
Abstract
Anopheles sergentii known as the “oasis vector” or the “desert malaria vector” is considered the main vector of malaria in the southern parts of Morocco. Its presence in Morocco is confirmed for the first time through sequencing of mitochondrial DNA (mDNA) cytochrome c oxidase subunit I (COI) barcodes and nuclear ribosomal DNA (rDNA) second internal transcribed spacer (ITS2) sequences and direct comparison with specimens of A. sergentii of other countries. The DNA barcodes (n = 39) obtained from A. sergentii collected in 2015 and 2016 showed more diversity with 10 haplotypes, compared with 3 haplotypes obtained from ITS2 sequences (n = 59). Moreover, the comparison using the ITS2 sequences showed closer evolutionary relationship between the Moroccan and Egyptian strains than the Iranian strain. Nevertheless, genetic differences due to geographical segregation were also observed. This study provides the first report on the sequence of rDNA-ITS2 and mtDNA COI, which could be used to better understand the biodiversity of A. sergentii.
Introduction
M
While A. labranchiae found in the northern parts of Maghreb was largely studied (Laboudi et al. 2011), data on A. sergentii populations found in the southern parts using characterization based on second internal transcribed spacer (ITS2), ribosomal DNA (rDNA), and barcoding cytochrome c oxidase subunit I (COI) are lacking. Hence, we conducted this study to fill this knowledge gap and better understand the biodiversity of A. sergentii.
For an effective investigation of the biodiversity of southern Moroccan A. sergentii population, the molecular identification remains a powerful tool that gives a detailed insight on the relationships of the vectorial capacity, genetic makeup, and geographical origin for a particular species of Anopheles, more detailed and precise taxonomy, as well as evolutionary studies (Kaura et al. 2010, Kamali et al. 2012).
Besides the morphological identification, which is no longer the only reliable method to differentiate between the species and the subspecies, the molecular tools have become indispensable for the identification of cryptic species (Collins and Paskewit 1996, Xu and Qu 1997, Proft et al. 1999). Moreover, the obtained sequences from these analyses are phylogenetically informative. For the mosquito species discrimination and phylogenetic studies, molecular markers, such as rDNA second ITS2, second and third domain (D2, D3) of rDNA 28S gene, and COI and COII of mitochondrial DNA, are largely used (Krzywinski et al. 2001, Sallum et al. 2002, Chen et al. 2003, Garros et al. 2005, Kampen 2005, Ma and Xu 2005, Marrelli et al. 2005).
Since it has been proven and recommended to combine ribosomal genes with protein-coding genes whenever possible (Danforth et al. 2005), this study was conducted to characterize and combine the ITS2 rDNA sequences and the mtDNA COI sequences to take advantage of the features of both types of data. In addition, to investigate the genetic variations among A. sergentii mosquitoes from different countries and the phylogenetics of closely related Anopheles species present in Morocco.
In this study, we used the sequence of the rDNA ITS2 and mtDNA COI as an identification tool that proved efficient for many different Anopheles species (Beebe and Saul 1995, Cohuet et al. 2003). Using universal primers and high fidelity DNA polymerase, we amplified and sequenced the ITS2 and COI regions from a subset of field larval or adult collected mosquitoes that were carefully determined by classical systematic tools such as the morphological identification using software programs. The ITS2 and COI sequences were compared, clustered to detect different variants at the population level, and also compared with the sequences available at GenBank belonging to A. sergentii population of diverse countries.
Materials and Methods
Study site
This study was conducted in the city of Marrakesh (327 km) southwest of the Moroccan capital of Rabat, in Douar Ait Daoud, Agafay, Oued N'fis (31°29′58.9″N-8°10′17.1″W). The area was characterized by a predominant hot semi-arid climate, from April to September 2015 and May to October 2016 (Fig. 1).
