
Abstract
PCR amplification and sequencing of Trypanosoma cruzi (T. cruzi) spliced-leader intergenic region of the mini-exon gene intergenic region (SL-IR) fragment was performed on intestinal tissue and fecal content DNA extracted from 19 Meccus pallidipennis (M. pallidipennis) specimens collected in the southern region of the State of Mexico. DNA sequence analysis from 49 bp T. cruzi SL-IR showed that all 19 samples corresponded to haplotype TcIa, and all of them were identical to GenBank sequence JQ028863. When extending the analysis to the whole 256 bp amplified sequence of the SL-IR, we found six sequences with a C insertion at position 10, one of which also presented a mutation (T/C) at position 54. One more sequence had an insertion (T) at position 223. Our findings suggest that two dominating TcIa clones are present in M. pallidipennis in the southern region of the State of Mexico. Interestingly, the SL-IR region of the dominating genotype was 100% identical to a circulating clone from Costa Rica present in humans, dogs, Triatoma dimidiata, and Panstrongylus rufotuberculatus. Future regional studies should explore the presence of this haplotype in humans and domestic animals.
Introduction
C
Materials and Methods
Geographical area
The present study was conducted in human settlements of the southern region of the State of Mexico (Fig. 1). This area was chosen based on previous studies where infected triatomines had been detected. These communities included the municipalities of Santo Tomás de los Plátanos, Zacazonapan, Valle de Bravo, Otzoloapan (from the Valle de Bravo Sanitary Jurisdiction), Malinalco, Sultepec, Zumpahuacán, Tonatico, Ocuilan (from the Tenancingo Sanitary Jurisdiction), and Luvianos (from the Tejupilco Sanitary Jurisdiction) (Instituto de Salud del Estado de México, 2018).
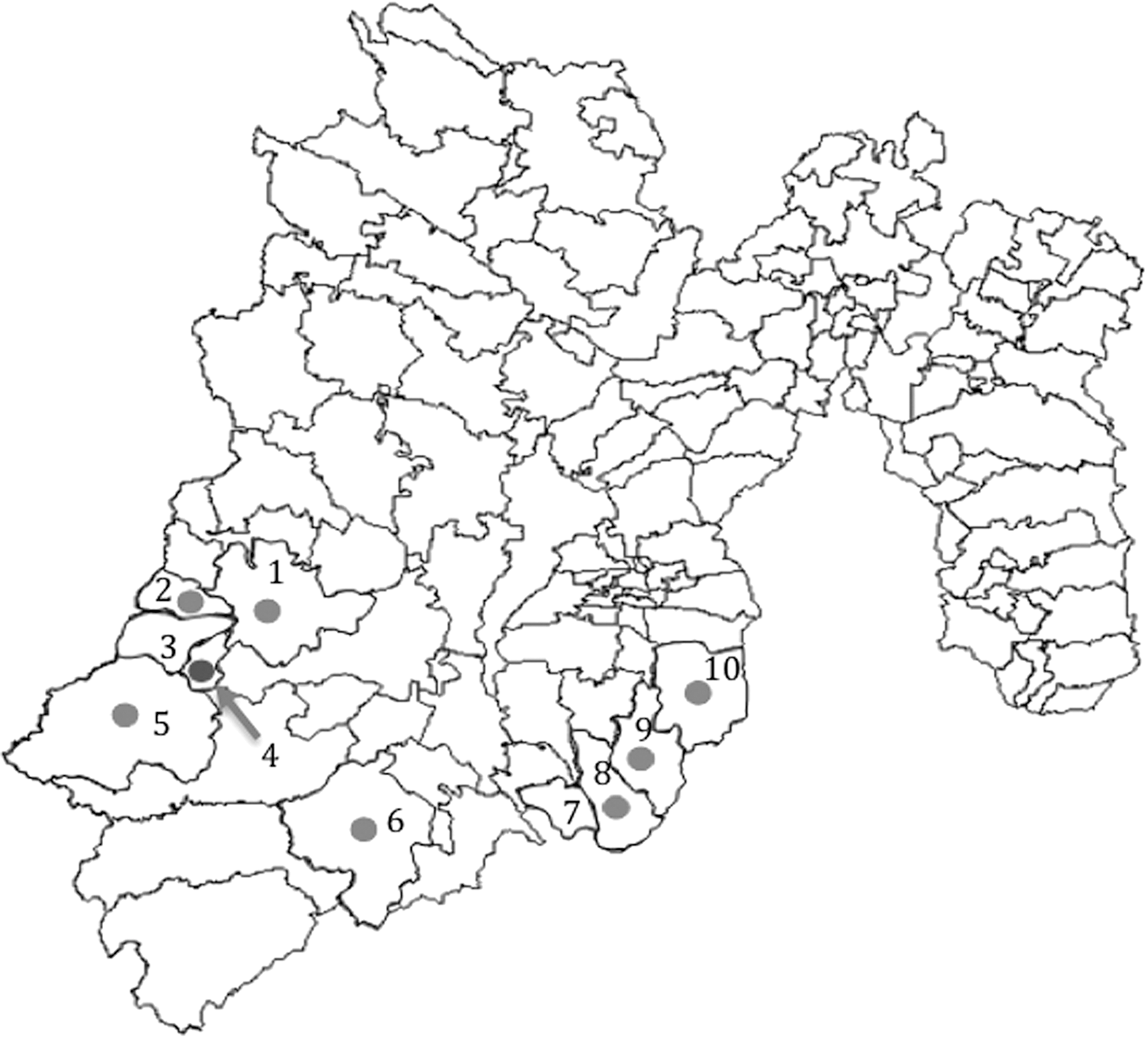
