
Abstract
This study investigated the abundance of mosquitoes and circulation of mosquito-borne arboviruses from 16 villages in 8 cities of Hunan Province, China, in July–August of 2010 and in August of 2011. In total, 16,076 mosquitoes consisting of seven species from four genera were collected by ultraviolet-light trap. Culex quinquefasciatus was the most common species, accounting for 50.63% (8140/16,076) of the total. Anopheles sinensis (24.26%, 3900/16,076) made up the second most common species, followed by Culex tritaeniorhynchus (9.76%, 1569/16,076). The proportions of Culex pipiens pallens, Armigeres subalbatus, and Culex modestus were 6.7%, 5.2%, and 3.31%, respectively. Fourteen Aedes albopictus were detected. The mosquitoes were identified by morphologic characteristics and frozen in liquid nitrogen. The mosquitoes were pooled, triturated, and centrifuged. The clarified supernatant was used to inoculate monolayers of C6/36 and baby hamster kidney-21 cells. We obtained six virus isolates that caused cytopathic effects. Phylogenetic analysis revealed that two isolates were Akabane virus (AKAV, from A. sinensis and C. quinquefasciatus), two isolates were Japanese encephalitis virus (from C. pipiens pallens and C. quinquefasciatus), and two isolates were Tibet orbivirus (from C. quinquefasciatus and C. tritaeniorhynchus). This is the first report of AKAV isolated from A. sinensis and C. quinquefasciatus in nature in China. The detection of AKAV in these species confirms circulation of AKAV in Hunan province and suggests potential challenges to the prevention and control of arthropod-borne animal viruses in mainland China.
Introduction
M
Hunan Province (108°47′–114°15′E and 24°39′–30°08′N), located in the southern part of China, has a humid subtropical climate with high average temperatures and heavy rainfall (Government of Hunan Province 2017). Hunan Province has numerous rivers and abundant water systems and is located near Dongting Lake, the second largest freshwater lake in China. The prolonged high temperatures and rainy weather in the summer provide an optimal geographical environment for breeding and propagation of various mosquitoes. Mosquito species surveys in Hunan Province show that the mosquito density begins to increase from June, peaks in mid-August, and decreases in mid-September; the mosquito density decreases significantly after October. Culex tritaeniorhynchus, Anopheles sinensis, Culex quinquefasciatus, and Armigeres subalbatus are the dominant mosquitoes in the area (Dai et al. 2015).
Hunan Province is a JE endemic area. From 1951 to 2000, JE cases were reported every year, and a total of 142,190 cases were reported with 18,328 deaths. The average annual incidence rate was 5.82/100,000 with a 12.89% fatality rate and a 0.75/100,000 mortality rate (Guo and Li 2002). In addition, JEV genotype I was isolated from mosquitoes collected in Yueyang, located northeast of Hunan Province (Xiang et al. 2017). To understand the circulation of mosquito-borne viruses in Hunan Province, we focused on the identification of mosquito species and mosquito-borne viruses from 16 collection sites in 8 cities of Hunan Province in 2010 and 2011. AKAV, JEV, and Tibet orbivirus (TIBOV) were isolated in this study. AKAV was isolated from A. sinensis and C. quinquefasciatus for the first time in mainland China.
Materials and Methods
Mosquito collections
Mosquitoes were collected from 16 villages in Huaihua, Zhangjiajie, Yueyang, Chenzhou, Xiangtan, Shaoyang, Zhuzhou, and Changsha cities of Hunan Province (Fig. 1). It was the first time that investigation on arboviruses was carried out in Hunan Province. Collection sites were selected based on the proximity to the breeding sites of mosquitoes and in villages inhabited by 10,000–20,000 people and containing pigpens, cattle farms, and ponds. These environments are suitable for the circulation of arboviruses in nature.
