
Abstract
The usefulness of a strategy involving the biological control of soil-transmitted helminths (STHs) with zoonotic potential is tested. A blend of spores of two filamentous fungi, Mucor circinelloides and Duddingtonia flagrans, was provided to one group (G-2) of puppies (five Griffon bleu de Gascogne and five Gascon Saintongeois) shedding eggs of Toxocara canis, Toxascaris leonina, Trichuris vulpis, and Ancylostoma caninum. Other identical group (G-1) remained without spores. Fecal samples were collected daily and divided into two portions, one sample was immediately analyzed by the flotation test and the other was examined 28 days later. Viability of eggs of T. canis, T. leonina, and T. vulpis in fecal pats was ascertained by microscopic examination; the numbers of larvae of A. caninum were assessed in each group. In the feces of G-2, spores of both fungi were observed 1 day after beginning the assay. Viability of eggs of T. canis was reduced by 57%, T. leonina by 61% and T. vulpis by 53% in the fecal pats of G-2, and by 87%, 92%, and 85%, respectively, in G-1. The numbers of third-stage larvae of A. caninum were 59% lower in G-2 than that in G-1. By considering the results obtained and the absence of adverse effects in the puppies of G-2, it is concluded that the presence of the filamentous fungi in the feces has a high potential to avoid the transmission of these parasites. Further studies are in progress to determine the appropriate way of administration to pets.
Introduction
S
Contamination of soil by infective eggs of ascarids and trichurids, and third-stage larvae of ancylostomids, increases the risk of their transmission between animal species, and consequently to humans. This situation tends to be addressed in pets by their regular deworming before, during and after pregnancy, but the effect in bitches can be lower than expected because the larvae remain arrested in their tissues and is then reactivated during pregnancy and lactating, causing infection in adult females and puppies (in utero) (Duprey and Schantz 2003). Frequent deworming is advised in puppies, but sometimes they become infected again several months after treatment. Pups can be affected by STHs through transplacental (Toxocara canis), percutaneous (Ancylostoma caninum), and transmammary routes (Toxocara and Ancylostoma), besides the ingestion of infective stages in contaminated ground where they can stand viable for years (Traversa et al. 2014). In accordance, bitches and progeny should be maintained in an environment (ground) free of these parasites.
It has been reported that the presence of certain soil saprophytic fungi with parasiticide activity (Pochonia chlamydosporia, Verticillium chlamydosporium, and Mucor circinelloides) is capable of destroying eggs of canine helminths such as T. canis and Dipilydium caninum and others (Duddingtonia flagrans and Monacrosporium thaumasium) can capture and eliminate the larvae released from the eggs of A. caninum (Araujo et al. 2009, Arias et al. 2013, De Mello et al. 2014).
With the aim to assess the possibility to reduce the risk of infection by some STHs, one group of puppies shedding eggs of some helminths received a blend of spores of two filamentous fungi, M. circinelloides (ovicide) and D. flagrans (larvicide). Viability of eggs of T. canis, Toxascaris leonina, and Trichuris vulpis, and variations in the counts of larvae of A. caninum were analyzed.
Materials and Methods
Dog breeding facility
The current assay was carried out in the “Soñar” commercial breeding kennel (42°57′32.689′′ N, 7°40′40.78′′ W; 632 m altitude; Lugo, NW Spain) focused on raising Griffon bleu de Gascogne and Gascon Saintongeois dogs. Adult dogs (≥1.5 years) are maintained in metal fenced pens with concrete floor, hygienized once a day by using pressurized hot water. Peri-parturient bitches are placed in individual plots for whelping. One-month-old puppies were moved to 0.5 Ha plots with dirt floor, shade, plants, shelters, drinkers, and feeders; they are fully weaned at 2 months old. Puppies and adults do not share ground plots.
