
Abstract
Schmallenberg virus (SBV) is an orthobunyavirus in the Simbu serogroup that emerged in Germany in late 2011 and was mostly associated with a mild transient disease of sheep and cattle. SBV is transmitted by biting midges (Culicoides species) and causes abortions, stillbirths, and congenital defects in naïve pregnant ruminants. Two separate studies were conducted with a primary objective of better understanding the virological and serological responses of sheep and cattle to different SBV isolates after experimental infection. The second objective was to produce immunoreagents and challenge materials for use in future vaccine and diagnostics research. These studies were carried out using the following infectious inocula: (i) infectious serum (IS) (ii) cell culture-grown virus, and (iii) infectious lamb brain homogenate. The responses were assessed in both species throughout the course of the experiment. SBV RNA in serum (RNAemia) was detected as early as 2 (in sheep) and 3 (in cattle) days postinfection (dpi) and peaked on 3 and 4 dpi in cattle and sheep, respectively. Cattle had higher levels of RNAemia compared with sheep. Experimental infection with IS resulted in the highest level of RNAemia in both species followed by cell culture-grown virus. A delayed, low level RNAemia was detected in cattle inoculated with infectious sheep brain. Isolation of SBV was only possible from 4 dpi sera from all cattle inoculated with IS and one sheep inoculated with cell culture-derived virus. SBV neutralizing antibodies were first detected on 14 dpi in both species. No specific gross and microscopic lesions were observed in either study. In conclusion, these studies highlight not only the difference in viremia and anti-SBV antibody level against the different SBV isolates, but also the extent of the response in the two host species.
Introduction
A
Though, there are strong regulatory and control mechanisms to prevent the introduction of foreign animal diseases into the United States, the extensive trade and travel between the United States and Europe along with the presence of both an SBV susceptible ruminant population and competent vectors (Smith et al. 1996) make Schmallenberg disease a potential threat to United States agriculture. Therefore, development of detection methods and vaccines for the control and prevention of SBV infection would provide animal health practitioners with important tools to manage cases of potential SBV introduction. Until now, subcutaneous inoculation of cattle, sheep, or goats with infectious serum (IS) or cell culture-grown virus followed by monitoring of clinical signs and viremia has been the model for studying SBV pathogenesis (Wernike et al. 2012, 2013b, Laloy et al. 2015). However, in these studies there have been no direct comparisons of the use of infected brain homogenate as the inoculum instead of IS or cell culture-derived virus. Furthermore, viremia is measured indirectly, extrapolated from reverse transcription–quantitative PCR (RT-qPCR) Ct values. Therefore, the primary objective of these studies was to better understand the virological and serological responses of local sheep and cattle breeds to different SBV isolates after experimental infection. A secondary objective was to produce immunoreagents and challenge materials for use in future vaccine and diagnostics research.
Materials and Methods
SBV inoculum
SBV (BH80/11-4) was isolated from the blood of a field-infected cow (103 plaque-forming units [pfu]/mL titer) (Hoffmann et al. 2012) and kindly provided by M. Beer, FLI (Insel Riems, Germany). This isolate was passaged once in the Aedes albopictus cell line C6/36 (ATCC CRL-1660) at 37°C and 5% CO2 for 5 days to achieve a titer of 105 pfu/mL before animal inoculation. The isolate was called infectious cell culture supernatant (ICCS). SBV infected lamb brain (RT-PCR positive [Ct = 30], virus isolation negative) designated as infectious sheep brain homogenate (ISBH) was kindly provided by Falko Steinbach, Animal Health and Veterinary Laboratories Agency (Surrey, United Kingdom). The infectious bovine serum (IS) was produced by inoculation of calves with blood samples from diseased cows of the initial outbreak series (Wernike et al. 2012) and had a titer of 103 pfu/mL. Sequencing revealed that the IS and ICCS inocula contained the same SBV isolate (data not shown).