Municipalities of the State of Mexico studied for the presence Trypanosoma cruzi-infected triatomines: 1, Valle de Bravo; 2, Santo Tomás de los Plátanos; 3, Otzoloapan; 4, Zacazonapan; 5, Luvianos; 6, Sultepec; 7, Tonatico; 8, Zumpahuacán; 9, Malinalco; 10, Ocuilan. Municipalities 1, 2, 3, and 4 belong to the Valle de Bravo Sanitary Jurisdiction. Municipality 5 belongs to the Tejupilco Jurisdiction. Municipality 6 belongs to the Tejupilco Sanitary Jurisdiction, and municipalities 7, 8, 9, and 10 belong to the Tenancingo Sanitary Jurisdiction. Triatomines were collected in all municipalities marked with a number. Marked in light gray dots are municipalities where T. cruzi-infected triatomines were found. Marked in dark gray dot is municipality where Triatoma dimidiata was found.
Vector sampling
Triatomines were collected during the rainy season (May–October 2015). Sampled houses were chosen according to previous reports of triatomine presence (Medina-Torres et al. 2010) and consisted mainly of rustic thatched and/or wood roofing. Walls were made of perishable materials, such as adobe, bricks without plastering, wood, cane, and straw, and most houses had dirt floors. Most of these building materials represent risk factors associated with the presence of triatomines. Other risk factors commonly associated with parasite transmission to humans present in these houses were indoors and/or outdoors clutter and presence of dogs and/or cats within the dwellings. In the peridomicile area, families frequently kept chicken coops and/or farm animal pens in close contact (∼12 meters) with the living area. Triatomine searching was performed according to the Mexican Official Standard for epidemiological surveillance, prevention, and control of vector-transmitted diseases (NOM 2003). Triatomines collected within the domicile were found by inspecting the house clockwise, searching under clothes or objects that could provide shelter to triatomines, and also in grooves in walls and roofs or similar spaces where the kissing bugs could find protection. Peridomicile triatomines were mainly found in dog shelters, chicken coops, pig pens, barns, and among piled rocks and/or wood.
