
Abstract
Biting midges of the genus Culicoides (Diptera: Ceratopogonidae) are the principal vectors of several notable viral pathogens infecting animal livestock. Sickness and animal deaths caused by the Culicoides-transmitted bluetongue virus, as well as the recent Schmallenberg virus outbreak, have threatened the livestock industry in Europe. Recent studies highlight how, in the near future, the application of “dry” fungal conidia of Metarhizium anisopliae in animal shelters and microenvironment (e.g., dung, manure, leaf litter, and livestock surroundings) may be used to control the Culicoides vector, thus, reducing the incidence of Culicoides-borne diseases.
Introduction
Several microorganisms
A few studies have shown the potential of entomopathogenic fungi to control Culicoides biting midges, hereafter referred to as “midges”, vectors of numerous important livestock diseases, including bluetongue, which pose a severe economic risk to the ruminant livestock industry (van Schaik et al. 2008, Velthuis et al. 2010, Zanella et al. 2012, Pinior et al. 2018). The economic impact of the bluetongue serotype 8 (BTV8) epidemics of 2006 and 2007 in the Netherlands alone accounted for 32.4 and 164–175 million, respectively (Velthuis et al. 2010). The recent estimates indicate that a total cost of €41.9 million was invested in the bluetongue virus vaccination and surveillance programs in Austria and Switzerland alone (Pinior et al. 2018).
There are many other fungal entomopathogens, apart from M. anisopliae and B. bassiana, which have been explored for controlling midges by many authors. For example, de Souza et al. (2014) reviewed thoroughly and gave a detailed account of fungal and oomycete parasites of chironomids, ceratopogonids, and simulids. The naturally occurring Oomycete fungal pathogen, Lagenidium giganteum, was recorded as biocontrol agent of Culicoides molestus larvae, which caused mortality up to 33% in New South Wales, Australia (Wright and Easton 1996). Another dominant marine Oomycete, Halophytophthora species, was reported to colonize both living and dead pupae of Culicoides subimmaculatus in coastal waters of Hervey Bay region in Queensland, Australia (Stephen and Kurtböke 2011). Yet another deuteromycete fungus, Culicinomyces clavisporus, was highlighted as the potential biocontrol agent against European biting midge, Culicoides nubeculosus larvae (Unkles et al. 2004).
The impact of Culicoides-transmitted viruses such as Akabane in Australia, African horse sickness in Africa, bluetongue (BTV) in North America, Africa, and Europe, as well as recently emerged Schmallenberg livestock disease in Europe, highlight the worldwide importance of midges (Elbers et al. 2013). The wide distribution of infected vector species of midges contributes to the rapid spread of the virus. At least 83 species of Culicoides are found in Europe (Venail et al. 2012); however, only around 30 species have been associated with BTV transmission (EFSA Panel on Animal Health and Welfare 2017). In Europe, Culicoides species that have been implicated as potential vectors of BTV generally belong to the subgenera Avaritia and Culicoides. Culicoides (Avaritia) imicola, C. (Avaritia) obsoletus, and C. (Avaritia) scoticus are presently considered confirmed BTV vectors, while C. (Avaritia) chiopterus, C. (Avaritia) dewulfi, C. (Culicoides) pulicaris, and C. (Culicoides) punctatus as probable vectors (Purse et al. 2015, Foxi et al. 2016).
Current surveillance measures and control programs focus on quarantine or movement restrictions of livestock during periods of insect activity, as well as animal vaccination (Racloz et al. 2006, OIE 2013, Collins et al. 2016, EFSA Panel on Animal Health and Welfare 2017). Where disease control by vaccines is not available, midge control by use of fungal biocontrol agents may play an important role in limiting disease outbreaks. Presently, midge control relies predominantly on synthetic pesticides, which pose a risk to humans and the environment (Carpenter et al. 2008a, Webb et al. 2010, Del Rio et al. 2014, Baker et al. 2015, De Keyser et al. 2017). Climate change models predict warmer and wetter weather, which in turn is expected to lead to larger midge densities (Guis et al. 2012, White et al. 2017). Therefore, safe and effective methods of vector control are urgently needed. The application of entomopathogenic fungi may provide potential eco-friendly alternatives for the reduction of midge numbers and consequent reduction in disease transmission.
Case Studies
Previous research carried by our group involved several of the commercially viable strains of Metarhizium, Beauveria, Isaria, and Lecanicillium (Deuteromycotina: Hyphomycetes) to test the ability of these strains in killing an indigenous C. nubeculosus (Ansari et al. 2010, 2011). Although C. nubeculosus is not a common midge, nor is it considered an important vector species for Schmallenberg or bluetongue viruses, it was used as a model insect in our studies, which was sourced from a colonized line. Ansari et al. (2011) demonstrated the biocontrol potential use of fungal application to different substrates (peat, leaf litter, manure) as the representative resting sites for Culicoides midges to simulate a more accurate estimation of fungal application in livestock microenvironment. Nicholas and McCorkell (2014) obtained 98% reduction in emergence of Culicoides brevitarsis adults by incorporating M. anisopliae conidia to cattle dung. Superior control was achieved as cattle dung serves as substrate for the growth and development of C. brevitarsis. In addition, Narladkar et al. (2015) reported the use of high dose of fungal spores against unknown species of Culicoides larvae (in drainage channel) and adults (resting on cattle shed walls) and claimed LC50 values of 3837 and 2692 mg (108 cfu/g) for M. anisopliae and B. bassiana, respectively.