Animals and experimental design
Experiments involving animals and viruses were performed in accordance with the Federation of Animal Science Societies Guide for the Care and Use of Agricultural Animals in Research and Teaching, the USDA Animal Welfare Act and Animal Welfare Regulations, and were approved by the Kansas State University Animal Care and Use Committees and Institutional Biosafety Committees.
In the first study, 10 Charollais sheep were assigned to three groups with each group receiving one of the following: IS (n = 4), ICCS (n = 5), or phosphate-buffered saline (PBS) (n = 1). In the second study, nine Holstein calves were assigned to three treatment groups: IS (n = 4), ICCS (n = 3), and ISBH (n = 2). All animals were inoculated subcutaneously with 1 mL of inoculum except for the ISBH, which was administered at a 3 mL dose. At 14 days postinfection (dpi), cattle #7 (ICCS inoculated) and #10 (ISBH inoculated) were boosted with IS, and #9 (ISBH inoculated) was boosted with ICCS. Animals were monitored for clinical signs including rectal temperature daily until 7 dpi. Blood samples were collected from both sheep and cattle on 0, 1, 2, 3, 4, 5, 6, 7, 14, and 21 dpi. Some animals were euthanized on 4 and 5 dpi based on RNAemia level and the remaining animals (two sheep and three cattle) were kept until the end of the experiment (21 dpi) to monitor RNAemia and seroconversion. Tissues were collected during necropsy on 4, 5, and 21 dpi (Fig. 1).
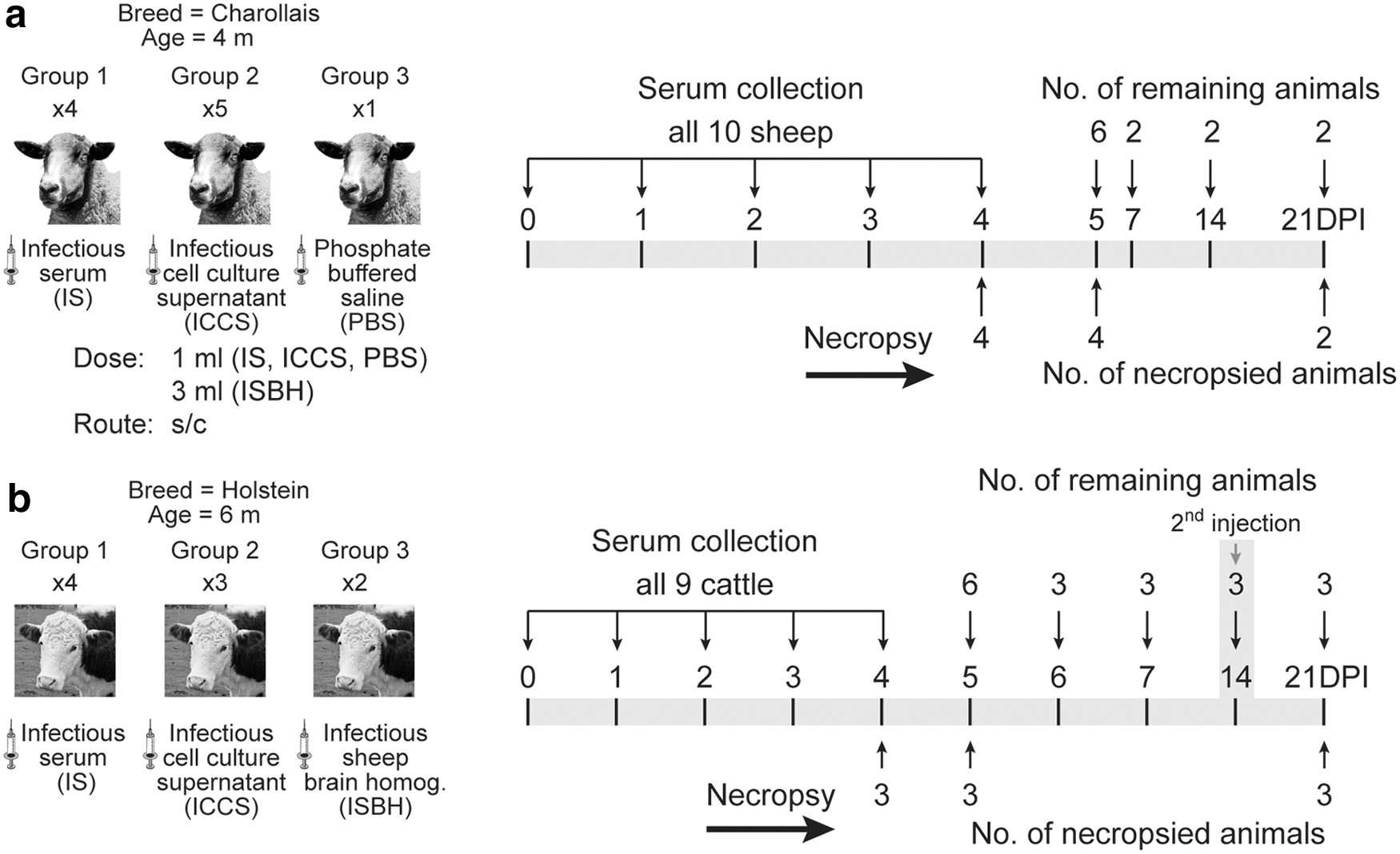
Experimental design.
Real-time RT-PCR
RNA was extracted from 250 μL of serum samples using the KingFisher automated magnetic particle processor (Thermoscientific, MA) and MagMAX RNA extraction kits (Ambion, Inc., TX) according to the manufacturers' recommendations. The SBV genome load was determined by an S segment-specific, real-time RT-qPCR as described previously (Bilk et al. 2012). The RT-qPCR assay was optimized with the AgPath-ID One-Step RT-PCR Kit (Applied Biosystems, CA) using 5 μL RNA template in a total reaction volume of 25 μL and 42 cycles of amplification. A cutoff value of 35 was set, based on the Ct value of the pre-SBV inoculation serum.
Virus isolation/titration