Insect handling
Triatomines were collected in containers that were labeled with the name and address of the householder where the insect was found. Identifiers included the site of collection (municipality and community names, community section, and street name and number). In the laboratory, triatomines were observed under stereoscopic microscope and taxonomically classified considering the morphological characteristics (Lent and Wygodzinsky 1979).
Parasite DNA extraction and purification
DNA was extracted and purified from fresh triatomine fecal samples collected by abdominal pressure. When it was not possible to obtain fecal samples from any specimen, the DNA from the whole intestine of the insect was extracted. Extraction and purification of DNA was performed with a commercial kit (ZR Tissue & Insect DNA MicroPrep™; Irvine, CA) following the manufacturer's instructions. DNA quantitation was performed with a Q5000 UV-Vis spectrophotometer (Quawell, San Jose, CA). Purified DNA samples were stored at −20°C until processing.
T. cruzi DNA isolation from triatomines
T. cruzi SL-IR DNA fragment is a molecular marker used to differentiate the TcI DTU from other major DTUs of T. cruzi (TcII to TcVI). Here, we performed an initial PCR screening to identify infected triatomines and, at the same time, to find out if DTUs other than TcI were present. The DNA fragment from T. cruzi SL-IR was amplified from the DNA extracted from fecal or intestinal samples from triatomines using a pool of the primers designed by Souto et al. (1996): Forward TcI specific, 5′GTGTCCGCCATCCTTCGGGCC3′ (TcI); forward common to TcII–TcVI, 5′CCTGCAGGCACACGTGTGTGTG3′ (TcII); and reverse common for TcI–TcVI 5′CCCCCCTCCCAGGCCACACTG3′ (Tc). Expected amplicons size was 350 bp for TcI and 300 bp for TcII–TcVI (Souto et al. 1996, Fernandes et al. 1998). PCR was performed using the Mastermix kit from Qiagen (Santa Clarita, CA) following the manufacturer's instructions. Briefly, 2.5 Taq DNA polymerase units, 200 μM DNTPs, 1.5 mM MgCl2, 0.2 μM of each primer, and 60–100 ng of purified DNA were mixed with PCR buffer and DNAse-free milliQ H2O in a total reaction volume of 25 μL. PCR conditions were as follows: initial denaturation at 95°C for 5 min, followed by 30 cycles of denaturation (95°C for 30 s), annealing (61.6°C for 30 s), and extension (72°C for 30 s); the cycle ended with a final extension incubation at 72°C for 5 min and a cooling-down incubation at 4°C for 5 min. Products were resolved by 3% agarose gel electrophoresis and stained with ethidium bromide.
Amplicon sequencing
Products were purified using the Wizard® SV Gel and PCR Clean-Up System from Promega (Madison, WI) following the manufacturer's instructions (Santana et al. 2014) and sent for sequencing to Macrogen, Inc. (Seoul, Korea). Forward and reverse electropherograms from each amplicon were edited with Codon Code Aligner (CodonCode Corporation,
DTU determination
A 350 bp amplification fragment was expected for all TcI DTU samples and a 300 bp amplification band for DTUs TcII to TcVI. To further analyze PCR amplification products, they were sent for sequencing and then compared for sequence match against reference DTUs sequences retrieved from GenBank (number in bold): TcI (BOL,
Haplotype analysis
Since no DTU other than TcI was found through PCR amplicon electrophoretic band size nor when comparing our samples against DTU reference sequences, we analyzed the SL-IR to identify the haplotypes present in local sequences and comparing them with the following haplotype sequences retrieved from GenBank (number in bold): TcIa (USA28,
Phylogenetic inferences
Phylogenetic and molecular evolutionary analyses were conducted using Mega version 6 (Edgar 2004, Tamura et al. 2013). All sequences were adjusted to 256 bp, the maximum length sequence considered for analysis was adjusted to the maximum length where all our electropherograms presented a clear reading. Phylogenetic analysis was carried out by building a maximum-likelihood tree set for a phylogeny test as follows: Analysis: Phylogeny reconstruction; Statistical method: Maximum likelihood; Phylogeny Test: Bootstrap method with 1500 bootstrap replications; Substitution model: Tamura–Nei model with nucleotide substitution; Rates and Patterns: Gamma Distributed (G) with five discrete gamma categories; Data subset: Gaps (complete deletion); Tree Inference: ML Heuristic Method (Nearest-Neighbor-Interchange); Branch swap filter: very strong with one thread.