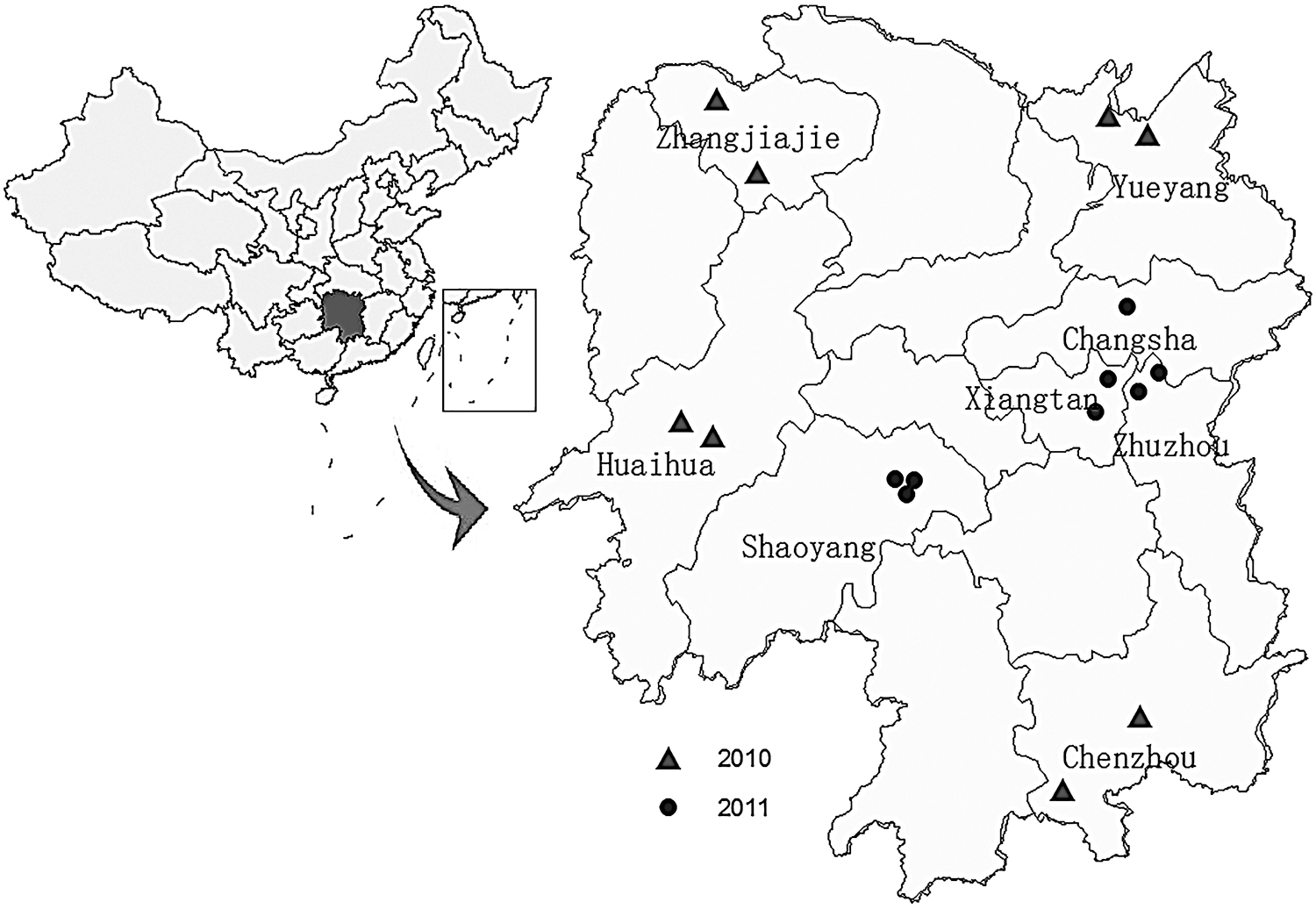
Locations of mosquito collection sites in Hunan Province, China (triangles: collection sites in eight counties in 2010; spots: collection sites in eight counties in 2011). The map of China (upper left) shows the location of Hunan Province, China.