Control of parasites involves the collection of feces from the ground every 6 months, which are analyzed by means of the flotation test; one single dose of 100 mg fenbendazole (Panacur® Granules 22.2%; Intervet GesmbH, Vienna, Austria)/kg bodyweight (b.w.) is administered trimonthly to the adult dogs. Pregnant bitches do not receive any special treatment. Puppies are given 5 mg pyrantel pamoate (Strongid® T; Zoetis, Spain)/kg b.w. at 2 weeks old; fenbendazole is administered at 1 month old, then bimonthly until they are 7 months old and later trimonthly (as pointed for adults).
Fungal spores
Two parasiticide fungi, M. circinelloides (CECT 20824; ovicide) and D. flagrans (CECT 20823; larvicide) were simultaneously cultured in a submerged medium (COPFr) for 1.5–2 months at room temperature.
Examination of fecal samples
Throughout an interval of 13 days (from 1 day before the start to the next day to the end of the assay with the spores), 20 fecal samples were taken daily from the ground in each pen, then divided into two portions (Fig. 1). Ten grams of each sample (portion 1) was examined the same day of collection. Five grams of each sample was evaluated for the presence of eggs of STHs by means of the flotation test. Feces were homogenized into water, passed through a 150 μm filter, and centrifuged at 2000 rpm for 5 min. The supernatant was discarded and the sediment was added to 10 mL saturated sodium chloride solution (ρ = 1.20 g/cm3) and observed in a McMaster chamber under a light microscope (Leica DM2500) at 10 × (Cazapal-Monteiro et al. 2015). The remaining 5 g was examined for the presence of fungal spores. Samples were processed as mentioned, but the sediment was added to 2 mL distilled water. Four aliquots of 50 μL were placed between a glass slide and a coverslip and observed at 40 × under a light microscope.

Experimental design of an assay to evaluate the potential usefulness of filamentous fungi to prevent zoonotic STHs. STH, soil-transmitted helminth.
The other portion of feces (portion 2, 10 g) was introduced into polypropylene translucent boxes (15 × 6 × 15 cm). A total of 12 boxes were prepared per day and group, maintained under field conditions during 28 days and then analyzed using the flotation and the Baermann tests. Eggs of ascarids or T. vulpis were observed by placing aliquots of 50 μL between a glass slide and a coverslip, and then observed at 20 × and 40 × under a light microscope until ≥150 eggs were visualized in each sample. The numbers of third-stage larvae of A. caninum were estimated by means of the Baermann probe.
The antagonistic activity of the filamentous fungi on the eggs of STHs was assessed by evaluating their viability. Those eggs without damage or with hyphae attached to the shells were sorted as viable (Cazapal-Monteiro et al. 2015). Variations in the egg viability were evaluated according to the percentage of viability reduction:
Regarding the larvae of A. caninum, reduction in the numbers was estimated by the following formula:
Experimental design
The experimental design was approved by the Ethical Committee of the University of Santiago de Compostela (Spain; protocol number CTM2015-65954) and complied with the Directive 2010/63/EU.
Twenty 3-month-old puppies were divided into groups G-1 and G-2, each composed of five Griffon bleu de Gascogne and five Gascon Saintongeois. All puppies were taken from the same plot and placed in two fenced cages; puppies had been previously dewormed by the administration of 5 mg pyrantel pamoate (Strongid T, Zoetis)/kg b.w. at 2 weeks old and then by fenbendazole at 1 month old.
Pups in G-1 received dry feed throughout the study and served as controls. During a period of 11 days, a daily dosage of 1 × 106 spores of M. circinelloides + 1 × 105 spores of D. flagrans was provided to the puppies in G-2 (Fig. 1). For this purpose, commercial dry feed was mixed with a submerged medium containing fungal spores, and an approximate amount of 100 g of this food premix was given daily to each puppy in individual feeders, taking care that every pup ingested the appropriate ration.