Results
Triatomine population composition
Sixty-three triatomines were collected: 25 (39.6%) were classified as adults and 38 (60.3%) as nymphs. Forty-seven specimens (74.6%) were collected from the peridomicile, 11 (17.5%) from the intradomicile, and 5 (7.9%) were not recorded appropriately (data not shown). Triatomines found in the region were mainly Meccus pallidipennis (M. pallidipennis) (n = 46; 73.01%). However, one specimen of Triatoma dimidiata (T. dimidiata) (n = 1; 1.58%) was also found. Sixteen first- and second-instar nymphs (25.4%) could not be taxonomically classified. Considering the composition of the whole collection, it is very likely that most of these nymphs were M. pallidipennis. However, no further identification tests were conducted. Interestingly, this was the first record of T. dimidiata from the municipality of Zacazonapan.
Molecular identification of T. cruzi in triatomines
Nineteen T. cruzi-infected triatomines (N = 63, 30.1%) were detected through PCR analysis. Seventeen were classified as M. pallidipennis and two remained unclassified. Eleven were adults and eight were nymphs. Infected triatomines were found in 7 of 10 communities (Table 1). All 19 samples showed a 350 bp PCR diagnostic fragment in agarose gel electrophoresis (Fig. 2), corresponding to TcI SL-IR DTU. Amplicons were purified and sent for sequencing for further phylogenetic analysis.

Amplified DNA samples from Meccus pallidipennis. MW, molecular weight 1000 bp; 1, TcI positive control; 2, negative control; 3–16, samples. Black arrow shows a 350 bp molecular weight.
Infected Triatomines Collected in Different Municipalities of the State of Mexico: Classification by Species, Site of Collection, and Developmental Stage
Triatomines were collected at indicated municipalities and classified by municipality, collection site, developmental stage, and species. DNA from fecal samples or intestinal content was purified, and SL-IR mini-exon region was amplified by PCR to detect infected triatomines. Sample number shown corresponds to specimens diagnosed as infected with Trypanosoma cruzi. W/I, without information.
DTU and haplotype determination
All PCR amplicons from T. cruzi SL-IR mini-exon gene amplified from DNA extracted from fecal samples or triatomine intestine tissue showed a 350 bp band in agarose electrophoresis, indicating that all the samples corresponded to TcI DTU (Fig. 2). For further DTU analysis, the 19 local sequences were compared with the following DTU sequences retrieved from GenBank (number in bold): TcI (BOL,
All 19 sequences successfully aligned with BOL (previously classified as TcI) but not with any other sequences (data not shown). These were then compared against the following GenBank TcI haplotype reference sequences (n = 7): USA28 (TcIa), Mg11 (TcIb), X380C (TcIc), V195 (TcId), TALAVERDE (TcIe), Pan4 (TcIa+TcId), and WTc17 (TcIa+TcIe) (Herrera et al. 2007, Cura et al. 2010). BOL (TcIa) was included as control. All local sequences clustered with TcIa haplotype (Table 2 and Fig. 3). Interestingly, 18 of 19 samples correctly aligned with haplotype TcIa when using the 49 bp SL-IR region (Cura et al. 2010). The remaining sample, although not identical, closely aligned with Pan4 (Table 2), classified as a TcIa+TcId haplotype, originally isolated from a human sample from Panama (Cura et al. 2010).