Mosquitoes in every collection site were trapped for two nights from 20 July to 6 August of 2010 and in 1–17 August of 2011. Mosquitoes were collected from 18:00 in the evening to 6:00 the following morning using ultraviolet (UV)-light traps (Wuhan Lucky Star Environmental Protection Co., Ltd.). The UV-light traps were placed in the pigpens and under the eaves of the houses on sites, where the traps were hung about 2 m high. Female mosquitoes were identified by species using morphologic keys (Lu 2003) and pooled into groups by species, date, and site of collection. Approximately 50 mosquitoes were placed in each pool and frozen in liquid nitrogen (Wang et al. 2011).
Cell culture
Aedes albopictus C6/36 cells and baby hamster kidney (BHK)-21 cells from the American Type Culture Collection (ATCC) were used in this study, and both cell lines were maintained in our laboratory. C6/36 cells were cultured in 90% RMPI 1640 medium (Invitrogen, Carlsbad, CA) supplemented with 10% inactive fetal bovine serum (FBS; Invitrogen) and 100 U/mL of penicillin and streptomycin. BHK-21 cells were cultured in 93% Dulbecco's modified Eagle's medium supplemented with 7% FBS and 100 U/mL of penicillin and streptomycin. C6/36 cells were cultured at 28°C and BHK-21 cells were propagated at 37°C under 5% CO2 atmosphere (Wang et al. 2011).
Virus isolation
The mosquito pools (50 mosquitoes/pool) were triturated in minimum essential medium (containing 1% FBS, 100 U/mL of penicillin, 100 U/mL of streptomycin, and 1 μL/mL of fungizone) using the TissueLyser (Qiagen, Valencia, CA) at room temperature for 3 min. The homogenates were clarified by centrifugation at 12,000 rpm and 4°C for 10 min. Clarified homogenates (100 μL) were inoculated onto monolayers of BHK-21 and C6/36 cells for 1 h at 37°C and 28°C, respectively. Then, the cells were washed with minimum essential medium and were maintained at 37°C and 28°C after adding the minimum essential medium containing 5% FBS, 100 U/mL of penicillin, 100 μg/mL of streptomycin, and 1 μL/mL of fungizone. Cells were observed daily for cytopathic effects (CPEs) from days 1–7 postinfection (Li et al. 2010).
Molecular identification
Viral RNA was extracted from the cell culture supernatants using the QIAamp Viral RNA Mini Kit (Qiagen, Valencia, CA), and first-strand complementary DNA (cDNA) was synthesized using Ready-to-Go You Prime First Strand Beads (GE Health care, Uppsala, Sweden) according to the manufacturer's protocol. Polymerase chain reaction (PCR) was conducted with primers (Table 1) designed for cDNA amplification of bunyaviruses, flaviviruses, and alphaviruses in addition to JEV, TIBOV, and AKAV (Kuno 1998, Feng et al. 2016). We tested CPE-positive cultures using genus-specific primers first and then proceeding to species-specific amplification. Primers designed to amplify the whole genome of AKAV were used to sequence the virus isolates (Table 2). Amplified DNA fragments were visualized on 1% agarose gels.
M: C/A; W: A/T; Y: C/T; K: G/T; R: G/A; V: G/A/C; D: T/A/G.
PrM, premembrane; E, envelope; NS, nonstructural; RdRp, RNA-dependent RNA polymerase.
AKAV, Akabane Virus.
The positive DNA fragments were extracted (Li et al. 2010) using the TaKaRa DNA Purification Kit (TaKaRa, Japan). The PCR fragments were sequenced using the BigDye Terminator V3.1 cycle sequencing kit (PE Applied Biosystems, Foster City, CA) and analyzed using an ABI 3730 XL DNA Analyzer (PE Applied Biosystems).
Sequences of cDNA amplicons were assembled using the SeqMan program (DNASTAR;
Results
Distribution of mosquitoes
In July–August 2010 and August 2011, we collected a total of 16,076 mosquitoes consisting of seven species from the Culex, Anopheles, Armigeres, and Aedes genera, which included C. quinquefasciatus, Culex pipiens pallens, C. tritaeniorhynchus, A. sinensis, Armigeres obturbans, and A. albopictus (Table 3). C. quinquefasciatus was the most common species, accounting for 50.63% (8140/16,076) of the total. A. sinensis (24.26%, 3900/16,076) was the second most abundant species, followed by C. tritaeniorhynchus (9.76%, 1569/16,076). The proportions of C. pipiens pallens, A. subalbatus, and Culex modestus were 6.7% (1078/16,076), 5.2% (843/16,076), and 3.31% (532/16,076), respectively. Fourteen A. albopictus mosquitoes were collected.