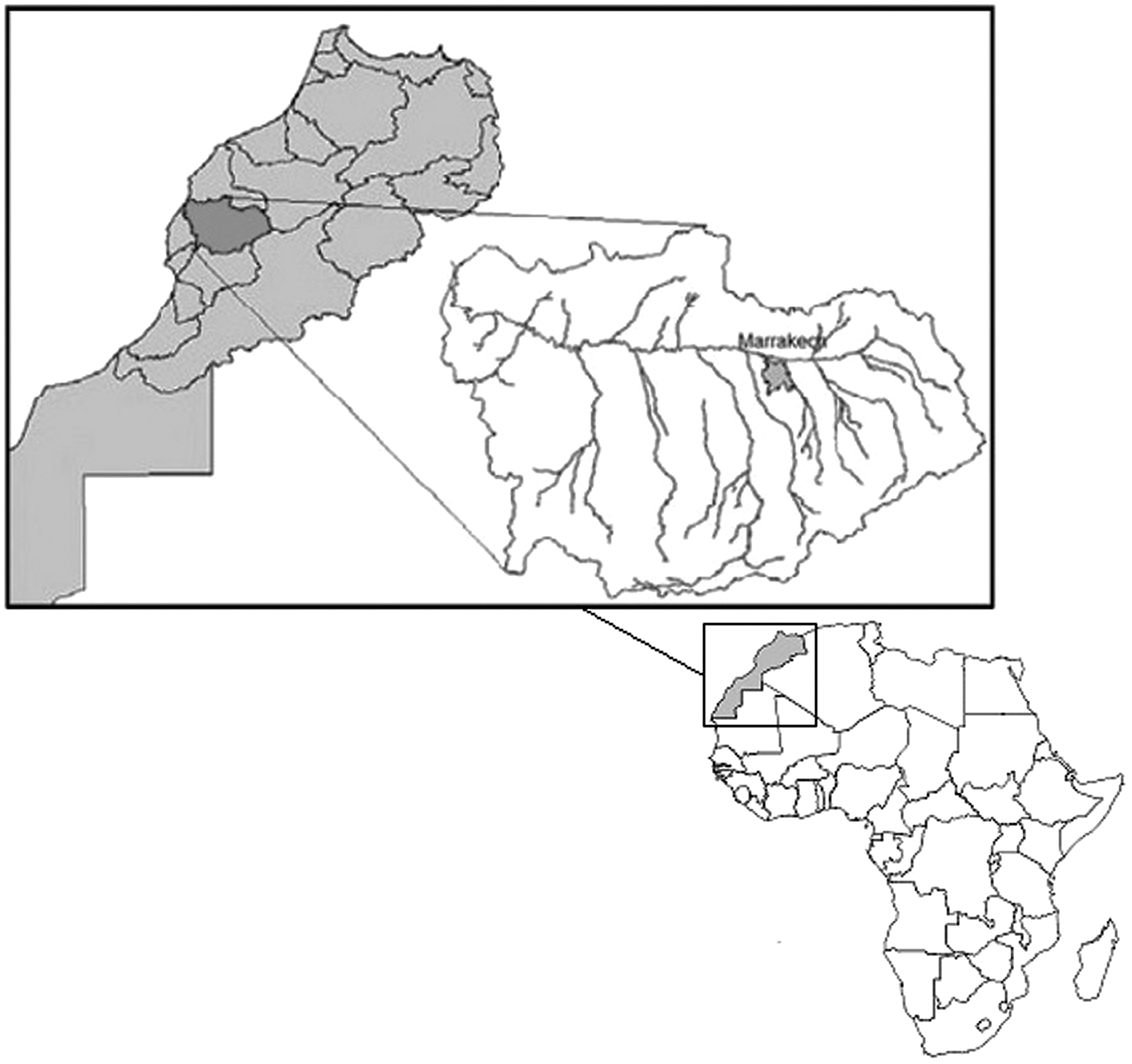
Map of Morocco indicating collection sites, where Anopheles sergentii were collected.