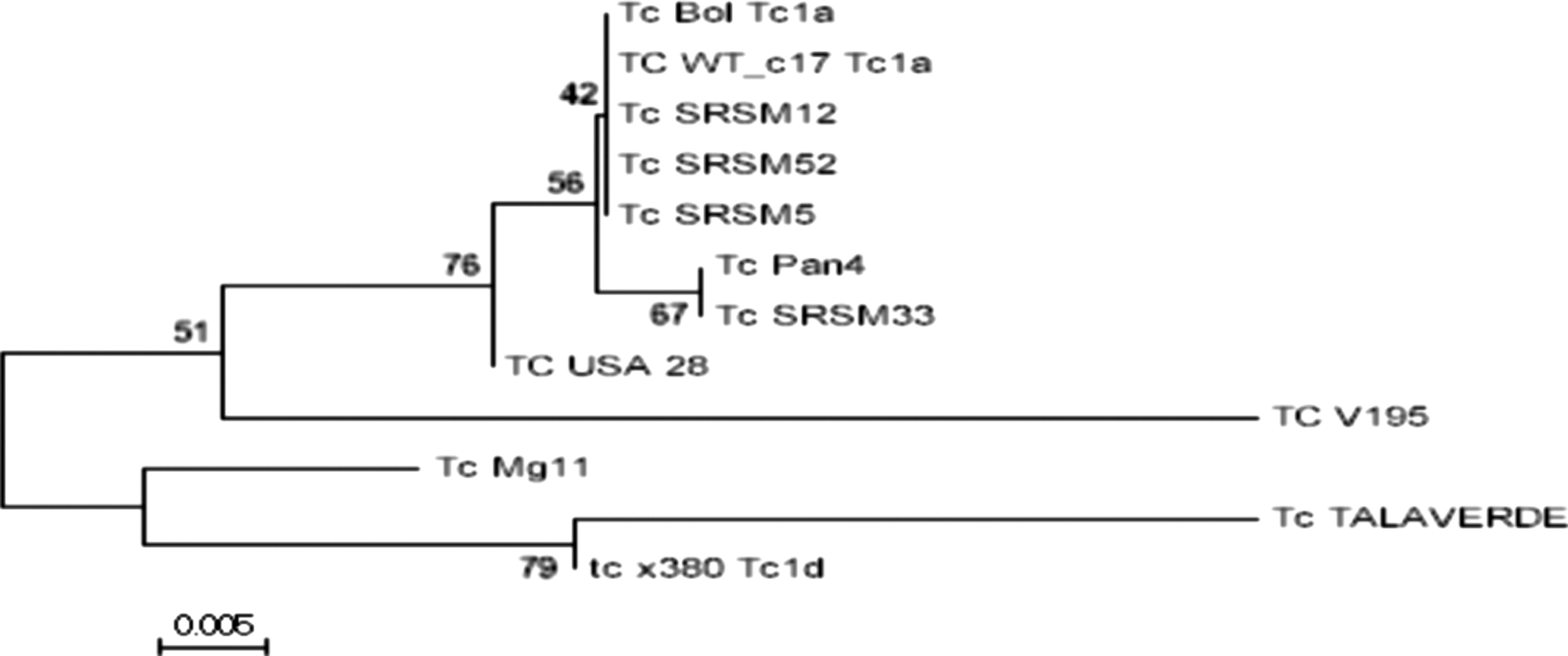
Phylogenetic tree constructed with T. cruzi SL-IR amplicons from local and TcI reference samples downloaded from GenBank: USA28|gb|GU179071.1| (TcIa); BOL|gb|JQ028863.1| (TcIa); Mg11|gb|FJ713379.1| (TcIb); X380C|gb|AM259472.1| (TcIc); V195|gb|GQ398820.2| (TcId); TALAVERDE|gb|GQ398816.2| (TcIe); Pan4|gb|GU903132.1| (TcIa+TcId); and WTc17|gb|GU903156.1| (TcIa+TcIe). Analysis was conducted using Mega version 6.06-Mac (Tamura et al. 2013) (see text for parameters used). When identical local sequence samples were joined within a single name: Tc SRSM 12 represents identical sequences found in samples 11, 14, 22, 23, 31, 86, 112, 117, 126, 127, 144, and 145; Tc SRSM 5 represents identical sequences found in samples 41, 46, 49, 114, and 142. Tc SRSM 33 and Tc SRSM 152 are sequences registered with a unique SNPs. SNPs, single-nucleotide polymorphisms.