Isolation and preliminary identification of viruses
The 16,076 mosquitoes were sorted into 347 pools according to species, location, and date of collection. Pools contained ∼50 mosquitoes each, and all the pools were processed for virus isolation using BHK-21 and C6/36 cells. Six isolates causing CPEs were obtained (Table 4). Isolates HN10169 and HN10174 caused CPE on day 2 in BHK-21 cells, but not in C6/36 cells (Fig. 2). Isolates HN10064 and HN11084 caused CPE on day 4 in BHK-21 cells and on day 3 in C6/36 cells. Isolates HN11066 and HN11121 caused CPEs on day 3 in BHK-21 cells, but not in C6/36 cells. Each virus isolate was detected by PCR using a variety of genus-specific and species-specific primers. The PCR tests showed that the six isolates were AKAV, JEV, and TIBOV.

Cytopathic effect of HN10169 and HN10174 isolates on BHK-21 cells (200× magnification).
JEV, Japanese encephalitis virus; TIBOV, Tibet orbivirus; BHK, baby hamster kidney.
Molecular biological characteristics of AKAV isolates
The HN10169 and HN10174 isolates were initially identified as AKAV, and the nucleotide sequences of the S, M, and L segments of both isolates were obtained using AKAV-specific primers (Table 2). Nucleotide sequence analysis showed that the S and M fragments of HN10169 and HN10174 isolates were 702 and 4206 bp in length, and encoded 234 and 1402 amino acids, respectively. The nucleotide/amino acid sequence homologies of the S segment of the HN10169 and HN10174 isolates were 99.1% and 99.4%, respectively, while the nucleotide/amino acid sequence homologies of the M segment of both isolates were 99.9% and 99.8%. The nucleotide sequence of the L segment of both HN10174 and HN10169 was 6756 bp, which encoded 2252 amino acids.
Comparison of nucleotide and amino acid homologies of the open reading frame sequences of AKAVs isolated from various countries, collection dates, and vectors (hosts) showed that the S segment of AKAV isolated from China was closely related to that in isolates from Japan and South Korea, (95.9–97.2% and 98.3–99.1%, respectively). The AKAV M genes in isolates from China and Japan in recent years (1993 and 2006) showed higher homology (96.7–98.1%) than that found in earlier isolates (93–96.9%) from China and Japan (1959 and 1974). The nucleotide and amino acid sequences of the S and M segments of AKAVs isolated from Australia and Kenya ranged from 70.1% to 92.9% and from 74.2% to 97.9%, respectively (Table 5).
Comparison of the deduced amino acid sequences of the S and M segments from the two AKAV isolates in this study with those of AKAVs in GenBank showed that there were several unique differences in the M and S segments between the two AKAV isolates in this study and other AKAV isolates (Supplementary Table S1; Supplementary Data are available online at
We constructed a phylogenetic tree based on the open reading frame sequences of S, M, and L segments of AKAVs isolated from various countries, host animals, and vectors. The cladogram showed that the sequence of S, M, and L segments of HN10169 and HN10174 isolates generated highly comparable topologies, indicating that these isolates have a high level of sequence homology with AKAV (Fig. 3A–C). Phylogenetic analysis of the S segment of AKAV showed that Chinese AKAV isolates formed a separate clade, while AKAV isolates from Japan, South Korea, and Australia were grouped in a different clade (Fig. 3A). The phylogenetic tree of the M segment revealed that the AKAV isolates in this study were in the same group as the AKAV strains isolated from cattle, midge, and mosquitoes in Japan (Fig. 3B).