The viral load in serum from calves and sheep was determined by plaque assay as described previously with minor modifications (Mansfield et al. 2013). Briefly, instead of methyl cellulose, 2.4% microcrystalline cellulose Avicel RC (FMC BioPolymer, PA) mixed with an equal volume of 2 × minimum essential media, supplemented with 10% fetal calf serum (FCS) and 1% antibiotics was used as an overlay. Cells were fixed and stained, and the number of plaques was counted to calculate virus titer as pfu/mL.
Plaque reduction neutralization test
SBV neutralizing antibody titers in serum samples collected from cattle and sheep were measured using a plaque reduction neutralization test (PRNT) as previously described (Mansfield et al. 2013); with the minor modification mentioned above. Titers were expressed as the reciprocal of highest serum dilutions yielding ≥50% reduction in the number of plaques (PRNT50).
Immunofluorescence assay
To further assess host serological responses, an immunofluorescence assay (IFA) was performed using the antisera obtained from the experimentally infected cattle and sheep. Vero-E6 cells were grown in eight-well chamber slides for 24 h at 37°C and 5% CO2, or until cells reached >90% confluency, then infected with cell culture-derived SBV with a multiplicity of infection (MOI) of 1. After 1 h incubation at 37°C and 5% CO2, inoculum was aspirated and replaced by 1 mL of Dulbecco's minimum essential media with 10% FCS. Slides were incubated at 37°C and 5% CO2 for 24 h, then the media removed and wells washed once with 1× PBS. The cells were fixed with 10% neutral buffered formalin for 15 min followed by three PBS washes, permeabilized with 0.1% Triton-X100 diluted in 1× PBS for 5 min followed by two PBS washes and blocked for 1 h at room temperature with 3% normal goat serum. Anti-SBV polyclonal antibody from 21 dpi sheep and cattle, diluted 1:50 in 1× PBS, was added and the slides incubated for 1 h at 37°C and 5% CO2. After washing the slides three times with 1× PBS, goat anti-bovine immunoglobulin G (IgG)-fluorescein isothiocyanate conjugate (Santa Cruz Biotechnology, Inc., TX) diluted to 1:200 in 1× PBS was added to each well. Slides were washed three times with 1× PBS and protected from light exposure until examined on an inverted epifluorescence microscope (Nikon eclipse TE2000-S; NY).