Comparative Single-Nucleotide Polymorphisms Found in Local and Reference TcI Haplotypes Based on SL-IR Trypanosoma cruzi Sequence (Sites 10–38)
Mutations found in a 256 bp DNA fragment from T. cruzi mini-exon gene intergenic region (SL-IR), which characterizes TcI haplotypes: TcIa, USA28; TcIb, MG11; TcIc, X380C; TcId, V195; TcIe, TALAVERDE; TcIa+TcId, Pan4; TcIa+TcIe, WTc17. Identical local sequence samples were merged within a single name: Tc SRSM 12 represents identical sequences found in samples 11, 14, 22, 23, 31, 86, 112, 117, 126, 127, 144, and 145; Tc SRSM 5 represents identical sequences found in samples 41, 46, 49, 114, and 142. Tc SRSM 33 and Tc SRSM 152 were not identical to any other sequences. SL-IR region DNA sequences obtained from triatomine fecal samples or intestinal tissue were aligned with Muscle (MEGA 6; Edgar 2004). Single-nucleotide polymorphisms were numbered according to their position in the sequence and annotated in order in the table. Dot (.) represents nucleotide identity with the sequence used as reference: WTc17 (TcIa + TcIe); underscore (_) represents a missing nucleotide with respect to other sequences.
Discussion
SL-IR has been used to allocate T. cruzi into seven DTUs, namely TcI–TcVI and TcBat (Zingales et al. 2012, Lima et al. 2015), and TcI into five haplotypes (Cura et al. 2010). These genotypes seem to be associated with different epidemiological, biological, and medical features of the parasite, such as transmission cycles and disease (Zingales et al. 2012, Brenière et al. 2016). This highlights the importance of identifying the genotypes of parasites circulating in both vectors and hosts in different endemic areas. T. cruzi is considered endemic in most of the Mexican territory, where the existence of six DTUs (I–VI) has been demonstrated (Ramos-Ligonio et al. 2012, Ibáñez-Cervantes et al. 2013, López-Cancino et al. 2015, Nguyen 2016). Although T. cruzi is also considered endemic in the State of Mexico, it had not previously been molecularly characterized. The aim of the present study was to genotypically characterize T. cruzi infecting triatomines present in the domicile and peridomicile of human dwellings of the southern region of the State of Mexico, where T. cruzi-infected triatomines, dogs, and humans were previously reported (Estrada-Franco et al. 2006, Medina-Torres, 2010, Barbabosa-Pliego et al. 2011).
Sixty-three triatomines were collected in the study area (Fig. 1). Forty-six (73.01%) were M. pallidipennis, 1 (1.58%) T. dimidiata, and 16 (25.4%) were not classified as they were early nymphal stages. The proportion of species found was similar to previous reports by Medina-Torres et al. (2010), who classified 97.4% of the collected specimens as M. pallidipennis and 2.6% as T. dimidiata in the same geographical region. Interestingly, T. dimidiata is considered one of the most widespread species of triatomines in Mexico (Bautista et al. 1999, Cruz-Reyes and Pickering-López, 2006), and although the ecological conditions in the study area suggested an ample presence of this species, the scarce number of T. dimidiata found in this survey is intriguing. Future studies exploring the biogeography patterns, ecology, and feeding behavior of T. dimidiata and M. pallidipennis in the southern region of the State of Mexico and neighboring states should be conducted to better understand triatomine ecological niche selection, which might result in better vector control strategies.