Phylogenetic analysis of AKAV isolates based on predicted complete protein coding sequences. This analysis applied the neighbor-joining method using MEGA 7.0 software. Bootstrap probabilities for each node were calculated using 1000 replicates. Scale bars indicate the number of nucleotide substitutions per site. HN10169 and HN10174 strains are indicated with black dots. (A) Phylogenetic tree constructed using the S segment coding region nucleotide sequences of HN10169 and HN10174 strains. (B) Phylogenetic tree constructed using the M segment coding region nucleotide sequences of HN10169 and HN10174 strains. (C) Phylogenetic tree constructed using the L segment coding region nucleotide sequences of HN10169 and HN10174 strains. AKAV, Akabane virus.

Phylogenetic classification of TIBOV isolates
We conducted PCR using specific primers to amplify the 10th segment gene of TIBOV from the HN11066 and HN11121 isolates and constructed a phylogenetic tree based on the sequences of the PCR products. The cladogram showed that both HN11066 and HN11121 isolates were grouped into the TIBOV clade and were closely related to TIBOV (XZ0906 strain) (Fig. 4).

Phylogenetic analysis of TIBOV isolates based on the 10th segment coding region nucleotide sequence. This analysis applied the neighbor-joining method using MEGA 7.0 software. Bootstrap probabilities for each node were calculated using 1000 replicates. Scale bars indicate the number of nucleotide substitutions per site. HN11066 and HN11121 strains are indicated by black dots. TIBOV, Tibet orbivirus.
Phylogenetic classification of JEV isolates
We also conducted PCR on the HN10064 and HN11084 isolates using JEV E gene-specific primers and constructed a phylogenetic tree based on the PCR product sequences. The cladogram showed that both HN10064 and HN11084 isolates were grouped with JEV genotype I (Fig. 5).
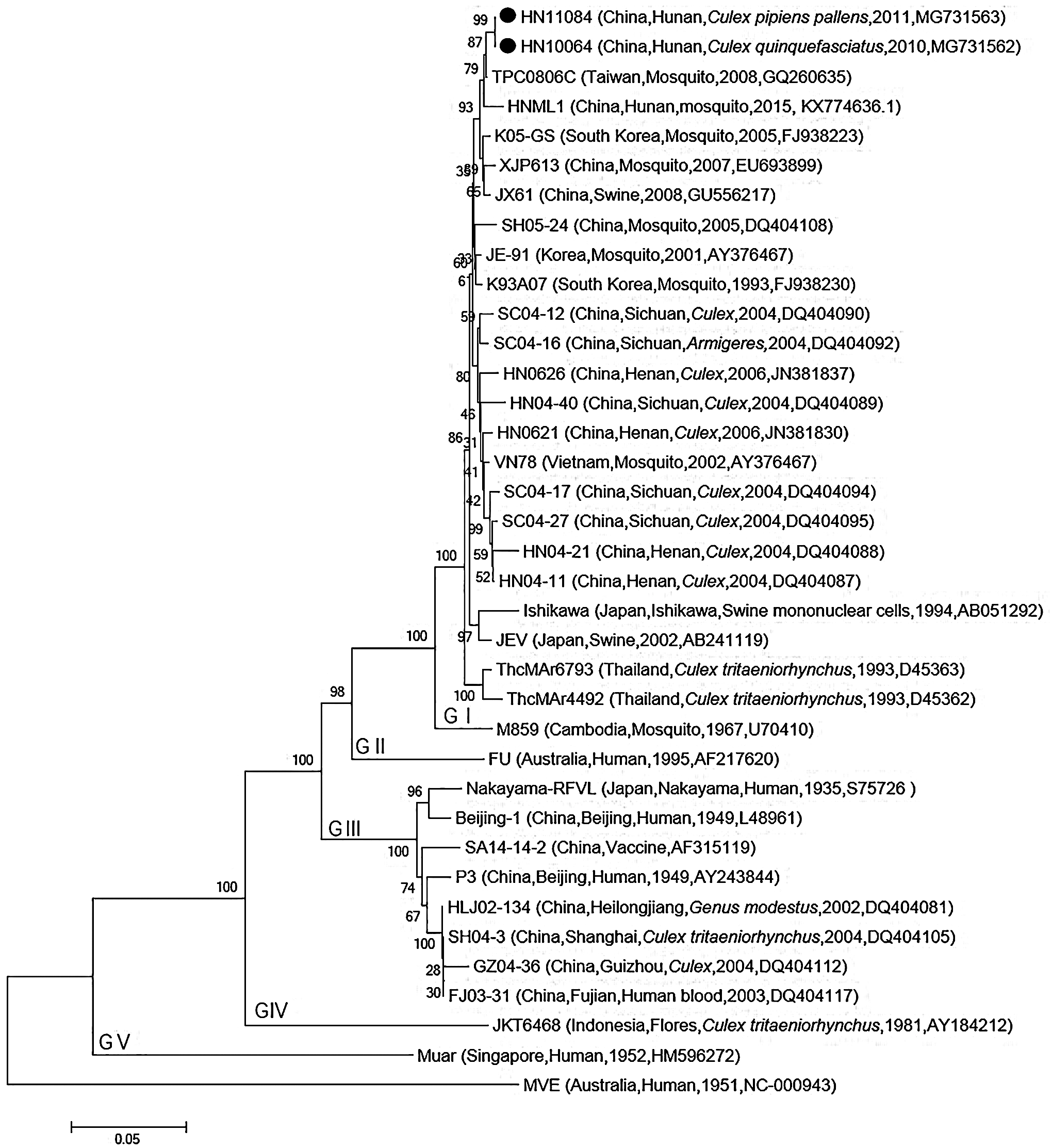
Phylogenetic analysis of Japanese encephalitis virus isolates based on nucleotide sequences of the E gene coding region. This analysis applied the neighbor-joining method using MEGA 7.0 software. Bootstrap probabilities for each node were calculated using 1000 replicates. Scale bars indicate the number of nucleotide substitutions per site. HN10064 and HN11084 isolates are indicated by black dots.