Western blot analysis
The serological response was further evaluated by western blot assay. SBV antigen was prepared by growing the virus in Vero-E6 cells infected with an MOI of 1. The cell lysate was then centrifuged at 13,000 g for 15 min and supernatant was collected in a sterile tube and stored at −80°C until further analysis. The proteins were separated using a 12% Bis-Tris gel (Life Technologies) and transferred to a polyvinylidene fluoride membrane (Millipore, MA). The membrane was blocked with 0.05% Tween-20 in 1× PBS (pH 7.4) containing 3% bovine serum albumin for 1 h, then incubated overnight at 4°C with the study-produced bovine anti-SBV hyperimmune serum (Calf #7, 21 dpi) at a 1:50 dilution in blocking buffer. After washing the membrane with 1× PBS, goat anti-bovine IgG (H + L)-horseradish peroxidase conjugate (Thermoscientific, IL) diluted 1:2000 in blocking buffer was applied for 1 h at 37°C. After washing the membrane with 1× PBS, antibody binding was visualized with a 4-chloro-1-naphthol and H2O2 (Fluka, WI) reaction.
Enzyme-linked immunosorbent assay
To measure the anti-SBV antibody response in sheep and cattle, an indirect enzyme-linked immunosorbent assay (ELISA) based on baculovirus-expressed SBV nucleoprotein was used according to a previously described protocol (Breard et al. 2013, Faburay et al. 2014). The cutoff point was determined by the addition of three standard deviations to the corresponding mean optical density (OD) value of the prevaccination serum. Mean OD values equal to or greater than the cutoff value were considered positive.
Pathology
Samples of the following tissues were collected during necropsy and placed in 10% neutral buffered formalin: heart, lung, liver, kidney, adrenal, spleen, sections of the gastrointestinal tract, pancreas, mesenteric lymph node, skeletal muscle, brain, ovary or testicle, thyroid, thymus, and haired skin. For both studies these tissues were trimmed, processed, and embedded in paraffin using standard processes by the Kansas State University, Veterinary Diagnostic Laboratory Histology Laboratory. Tissues were sectioned at 4 μm, stained with hematoxylin and eosin and reviewed by a veterinary pathologist.
Results
Clinical observation
No clinical signs or increased rectal temperatures were observed in sheep. A single calf that was inoculated with the IS was pyrexic, 103.1°F and 103.4°F, on 3 and 4 dpi, respectively. All other infected calves had normal rectal temperatures ranging from 100.4°F to 102.8°F. No additional clinical responses associated with the experimental infections were noted.