Climatic parameters collected from an automatic weather station located at 21 km (Corno do Boi, Friol, Lugo, NW Spain, 43°02′24′′N, 7°53′24′′W) showed values of 19°C ± 5.9°C maximum temperature, 10.5°C ± 2.6°C minimum temperature, 89.5% ± 8.6% relative humidity, 5.5 ± 9.6 L/m2 rainfall, and 2.2 ± 10.8 water balance (L/m2) during the trial.
Adverse effects
The possibility of puppies in G-2 could refuse the food premix was evaluated. Digestive activity was examined in all puppies given fungal spores (G-2) looking also for weakness, any adverse effect on appetite, increased thirst, and consistency of feces.
Respiratory function was assessed by looking at the rate of breathing, fatigue, and recovery rate after exercise, including persistent cough or nasal discharge. Finally, attention was paid to the appearance of skin damage (redness, blistering, peeling, or cracking).
Statistical analysis
According to the Kolmogorov–Smirnov test, data were not normally distributed (Z values <0.05), and the Levene's test showed the variances were not homogeneous (p < 0.05). The nonparametric probe Mann–Whitney U test was performed at a significance level of p < 0.05. All tests were carried out by using the statistical package SPSS, version 20 (IBM SPSS, Inc., Chicago, IL).
Results
Detection of parasites in feces