Discussion
AKAV belongs to the genus Orthobunyavirus of Bunyaviridae. AKAV was first isolated from Aedes vexans in Akabane of Gumma, Japan, in 1959 (Karabatsos 1985). It was subsequently isolated from vector specimens (biting midges and other species of mosquito) and sick animal specimens in Australia, Kenya, Israel, Saudi Arabia, Korea, and Taiwan Province of China (Metselaar and Robin 1976, Coverdale et al. 1978, Liao et al. 1996, AI-Afaleq et al. 1998, Strm et al. 2004, An et al. 2010). Recent studies have shown that two AKAV strains were isolated from C. tritaeniorhynchus (DHL10M117 virus strain) and Anopheles vagas (DHL10M110 virus strain) in Yunnan, China (Feng et al. 2016). In this study, the AKAVs (HN10169 and HN10174 isolates) were isolated from A. sinensis and C. quinquefasciatus in Hunan Province, further suggesting that AKAV can be carried by various mosquito vectors. C. tritaeniorhynchus, C. quinquefasciatus, and A. sinensis are the dominant mosquitoes in the local areas, and there is a wide range of distribution of these mosquitoes in China, spanning from the southern part of China (south of 30°N) (Liu et al. 2011 and Zheng et al. 2012). Therefore, AKAV is likely to exist in other parts of southern China as well.
Previous studies reported that the seroprevalence of AKAV neutralizing antibody in cattle and sheep was 19.06% (85/446) in Xinjiang, and the antibody seroprevalence rates of cattle and sheep were 20.32% (38/187) and 18.15% (47/259), respectively (Jun et al. 2012). The results suggested that the circulation of AKAV in mosquito vectors in China not only exists but also has caused the infection of domesticated animals such as cattle and sheep. Up to now, no studies have reported an AKAV-caused epidemic of animal diseases in China. Therefore, it is important to strengthen the detection and monitoring of AKAV in mosquitoes, midges, and animals. In particular, it is extremely important to detect and monitor the disease caused by AKAV infection in animals.