Mosquito sampling and identification
Four randomly selected buildings were assayed for resting mosquitoes. The collections were conducted in the morning from human dwellings and animal shelters using pyrethrum spray catches. The methods involved covering the floor with a white sheet, spraying outside and inside the buildings, and collecting the mosquitoes that were knocked down after 10 mins (Service 1993).
Larvae of A. sergentii were collected from their breeding sites using a dipper and transported to the insectary at the Institut Pasteur du Maroc, Casablanca, Morocco, and maintained at 25–28°C, ≈70–80% relative humidity with a photoperiod of 16:8 h (L:D) and reared to adults. Mosquitoes were identified using the dichotomous key for the identification of the Culicidae in the Mediterranean area to distinguish A. sergentii from other Anopheles species and stored at −20°C to carry out the molecular analyses (Brunhes et al. 2000).
DNA extraction and polymerase chain reaction amplification of the ITS2 and COI genes
DNA was isolated from individual adult mosquitoes after homogenization of the sample using DNAzol® (Genomic DNA Isolation Reagent) protocol in accordance with the manufacturer's instruction (Invitrogen, Thermo Fisher Scientific, Waltham, MA).
The ITS2 region of the rDNA was entirely amplified by polymerase chain reaction (PCR) using primers described by Porter and Collins (1991); Wesson et al. (1992); Collins and Paskewitz (1996), and Proft et al. (1999).
The COI barcode region was amplified using the primer pair LCO1490 (5′-GGTCAACAAATCATAAAGATATTGG-3′) and HCO2198 (5′-TAAACTTCAGGGTGACCAAAAAATCA-3′) (Folmer et al. 1994, Laboudi et al. 2011).
DNA sequencing
The ITS2 and COI final PCR products of 456 and ≈580 bp, respectively, were purified using the ExS-Pure™ Enzymatic PCR Cleanup Kit (NimaGen BV, The Netherlands), sequenced using the BigDye Terminator v.3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA), and run on an ABI PRISM® 3130 DNA automated sequencer (Applied Biosystems, Inc., Foster City, CA).
Phylogenetic analysis and haplotype networks
Primary ITS2 and COI sequences were examined and analyzed using Chromas v.2.6.2 software (Technelysium Pty Ltd., South Brisbane QLD 4101, Australia). Basic Local Alignment Search Tool (BLAST) website at
Results
In this study, the ITS2 and COI genes of Moroccan A. sergentii were amplified, sequenced, and the identified sequences were confirmed by BLAST analysis.
BLAST searches at GenBank database revealed the presence of a few A. sergentii ITS2 and COI sequences. Two entries were from Egypt for the COI and ITS2 genes each and one from Iran for the ITS2 gene only. Table 1 shows that there were no entries from Morocco, indicating there was no previous study or sequence submission for this region in the Moroccan A. sergentii populations. On the other hand, this population showed a high homology with the Egyptian A. sergentii strain compared with the Iranian strain.
ITS2, internal transcribed spacer 2.
Analysis of COI and ITS2 nucleotide sequences
The rDNA ITS2 of 59 A. sergentii was amplified and sequenced. The obtained nucleotide length of the ITS2 region was ∼456 bp. A fragment of 625 bp was obtained after the alignment of the sequences of the COI gene determined from a set of 39 randomly chosen from the same A. sergentii analyzed for ITS2. One sequence of each haplotype (Hap) was deposited in GenBank (Table 1). Thirty-five nucleotide sequences of the studied ITS2 and COI genes each were used in this study. The COI gene had the highest nucleotide and haplotype diversity compared with the ITS2 gene (Table 2). The analysis of the ITS2 sequences showed the presence of three haplotypes with two parsimony informative polymorphic sites that will allow us to construct a phylogenetic tree showing the haplotypes. Eighteen sequences shared the Hap1 (51.430%), 12 shared the Hap 2 (34.285%), and the remaining 6 shared the Hap 3 (14.285%) (GenBank Accession No.: MF631763-MF631765) (Fig. 2A). The COI gene showed more diversity than the ITS2 gene with 14 polymorphic sites that defined 10 haplotypes, 4 singleton variable sites, and 10 parsimony informative. Seven COI haplotypes were shared by more than one individual [Hap 1 (n = 13), Hap 2 (n = 6), Hap 3 (n = 4), Hap 6 (n = 3), and Hap 5, 7, and 8 (n = 2 each)] and the remaining three haplotypes (Hap 4, 9, and 10) were represented by one sequence each (GenBank accession No.: MF498559–MF498568) (Fig. 2B). As expected, the ITS2 sequences average of Guanine–Cytosine content was higher (56.2%) compared with the Thymine–Adenine (TA) content (43.2%) (Danforth et al. 2005). In contrast with the ITS2 data, the COI sequences average of TA content was higher (66.6%) compared with the CG content (33.4%) (Lin and Danforth 2004).