At the beginning of the trial, the analysis of feces collected in the two pens showed the presence of STHs only, and the average counts of eggs per gram feces ranged between 894 and 1575 T. canis, 313–518 T. leonina, 31–69 T. vulpis, and 63–156 A. caninum. No statistical differences were obtained in the numbers of eggs between the two groups of puppies (p > 0.05).
Viability of STHs in canine feces
In the control group (G-1), viability of eggs of STHs did not show changes after 28 days. As given in Figure 2, the percentages of viable eggs of T. canis ranged from 88% to 94%, and similar results were obtained for T. leonina (87–94%) and T. vulpis (83–93%).
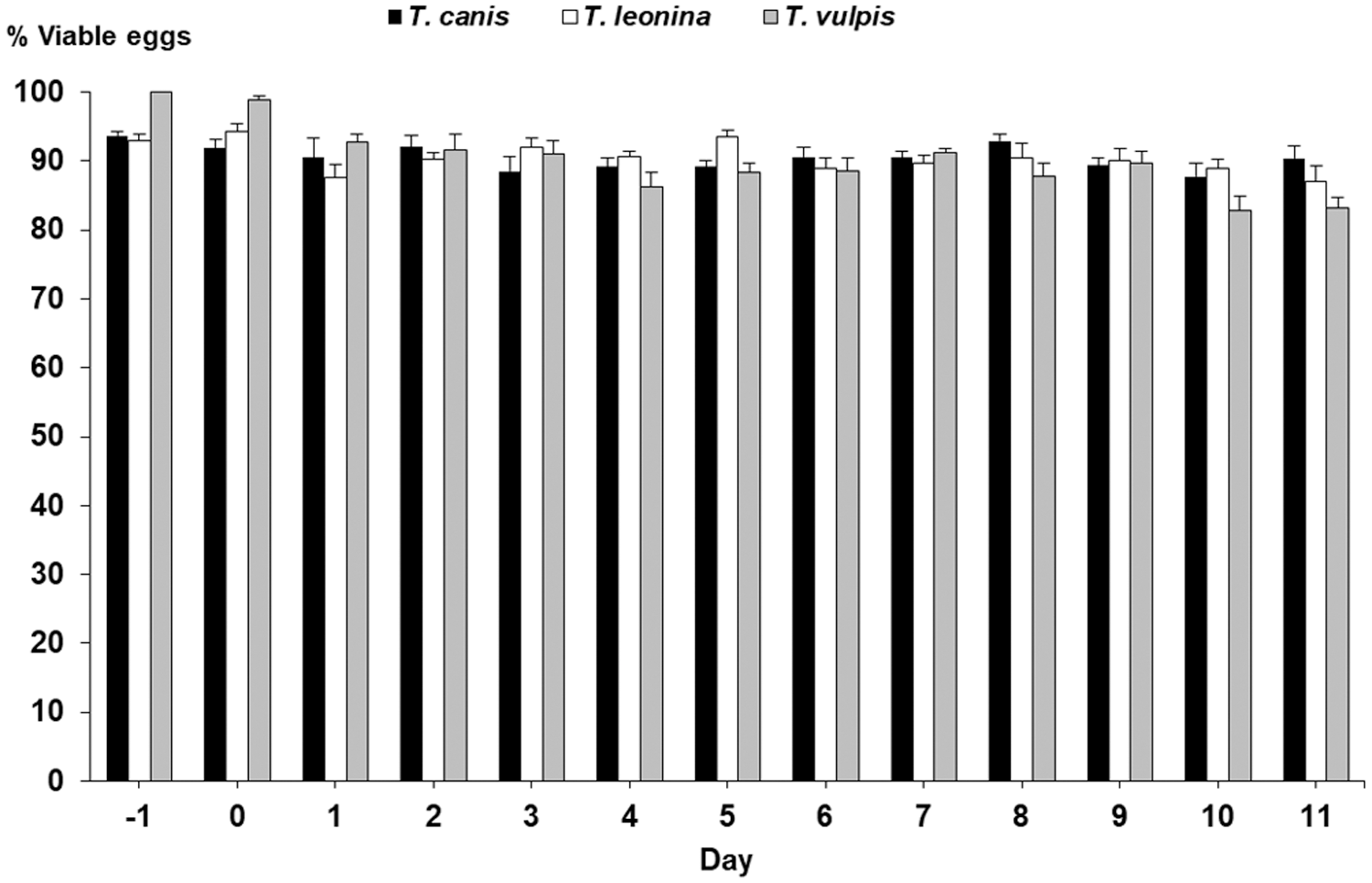
Percentages of viable eggs of Toxocara canis, Toxascaris leonina, and Trichuris vulpis in the feces of G-1 (control puppies) after 28 days.
The percentages of eggs of T. canis viable in the feces of the puppies given fungal spores (G-2) were reduced to one third after 28 days (Fig. 3). Viability of fecal eggs of T. leonina dropped to 36–50% and that of T. vulpis by 40–50%. Significant differences were observed between G-1 and G-2 for the eggs of T. canis (Z = −9.812, p = 0.001), T. leonina (Z = −9.991, p = 0.001), and T. vulpis (Z = −8.861, p = 0.001).
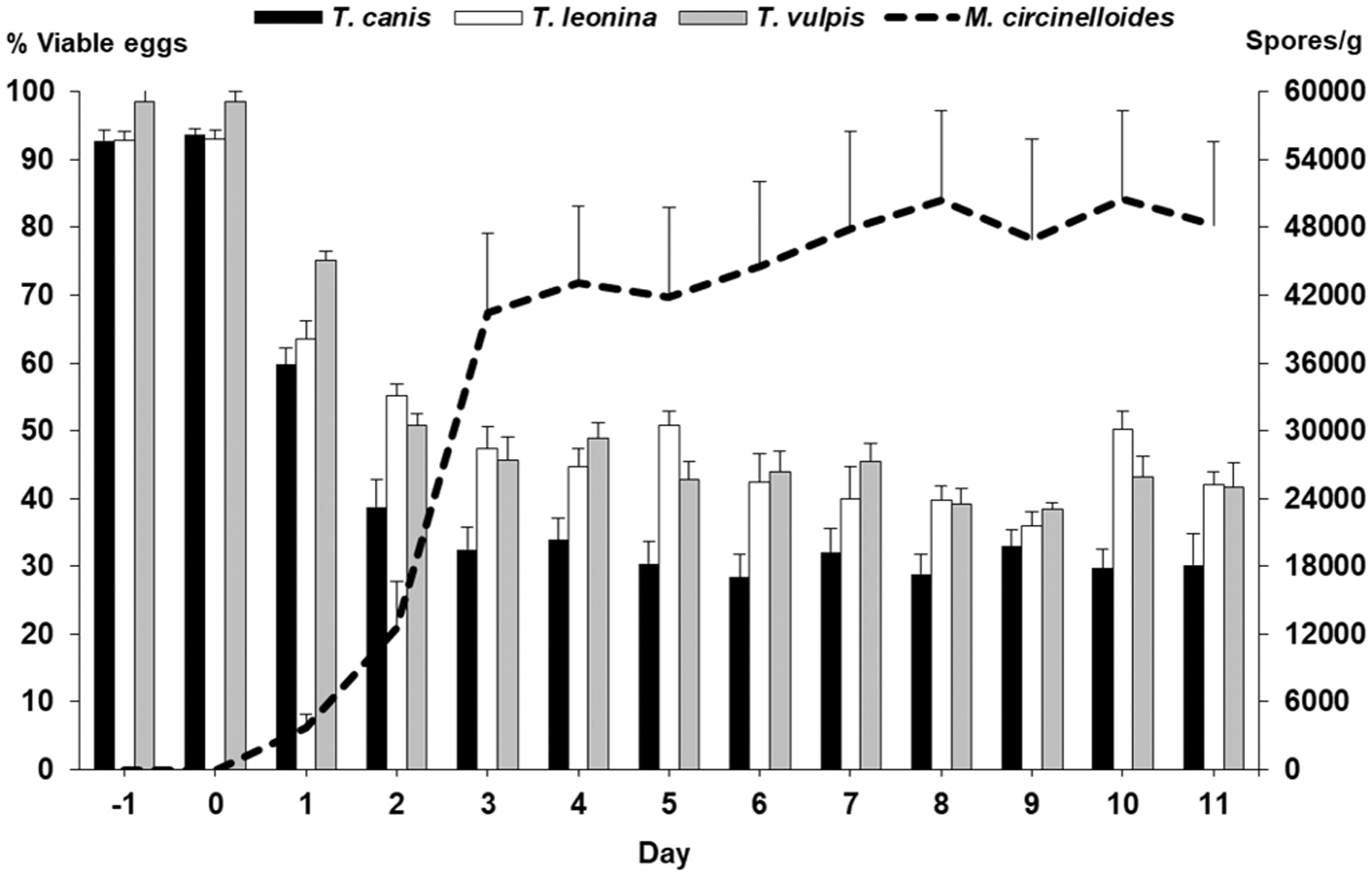