Viremia/RNAemia level
Viral RNA was first detected in blood by RT-qPCR on 2 dpi in sheep and on 3 dpi in cattle, and was detectable until 5 and 7 dpi in sheep and cattle, respectively (Fig. 2). The level of RNAemia peaked on 3 and 4 dpi in cattle and sheep, respectively (Fig. 2). Based on absolute Ct values, animals inoculated with the IS had higher levels of RNA compared with those inoculated with cell culture-grown virus, which in turn had higher levels compared with those inoculated with the infectious brain homogenate. Additionally, the onset of RNAemia in calves inoculated with the infectious brain homogenate was delayed by 2 days. Despite daily attempts from 1 to 7 dpi with all cattle and sheep serum samples, virus was only isolated from 4 dpi sera from the four IS inoculated calves, and one sheep infected with cell culture-derived virus (Table 1). Calf #5 had the highest RNAemia level with a Ct of 22 and the highest isolated virus titer of 1.8 × 104 pfu/mL (Table 1). The SBV genome was also detected in various tissues by RT-qPCR in both species (Table 2). Liver from 5 out of 10 sheep, and 4 out of 9 of the calves were found RT-qPCR positive on day 4 and 5 postinfection. RNA positive brain tissues were detected in 7 out of 10 sheep and 3 out of 8 cattle. Spleen was found positive in a significant portion of the calves (four out of nine); however, spleen from sheep were not collected during necropsy.
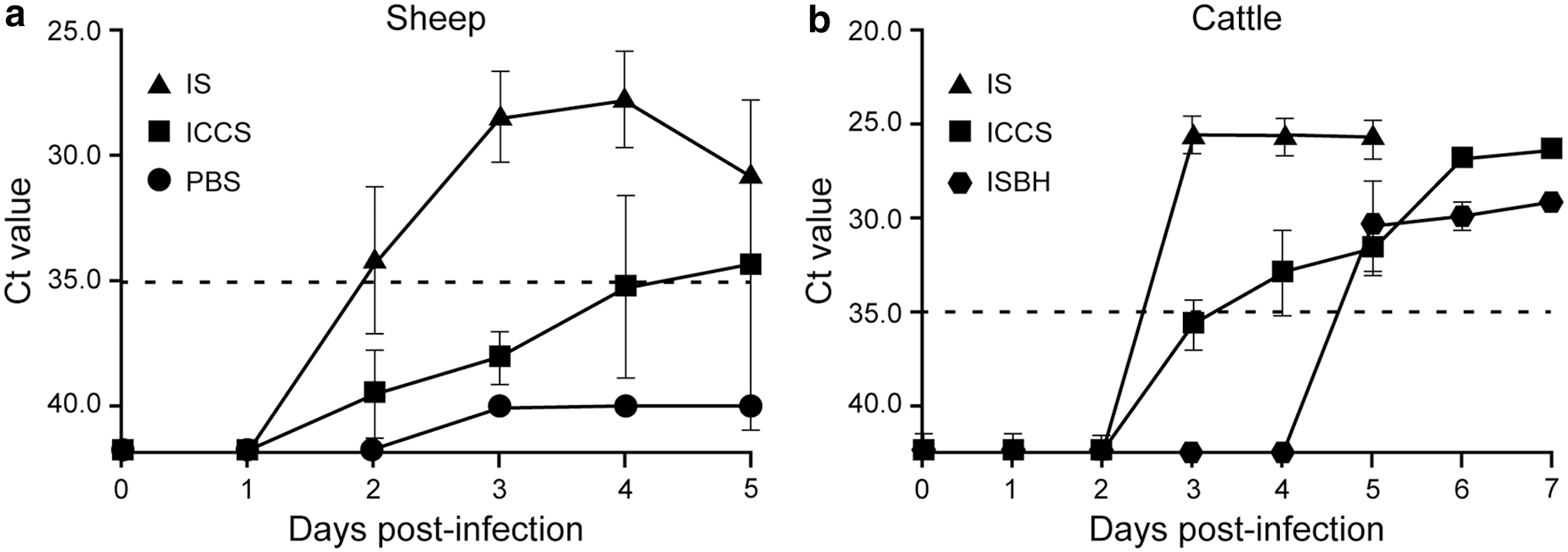
Detection of SBV RNA in serum by reverse transcription–quantitative PCR after inoculation of
Schmallenberg Virus Titers in Four Days Postinfection Sera
S, sheep; C, cattle; IS, infectious serum; ICCS, infectious cell culture supernatant; PBS, phosphate-buffered saline; ISBH, infectious sheep brain homogenate; pfu, plaque-forming units.