Triatomines collected were mainly first- to fourth-instar nymphs (60.49%), with a lower proportion of adults (39.5%). Of all collected specimens, 74.6% were found in the peridomicile and only 17.5% were within the human dwellings. These data support previous reports by Medina-Torrres et al. (2010) of M. pallidipennis presence within the peridomicile and inside the human dwellings in the southern region of the State of Mexico.
Nineteen (30.1%) collected triatomine specimens were infected with T. cruzi as demonstrated by PCR and electrophoresis analysis, based on a 350 bp DNA amplification band from the SL-IR T. cruzi region. This proportion of T. cruzi-infected insects agrees with previous reports that registered infection rates in M. pallidipennis of ∼30% in the southern region of the State of Mexico (Estrada-Franco et al. 2006) and suggests the occurrence of a constant infection prevalence in regional triatomine vectors. Since the vectors were collected in domestic and peridomestic areas, these results suggest that triatomines still alarmingly show similar infection rates as those first reported more than 10 years ago. This should be considered a reminder to public health authorities, policy makers, researchers, medical staff, and all the actors involved in the control of vector-borne diseases about the complexity of Chagas disease. A public health issue still constitutes a silent threat to the population in the southern region of the State of Mexico.
All 19 amplified products were 350 bp long and classified as TcI DTU. To further analyze the samples, amplicons were purified and sent for sequencing. Forward and reverse electropherograms from each product were edited to correct ambiguities, compared to find a consensus sequence between them, and then aligned and compared against six GenBank reference sequences for DTUs I–VI (BOL, IGRE F544, MN, CANIII, Tu18, and M5631). Comparisons confirmed that all 19 samples belong to TcI clade (data not shown). These results suggest that M. pallidipennis from the study area are mainly infected with T. cruzi genotyped as TcI and support previous reports showing that TcI is the predominant genotype in Mexico and the main DTU infecting Meccus spp. (Bosseno et al. 2002, Espinoza-Gómez et al. 2002, Guhl and Ramirez 2011, Alkmim-Oliveira et al. 2013, Brenière et al. 2016). It has been previously reported that any of four T. cruzi DTUs (TcI–TcIV) naturally infect M. pallidipennis, either individually or simultaneously (TcI/TcIV and TcII/TcIV) (Ibañez–Cervantes et al. 2013). Although only DTU TcI was observed in the present study, the presence of other T. cruzi lineages should not be discarded until further and more extensive studies are performed, increasing sample size aiming to less abundant T. cruzi genotypes.
To identify the TcI haplotypes circulating in the region, the local sequences were aligned and a phylogenetic tree was constructed (Mega 6.06-Mac software, Tamura et al. 2013) including eight reference SL-IR sequences obtained from GenBank (number in bold), which represent the different reported TcI haplotypes: TcIa (BOL,
It is interesting to note that the clones present in M. pallidipennis in the southern region of the State of Mexico are nearly identical to those circulating in Costa Rica, which were isolated from both mammals (including humans) and triatomines. These results support the current population distribution model where lineage TcI is widespread in most T. cruzi endemic environments of the Americas (Brenière et al. 2016).
Conclusions
M. pallidipennis is the most abundant triatomine species in the southern region of the State of Mexico. Specimens collected during this study presented T. cruzi infection rates of 30.1% and were similar to those first recorded over a decade ago. DTU and haplotype determination through DNA sequencing of the PCR amplicons of the mini-exon gene (SL-IR) obtained from the DNA extracted from gut tissue and/or fecal content of triatomines suggest that TcI constitutes the predominant DTU in triatomines and that TcIa is the preponderant haplotype present in these vectors in the southern region of the State of Mexico. Further studies aiming to isolate and molecularly characterize T. cruzi from humans and other mammals should confirm this population structure.
Footnotes
Acknowledgment
This work was funded by the Universidad Autónoma del Estado de México (UAEM): project number 1042/2014.
Author Disclosure Statement
No competing financial interests exist.