Mean numbers of spores of Mucor circinelloides per gram of feces in G-2. Percentages of viable eggs of T. canis, T. Leonina, and T. vulpis 28 days after providing spores of M. circinelloides to the pups from G-2.
In G-1, the counts of third-stage larvae of A. caninum at day 28 of assay oscillated between 72 and 117 numbers of L3 larvae per gram of feces (LPG) (average = 72 ± 22) (Table 1). Numbers between 8 and 81 were obtained in the feces of G-2, with an average of 23 ± 9 LPG, which means a reduction of 68%. Significant differences were demonstrated between the two groups (Z = −3.441, p = 0.001).
Mean Numbers of Chlamydospores of Duddingtonia flagrans in Feces of G-1
Mean counts of third-stage larvae of Ancylostoma caninum 28 days after the collection of the feces.
G-1, puppies provided spores of Duddingtonia flagrans; G-2, control puppies.
Morphological damage on STHs exposed to the fungi
Twenty-eight days after the addition of fungal spores to the feces of G-2, hyphae of M. circinelloides attached to eggs of ascarids and destruction of the inner embryo were observed (Fig. 4); damaged larvae (Fig. 5) and eggshell breakings were also detected (Fig. 6). Injury of the polar plugs, disappearance of the embryo, and vacuolization of eggs of T. vulpis were also recorded (Fig. 7).