Phylogenetic analyses of haplotype data of A. sergentii in Morocco.
Number of sequences used in this study to compare between the two genes and their corresponding characteristic values.
The neutrality test described by Fu (1997) in Table 2 showed some evidence of expansion of rare polymorphisms in the ITS2 gene, whereas COI gene showed an excess of high-frequency variants. However, the negative value of this neutrality test for the concatenated sequences (ITS2 and COI combined) suggested evidence for purifying selection.
Phylogenetic analysis
For the phylogenetic comparison between A. sergentii and other Anophelines, the neighbor-joining trees were constructed using the ITS2 and COI sequences available in GenBank (Table 1). The ITS2 and COI phylogenetic trees clearly indicate that the Moroccan, Egyptian, and Iranian strains represent one and the same species, as shown in Figures 3 and 4. As expected, A. sergentii was relatively closer to An. dthali Patton than it is to A. labranchiae, as these two species can show a high morphological resemblance (Brunhes et al. 2000).
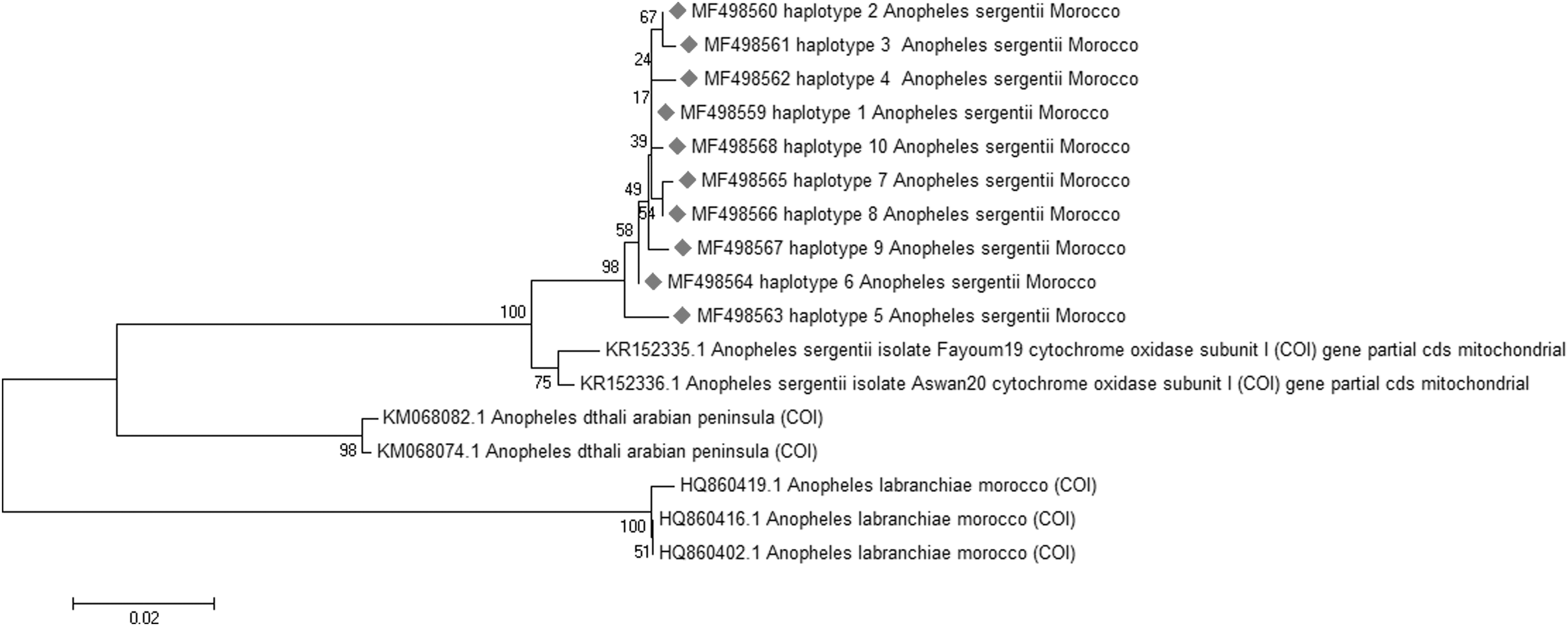
Cytochrome oxidase subunit I (COI) gene neighbor-joining tree.