TIBOV was first isolated from Anopheles maculatus collected in Tibet, China. TIBOV contains a double-stranded RNA genome, consisting of 10 segments of various lengths and is a new member of the genus Orbivirus (Li et al. 2014). Since the first report, TIBOV has been isolated many times from midges collected in China (Lei et al. 2015, Wang et al. 2017). TIBOV was isolated from C. quinquefasciatus and C. tritaeniorhynchus collected in Hunan Province, suggesting that Culex could also become the transmission vector of TIBOV. In serum samples of domestic animals in southern China, the positive rate of TIBOV neutralizing antibody was 26% (35/135). The infection rate was highest in cattle (44%, 22/50), followed by buffalo (20%, 12/60), and goat (4%, 1/25), suggesting that TIBOV was generally infectious in domestic animals, and the susceptibility levels in livestock are distinct in different animals (Wang et al. 2017). These results suggest that TIBOV can not only be isolated from various species of mosquitoes but also infect domestic animals.
China is a JE epidemic area, and Hunan Province is a highly endemic area for JE infection in China. JE cases have been reported in Hunan Province annually since 1950 (Li et al. 2014). In recent years, the incidence rate of JE in Hunan Province has decreased year by year due to the inclusion of JE vaccine in the China Expanded Program of Immunization. In this study, mosquito collection sites were located near latrines, domestic animal pens, and sewage ditches around rural houses. JEV was isolated from two species of mosquitoes (C. pipiens pallens and C. quinquefasciatus) collected in Zhangjiajie in 2010 and Shaoyang in 2011. The results suggest that JEV was circulating in those mosquitoes in those years in Hunan. In view of the fact that C. pipiens pallens and C. quinquefasciatus can spread many species of mosquito-borne viruses such as JEV, WNV, and St. Louis encephalitis virus, it is of great significance for local public health agencies to strengthen the monitoring of the species and density of mosquitoes to detect timely mosquito-borne viruses and related diseases.
In this study, AKAV was isolated from A. sinensis and C. quinquefasciatus collected in Hunan Province, China. This finding represents not only the first isolation of AKAV from mosquitoes in Hunan Province but also the first AKAV isolation from these two mosquitoes in mainland China. AKAV has been isolated from C. tritaeniorhynchus and Anopheles vagas in China (Feng et al. 2016). These results suggest the existence of AKAV in many mosquito species in China. In Japan and Australia, AKAV has been isolated from blood samples of seemingly healthy young bulls and various tissue samples of dead fetal calves (Karabatsos 1985, Kobayashi et al. 2007). At present, although AKAV has been isolated from a variety of mosquito specimens in China, it has not yet been detected in animal specimens, and no animal infection of AKAV has been reported. Therefore, it is important to strengthen the detection and monitoring of AKAV in animal specimens of healthy cattle and sheep, and to quickly identify AKAV in sick animal specimens to reduce economic losses caused by the virus infection.
In this study, we carried out an investigation on mosquitoes and mosquito-borne viruses in 16 counties of Hunan province located in southern China, and three species of virus (AKAV, JEV, and TIBOV) belonging to 3 families were isolated from mosquito specimens. The mosquito samples were frozen in liquid nitrogen immediately after being collected, so the blood in the midgut of mosquitoes was not digested. Therefore, the viruses isolated from these mosquitoes may come from the recent engorgement of viremic blood or be carried by the mosquitoes themselves. In view of the fact that these virus isolates are all derived from CPE-positive culture cells, which we specially focused on, some viruses that cannot cause CPE in these selected culture cells may be neglected. Therefore, it is necessary to increase the application of more tissue culture cell lines to improve the chance of detecting various species of virus and to broaden the range of virus genes detection using advanced sequencing techniques such as Next-Generation Sequencing in the future study.
Footnotes
Acknowledgments
We thank the staff from the local Center for Disease Control in Hunan Province for helping to collect the mosquitoes on site. This work was supported by the National Key R&D Program of China (2016YFC1201904), the National Natural Science Foundation of China (81501757), the National Key Plan for Scientific Research and Development of China (2016YFD0500300), the Development Grants of State Key Laboratory of Infectious Disease Prevention and Control (2014SKLID103, 2015SKLID505). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statements
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.