Egg of T. canis with hyphae attached to the shell and destruction of the inner embryo 28 days after giving spores of M. circinelloides to the pups in G-2.

Larvae inside the eggs of STHs can result in permanent injured when exposed to M. circinelloides.

Eggshells of ascarids broken by the action of M. circinelloides in the feces of infected puppies receiving spores of the fungi (G-2).

The polar plugs in eggs of T. vulpis were removed by hyphae of M. circinelloides in the feces of G-2.
Presence of fungal spores in feces
Spores belonging to the fungi M. circinelloides or D. flagrans were not observed in the feces of the control puppies (G-1). In group G-2, the spores of M. circinelloides and D. flagrans were first observed 1 day after beginning the administration of premixed dry feed (Fig. 3, Table 1). The counts of spores of M. circinelloides increased significantly from the third day and constant values around 40,000–50,000 spores were observed until day 11 (Fig. 3). As summarized in Table 1, the numbers of chlamydospores of D. flagrans in the feces of the puppies of G-2 rose from 1 day after starting the administration of the premixed feed, and ranged between 516 and 10,153 chlamydospores per gram feces.
Adverse effects
Pups belonging to G-2 did not refuse to take the dry feed mixed with the blend of fungal spores. Examination of the puppies did not reveal alterations regarding digestive activity, respiratory system, or the skin. No differences were observed in relation to G-1.
Discussion
Among the STHs, T. canis, T. vulpis and A. caninum are parasites affecting domestic and wild carnivores, including humans. Infection by STHs occurs mainly through the exposure to parasitic stages (ingestion of eggs or larvae or percutaneous invasion of third-stage larvae of A. caninum) that became infective after a variable period in the soil. Humans can be infected by accidental contact with these helminths that reinforces the zoonotic risk associated to animal feces.
To limit the survival of the STHs in the feces of dogs, spores of two filamentous fungi with antagonistic effect on eggs (M. circinelloides) and larvae (D. flagrans) were mixed with dry feed before the administration to puppies. The finding of fungal spores in fecal samples 1 day afterward indicates the usefulness of this formulation to ensure the presence of parasiticide fungi in the feces, and their ability to resist the gastrointestinal tract, in agreement with Carvalho et al. (2009). Analysis of the feces 28 days later revealed a significant reduction (by half at least) of the viability of eggs of T. canis, T. leonina, or T. vulpis, confirming that fungi retained their ovicide effect. Previous studies reported successful results through hand spraying spores of M. circinelloides, Trichoderma spp., or Verticillium spp. directly on fecal pats of dogs and pigs infected by T. canis and Ascaris suum, respectively (Arias et al. 2013, Cortiñas et al. 2015). Silva et al. (2010) found 29.5–94.8% reduction of viable eggs of T. vulpis in Petri plates. Furthermore, in this investigation, the numbers of third-stage larvae of A. caninum were reduced by two thirds in the feces of puppies given the fungal spores, underlining that the high activity of nematophagous fungi D. flagrans develops against larvae of A. caninum (Maciel et al. 2010).
As puppies born are frequently infected by certain STHs, high counts of eggs can be passed in their feces after a minimum period of 2–3 weeks after birth (Traversa et al. 2014), exacerbating the environmental contamination. One possible solution could rely on the appropriate deworming of puppies from 2 weeks after birth and then repetitive treatments at suitable intervals to preclude the transmission of larvae through milking (Overgaauw and Van Knapen 2013). For instance, pups should receive parasiticide therapy at 2, 4, 6, and 8 weeks of age, then monthly treatments up to 6 months (ESCCAP 2017). In the current assay, the examination of feces of 3-month-old puppies that had been dewormed 2 months before with fenbendazole and two and a half months with pyrantel pamoate showed high levels of eggs of T. canis, T. leonina, T. vulpis, and A. caninum. These results seem to indicate that puppies are maintained in a high-risk area of infection by STHs, and emphasize the need to develop helpful strategies against the parasitic stages in the ground.