Detection of Schmallenberg Virus RNA in Tissues by Reverse Transcription–Quantitative PCR
ND, not detected; NC, tissue not collected.
Serology
The antibody response in sheep was measured by a SBV nucleoprotein-based in-house indirect ELISA, which had high background across all samples including baseline controls. Despite this, a slight antibody level increase was visible at 21 dpi (Fig. 3a). In contrast, the calf sera had minimal background and a five-fold higher response was observed at 21 dpi compared with 0 dpi for calf #7 that received cell culture-grown virus; whereas calves #9 and #10 both inoculated with SBV-positive brain homogenate failed to seroconvert (Fig. 3b). Sheep #9 exhibited a low neutralizing antibody response, as measured by PRNT50, with a titer of 32 at 21 dpi. The rest were negative (Table 3). In contrast, neutralizing antibody titers of 1024, 16, and 256 were detected in the 21 dpi serum from calf #7, #9, and #10, respectively (Table 3). Neutralizing antibodies in cattle were detected only in serum collected after booster injection at 14 dpi, since one of the objectives of this study was to produce serological test reagents that could be used in subsequent studies. Only the cattle serum was able to detect SBV infection on infected Vero cells by IFA. A diffuse fluorescence was observed in the cytoplasm of infected cells but not in un-infected cell controls (Fig. 4a). Similarly, infected cell lysates were analyzed by western blot assay and the two major SBV proteins, nucleocapsid (N) and glycoprotein C (Gc) were detected at 48 h postinfection (Fig. 4b). Both, the recombinant N and Gc protein were detected by the day 21 cattle serum (Fig. 4c, d).
Anti-SBV nucleoprotein antibody responses. Host immunoglobulin was measured by indirect enzyme-linked immunosorbent assay against SBV nucleoprotein.
Performance of generated cattle SBV antiserum in different serological assays.
Schmallenberg Virus Neutralizing Antibody Titers in Sera at Different Days Postinfection
Pathology
Despite extensive review of the tissues collected from all animals in both studies, no lesions directly attributable to SBV were observed. However, in three calves, there were some central nervous system (CNS) lesions that are nonspecific for a variety of viruses, or other pathogens. In the brain stem of calf #4 there was a focal area of gliosis and another region with hypercellularity. Additionally, calves #6 and #7 both had rare foci of cerebral perivascular cuffing with mononuclear cells but no evidence of vasculitis.
Discussion
The experimental infection with SBV, regardless of the type of inoculum, did not cause any observable clinical signs in either of the animal species evaluated. One calf had pyrexia on 3 and 4 dpi. Similarly, with the exception of three calves that had low numbers of focal changes in their brain parenchyma that could be attributable to a viral etiology, there were no histopathological findings attributable to SBV. Immunohistochemistry for viral antigen was attempted on these tissue sets but was unsuccessful, likely due to the absence or low levels of viral antigens present post formalin-fixation, processing and paraffin-embedding. These results are consistent with Wernike et al. (2012, 2013b) and Poskin et al. (2014) who observed a short RNAemia in both species and no clinical signs, except diarrhea in individual animals, and gross lesions.
The onset of RNAemia after experimental infection was found to be dependent on the inoculum type. RNAemia was detected 2 days earlier in both the IS and ICCS inoculated calves compared with animals inoculated with the infectious brain homogenate. Wernike et al. (2013a) did not detect RNA in the serum from sheep that were inoculated with virus (108 median tissue culture infective dose [TCID50]/mL) originating from lamb's brain and passaged three times in Vero cells before use. The detection of RNA in the serum from the ISBH inoculated cattle, in this study, could be explained by either the species difference (cattle vs. sheep) or the inoculum itself, since in this experiment virus from lamb brain homogenate was used without prior cell culture passage. Overall, the IS inoculation resulted in higher RNAemia compared with the ICCS or the infectious brain homogenate in cattle as seen previously (Wernike et al. 2013b). Additionally, detection of viral RNA in the spleen, liver, and brain samples was consistent with previous reports (Bilk et al. 2012, Wernike et al. 2012, 2013b).