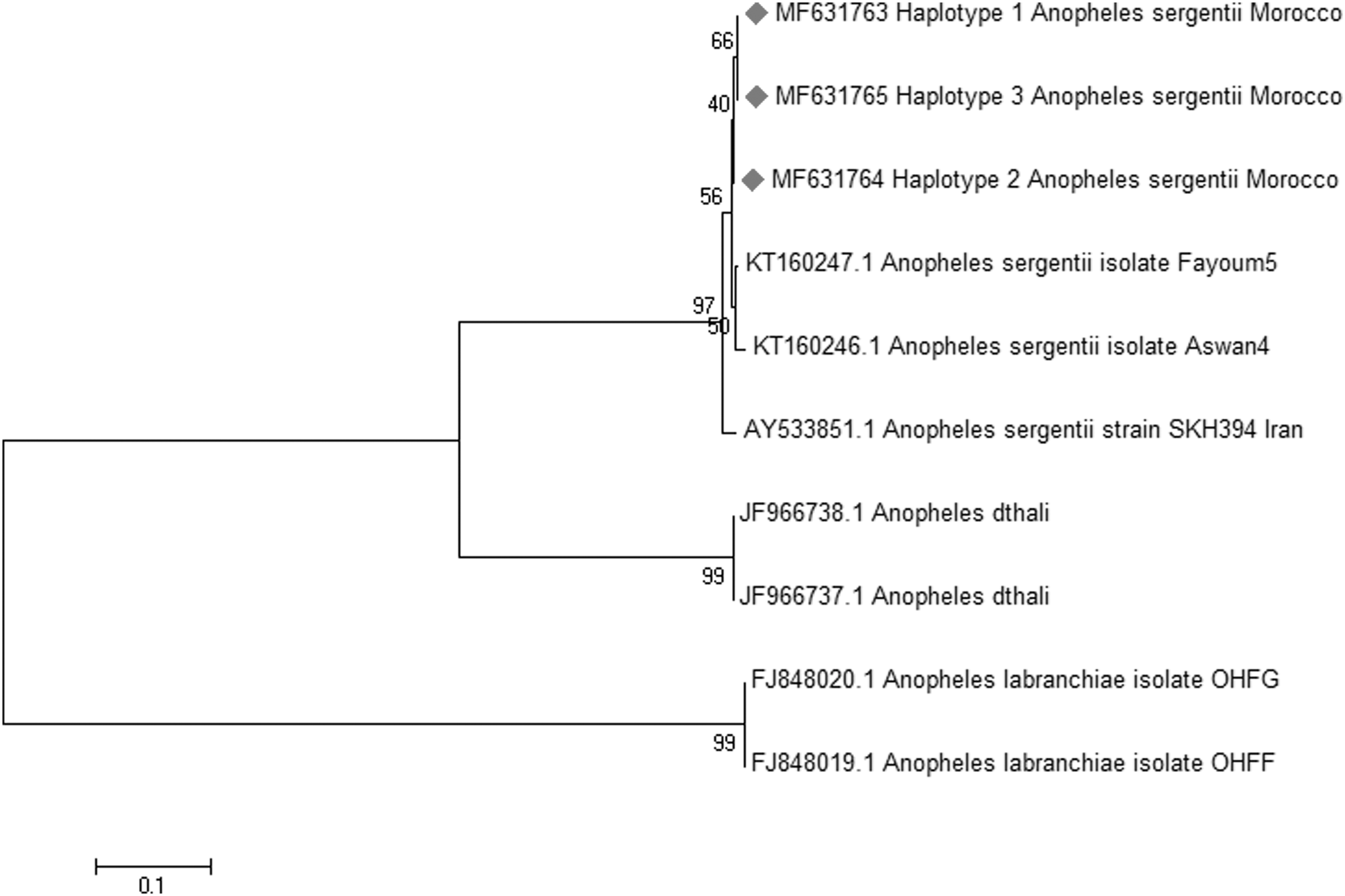
ITS2 gene neighbor-joining tree.
Discussion
In this study, we verified the presence of A. sergentii in Morocco for the first time using both ITS2 sequences and COI barcodes and provided DNA sequence data for its population for the first time to establish its genetic identity.
The mutation patterns detected on the ITS2 and COI genes of the Moroccan A. sergentii were different from those of the few Egyptian and Iranian A. sergentii sequences found in GenBank. However, based on ITS2 gene (rDNA), the analysis has shown that the North African A. sergentii (Moroccan and Egyptian) are phylogenetically closer to each other than they are to the Iranian strain, where more differences were detected. Therefore, this may suggest a high possibility of similarity between Moroccan A. sergentii and strains from the rest of the North African countries. However, further studies of other genes such as the mitochondrial genes, including more North African A. sergentii specimens (Algerian and Tunisian) are needed to verify this finding.
Correlation of the phylogenetic trees using the ITS2 rDNA and COI barcodes confirm that the different A. sergentii displayed in this study are indeed a single species. Additionally, as expected, the COI sequences showed a higher level of diversity compared with the ITS2 gene.
Other studies (Álvarez and Wendel 2003, Nolan and Cribb 2005) have reported the phylogenetic utility of the nuclear genes over the mitochondrial genes. Regarding the slow evolution rate of the nuclear genes compared with the mitochondrial genes, and the fact that the latter genes show higher levels of homoplasy (Lin and Danforth 2004) makes it less reliable in the phylogenetic study of different species. This supports our finding and the generated phylogenetic tree based on the ITS2 gene that was more accurate in the classification of the species, and thus support that ITS2 is favored on the COI gene at this point (Karimian et al. 2014). On the other hand, the COI gene gave a higher resolution tree with regard to differentiating between the A. sergentii of different countries. Using this gene, we could differentiate between the Moroccan and the Egyptian A. sergentii (the only available one in GenBank) with similar results reported by the work of Moreno et al. (2013). Therefore, the COI gene can be seen in similar cases as a better alternative to ITS2 gene (Danforth et al. 2005).
Conclusions
This study showed that the results of the COI and the ITS2 gene analysis complemented each other and both explained the presence of a geographical variation in the genotype of A. sergentii, a finding that cannot be neglected. Also, there was no clear indication of which gene was more phylogenetically informative since it was not firmly determined whether protein coding genes or ribosomal genes consistently gave the stronger results (Danforth et al. 2005). Such studies not only increase our understanding of Anopheles taxonomy, but also provide a platform for investigating different epidemiological phenomena, such as vector prevalence and species divergence.
Footnotes
Acknowledgment
This work was supported by Actions Concertees Internationales Pasteuriennes (ACIP) project no. A-02-2014.
Author Disclosure Statement
No competing financial interests exist.