Different measures have been advised to reduce the environmental contamination by parasites, consisting in improving education of public and pet owners, hygiene, avoiding pets to defecate in public places, cleaning up of feces from soil and restriction of free-roaming pets to public areas (Traversa, 2012). The application of covering sandpits with a canvas every time it remains unoccupied for avoiding pets defecate in public zones appears possible in small areas only, as occurs with the regular removal of feces (Vanhee et al. 2015). Pouring chemicals, hot water, or even fire directly on fecal pats have been counseled to destroy eggs of STHs as Baylisascaris procyonis, the ascarid affecting raccoons, coatis, and even people (Blizzard 2010); these strategies can be difficult to apply and harmful to the environment. In the kennel where the present investigation was carried out, feces are never removed from the soil in the outdoors runs, and fences block the entry of free-roaming animals, but the possible role of paratenic hosts should not be discarded (Epe 2009).
One of the most common situations associated with the possible transmission of STHs occur between pregnant females and their progeny. Adult infected females shed eggs of STHs that contaminate the environment where the offspring is maintained. In addition, vertical transmission (transplacental or lactogenic) of T. canis takes place between bitches and puppies, and ascariosis can occur also through the ingestion of larvae in paratenic hosts (Lee et al. 2010, Schnieder et al. 2011). Bitches are a major source of infection for their offspring because they can harbor somatic larvae for many years, which mobilize during pregnancies and infect next litters. As a result, puppies are frequently born infected by T. canis or A. caninum (Overgaauw and Van Knappen 2013). Under this situation, deworming is often considered as the most reasonable strategy in dog-breeding centers or even in kennels (Traversa et al. 2014). With the aim to prevent transmission of helminths from bitches to pups, there are counseled protocols comprising the daily administration of fenbendazole to bitches between day 40 of pregnancy and day 14 postpartum, or macrocyclic lactones on days 40 and 55 of pregnancy (ESCCAP 2017). In the kennel where the present investigation was carried out, pregnant or lactating bitches do not receive additional deworming, and all adult dogs are dewormed every 3 months to avoid shedding of parasites eggs during the puppyhood also. It has been stated that the overall egg output of T. canis could be lessened by 29% if 90% of them receive therapy four times a year (Nijsse et al. 2015).
Eggs of STHs shed in shadowed areas with vegetation and humidity require a variable period to attain the infective stages, fertilized eggs (T. canis, T. leonina, T. vulpis) and third-stage larvae (A. caninum). This is a crucial point when considering fungi as parasite antagonists, because these organisms should act before the infective phase is reached. The period that has been stated is >3 weeks for T. canis and T. leonina and T. vulpis, and ≥2 weeks for third-stage larvae of A. caninum (De Mello et al. 2014). In this study, the effect of the fungi was measured 28 days after the administration of spores to the puppies, and viability and counts of ascarids, trichurids, and ancylostomids were significantly reduced, whereas adverse effects among puppies were not recorded. In accordance, it has been concluded that mixing a blend of spores of M. circinelloides and D. flagrans with dry feed represents an innocuous, innovative, sustainable, and successful approach to reduce the risk of soil contamination by STHs as T. canis, T. leonina, T. vulpis, and A. caninum. A long-term investigation is in progress to confirm the preliminary results obtained in the present research.
Footnotes
Acknowledgments
This study was partly supported by the Research Project CTM2015-65954-R (Ministerio de Economía y Competitividad, Spain; FEDER). Dr. María Sol Arias Vázquez is the recipient of a Ramón y Cajal (Spanish Ministry of Economy and Competitiveness) contract. Cristiana F. Cazapal-Monteiro is the beneficiary of a postdoctoral research fellowship (Xunta de Galicia, Spain).
Author Disclosure Statement
All authors have approved the final version of the article and certify that they have no affiliations with or involvement in any organization or entity with any financial interest in the subject matter or materials discussed in this article.