Wernike et al. (2013b) detected anti-SBV antibodies on 10, 14, and 22 dpi in sheep using a commercial ELISA kit. Using an in-house indirect ELISA, we detected higher OD levels in cattle versus sheep sera when testing antibodies reacting to the SBV N protein by 21 dpi (Fig. 3). The same was true for the neutralizing antibody titer as measured by PRNT (Table 3). The higher ELISA and neutralizing antibody titers seen in the two cattle sera are most likely due to these cattle receiving a second inoculation. The high background observed in the sheep sera ELISA may be due to an inherent species effect or prior exposure to other orthobunyaviruses. The N protein is the most conserved protein among orthobunyaviruses and elicits a strong humoral immune response in infected animals (Gonzalez-Scarano et al. 1982). A recent cross-sectional study revealed a Cache valley virus (CVV) seroprevalence rate of 49%, 17%, and 10% in sheep in the eastern, central, and western United States, respectively (Meyers et al. 2015). We ruled out CVV by performing PRNT with the collected sheep sera that showed absence of neutralizing antibodies except sheep #4 (data not shown). However, the high background observed in sheep could be due to the circulation of other unidentified orthobunyaviruses in the sheep population of the region.
The increase in the antibody response in cattle (Fig. 3b) after boost with the IS appears to be dependent on the initial type of inoculum given to the cattle. This increase was observed in the calf primed with the cell culture-derived virus, but not in the calves inoculated with brain homogenate. Since no animal from the IS inoculated cattle group were kept past 5 dpi, we do not know the response in this group. The hyper-immune serum collected from a calf on 21 dpi was used to detect SBV replication in cell culture by IFA, which is more specific than cytopathic effects (Fig. 4a). Similarly, the same serum detected both the SBV Gc and N proteins on the western blot indicating its suitability as a reagent for detection of SBV antigens (Fig. 4b). The size of the two proteins was confirmed with separate western blots using recombinant SBV N (Fig. 4c) and Gc (Fig. 4d) proteins.
Conclusions
SBV has spread throughout most of Europe since its initial detection in 2011 in Germany. However, to date, this virus' emergence in Europe is poorly understood (Claine et al. 2015). Consequently, SBV poses a serious threat to other countries like the United States, which has competent insect vector species (Smith et al. 1996), susceptible livestock populations, and conducts extensive trade with Europe. Although, both cattle and sheep are reported as good experimental animals for researching SBV disease pathogenesis and vaccine efficacy, to our knowledge there is no published information on the response and pathogenesis of SBV infection in local United States sheep and cattle breeds. These studies highlighted not only the difference in viremia and anti-SBV antibody level against the different SBV isolates, but also the extent of the response in the two host species. The virological and serological responses were more apparent in cattle than in sheep. Overall, this work creates an important basis for subsequent development and evaluation of effective countermeasures such as novel vaccines and improved diagnostics.
Footnotes
Acknowledgments
We kindly thank Dr. Martin Beer from Friedrich Loeffler Institut (Germany) for providing us the SBV infectious serum and lab training and Dr. Falco Steinbeck from the Veterinary Laboratory Agency (United Kingdom) for providing the SBV infected lamb brain tissue. We also appreciate Dane Jasperson, Lindsey Reister-Hendricks, and Kruger Bryant for providing assistance in the lab and with animal work. We thank Mya Masterson and Mal Hover at KSU for immunochemistry work and figure formatting, respectively. This research was funded by the Kansas Bioscience Authority and the USDA, ARS, NP-103, CRIS #3020-32000-009-00D.
Author Disclosure Statement
No conflicting financial interests exist.