijman et al. 2011). The satellite DNA has ∼120,000 copies (Vargas et al. 2004), while the kDNA minicircle contains 10,000–20,000 copies (Degrave et al. 1988, Sturm et al. 1989). In our hands, the RPA-LF test that targeted the satellite DNA showed higher reproducibility to amplify all T. cruzi DTUs compared to other amplification targets like kDNA (Schijman et al. 2011).
The diagnosis of Chagas disease relies mostly on serological methods. Indirect hemagglutination, ELISA, and indirect immunofluorescent antibody tests are still widely used (Schijman et al. 2011). Conventional serological tests were successfully adapted for canine diagnosis with good sensitivity and specificity results (Lauricella et al. 1998, Cardinal et al. 2006). However, none of the serological tests alone can unequivocally diagnose Chagas disease, requiring that two tests with different principles result positive to confirm T. cruzi infection (Enriquez et al. 2013, WHO 2015). An additional drawback of all serological tests is the cross-reactivity with other trypanosomatids (T. rangeli and Leishmania spp.) in areas where these parasites co-circulate (Enriquez et al. 2013, Dias et al. 2016). We used an ELISA kit to reach a preliminary diagnosis of Chagas disease in dogs presenting with different cardiomyopathies, which was then confirmed using molecular methods.
The dogs used in this study do not represent the actual prevalence of T. cruzi infection in the canine population of Mérida since they were brought for consultation due to cardiac alterations. However, all these animals demonstrated to be infected with T. cruzi, suggesting that urban transmission is occurring at a high rate, as recently observed in a cross-sectional study in Mérida (34% infection prevalence (Jimenez-Coello et al. 2010). Dogs can develop fatal cardiological alterations compatible with acute or chronic Chagas disease, principally in infected puppies (Vitt et al. 2016). Cardiac hypertrophy, arrhythmias, and bundle branch blockades similar to humans are found in naturally infected dogs (Barr et al. 1989, Barr 2009). Although canine cardiomyopathies were not the focus of this study, it is worth noting that 48% (40/84) of these animals had cardiac alterations compatible with Chagas disease, 23% characteristic of the acute-indeterminate phase, and 60% related to the chronic phase (Supplementary Table S1). In this group of dogs, no race could be identified as more prone to develop chagasic cardiomyopathy since the higher number of Chiguagua and Poodle may reflect dog-owner preferences for these races (Supplementary Table S2).
We believe that RPA-LF could be implemented in resource-limited areas lacking sophisticated healthcare infrastructure. The test could be accomplished in less than an hour, requiring only minimal plastic ware and a water bath or dry bath. We estimated that the total cost of using the latter DNA extraction method and the T. cruzi RPA-LF test is approximately USD 3.00.
Ideally, all POC molecular tests, including RPA-LF, need complementation by an efficient and simple-to-use DNA extraction method. Of concern is the possibility that the test sensitivity in the field is hampered by the utilization of simple, but less efficient DNA extraction methods. Published work with different microorganisms or human blood suggested that simple extraction methods could be available for resource-limited settings, yet its applicability needs to be demonstrated (Mahalanabis et al. 2010, Govindarajan et al. 2012, Gan et al. 2014). We have successfully detected L. infantum in dogs by extracting DNA from blood adsorbed in filter paper using a 30-min boiling step in DNAse-free water (Travi et al. unpublished). However, the samples were from polysymptomatic animals that putatively harbored high numbers of parasites. Alternatively, utilization of Whatman FTA filter papers to collect blood samples followed by washings with FTA buffer, suspension in TE buffer, and a 30-min boiling should be evaluated at the POC. In our hands, the evaluation of DNA samples purified from 1 mL of blood showed high detection efficiency of T. cruzi when using molecular diagnostic tests.
Testing of three consecutive samples by qPCR has shown to increase significantly the sensitivity (from 75.6% to 95.6%) to identify pregnant women infected with T. cruzi (Bisio et al. 2011). This would be logistically challenging in canine field studies, but feasible in a veterinary office. In the latter case, conventional PCR or qPCR could be used to detect markedly low numbers of T. cruzi, which are typically found during the chronic phase.
The current T. cruzi RPA-LF, like all qualitative LF tests, has no internal amplification control. We plan to optimize the test by including an internal DNA control that will be read in an additional LF band, thereby ruling out the presence of inhibitors of DNA amplification leading to false negative results.
In this work, the analysis of a well-characterized number of clinical samples indicated that RPA-LF can detect almost equal to two parasites per reaction with 95% repeatability. We expect that RPA-LF will efficiently identify Chagas disease in dogs with clinical manifestations. However, the efficacy of RPA-LF for detecting subclinical infections needs to be defined. Data on the natural course of Tc infections in dogs were based only on cross-sectional studies and therefore there is no precise information of the parasitological evolution spanning from the acute to the chronic stage of infection. Results from experimental infections were variable due to different infection protocols and Tc strains, but in general, the parasitemia became patent at 14 days postinfection (p.i.) and significantly decreased after 30–45 days p.i. (Machado et al. 2001, Bahia et al. 2002, Quijano-Hernandez et al. 2013).
The advantage of POC tests is the support they bring to rapid interventions upon implementation of control strategies. To our knowledge, control programs have focused principally on the elimination of triatomine vectors, but no attempts have been made to intervene on infected dogs, despite its role as both sentinel animals and reservoir hosts (Gurtler et al. 1996, Castillo-Neyra et al. 2015). A sensible control strategy would be to identify dogs with established infectious potential and remove them from the transmission cycle or significantly decrease their infectivity by treatment or immunotherapy. It is possible that a suboptimal vaccine could curb vector-mediated transmission from reservoirs to humans, provided it decreases parasite burden below infectivity levels, as suggested by recent vaccine trials in dogs (Aparicio-Burgos et al. 2015, Beaumier et al. 2016). It will be important to determine the prognostic value when positive or negative RPA-LF results are positive by qPCR. We hypothesize that infected canines with parasitemia below the RPA-LF detection limit could be significantly less infectious than RPA-LF-positive dogs harboring higher parasite burdens. The prospective implementation of the test in control programs will require assessing the utility of RPA-LF as a tool for determining the reservoir status.
Additional studies could evaluate the efficacy of this test to detect epimastigotes or trypomastigotes of T. cruzi in feces of triatomine bugs as a rapid screening method of vector populations or to evaluate xenodiagnosis results of humans or dogs.
The capacity of RPA-LF to amplify all the DTUs that circulate in different countries suggested that this diagnostic tool could aid epidemiological studies of both urban and sylvatic cycles of T. cruzi in Latin America. Our preliminary results on the diagnostic efficacy of the RPA-LF warrant further large-scale field testing of this fast, accurate, and inexpensive test for T. cruzi infection in endemic areas.
Footnotes
Acknowledgments
This work received partial support from The Center for Tropical Diseases-CTD, University of Texas Medical Branch, Galveston; PRODEP (Program for Professional Teacher Development, for University Academics) Registration number SISTPROY FMVZ-2016-0011, Mexico.
Author Disclosure Statement
The authors do not have conflicts of interest related to this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.