Different conidial formulations aimed at improving conidial application and consequently ease of use were tested, that is, dry conidia dusted uniformly on each substrate (“dry” formulation) and conidia suspended in 0.03% aq. Tween 80 (“wet” formulation). It was found that conidia attach to the adult midge and infect it by penetrating the cuticle or integument. Once inside the insect, the fungus grows rapidly producing toxins that kill the midges within 24 h (Ansari et al. 2011). Following colonization of the hemocoel, the fungus erupts through the intersegmental sections and produces conidiophores and conidia (Fig. 1). A commercially available strain, M. anisopliae F52 (Met52® G bioinsecticide, currently available for control of black vine weevil, Otiorhynchus sulcatus (Coleoptera: Curculionidae) in horticultural crops) killed 100% of C. nubeculosus within 24 h at 1011 conidia per m2. Furthermore, C. nubeculosus adults exposed to “dry” or “wet” conidia under semi-field condition showed that dry conidia were more effective than wet conidia, causing 100% mortality after 5 days compared to 70%, respectively. Met52 granular formulation is approved in several European countries, but is not available as dry spores or in a powder form for use in midge control.
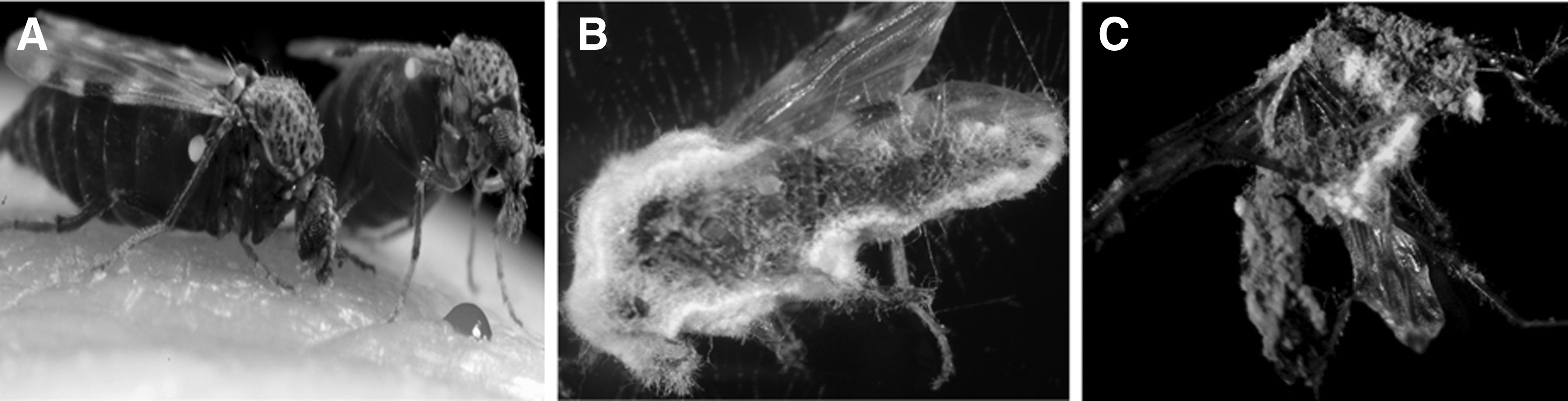
Culicoides nubeculous midges at different periods after contact with dry conidia of the entomopathogenic fungus Metarhizium anisopliae V275.
Irrespective of application method or substrate, all surviving adults collected from M. anisopliae-treated substrates in a greenhouse study died from fungal infection. Midges were observed directly transmitting infective conidia between males and females. Similarly, transmission of M. anisopliae between adult mosquitoes (Anopheles gambiae) has been demonstrated. Further studies in Australia demonstrated the susceptibility of another important species of biting midge (C. brevitarsis) to different strains of M. anisopliae infection (Nicholas and McCorkell 2014). The authors suggest that M. anisopliae has the potential to control C. brevitarsis through either surface treatment or topical application to cattle or through incorporation into fresh cattle dung. They found that the two strains of M. anisopliae were able to cause 70% mortality in adult C. brevitarsis after exposure for 5 days to surfaces treated with ∼0.6 g/m2 of dry conidia. These mortalities increased to 96% and 94% after 7 days. Moreover, they showed that when M. anisopliae spores were incorporated into fresh cattle dung (between 0.25 and 1 g conidia/kg) the emergence of adult C. brevitarsis was reduced by up to 98%.
Importantly, the fungal strains tested pose no obvious risk to humans or the environment (Strasser et al. 2000, Darbro and Thomas 2009). US Environmental Protection Agency (EPA) conducted risk assessment and found that Metarhizium brunneum ( = M. anisopliae strain F52) was not harmful to earthworms or to such beneficial insects as lady beetles, green lacewings, parasitic wasps, honey bee larvae, and honey bee adults (US EPA 2011, Fischhoff et al. 2017). Their production involves relatively low cost and simple technology processes, facilitating the potential for large-scale production.
Currently, resting sites are poorly defined for Culicoides species, and different vector species have different larval habitats and feeding preferences; for example, adult C. brevitarsis prefers grass tussocks (Bishop et al. 1995), whereas C. impunctatus prefers downy birch (Carpenter et al. 2008b); therefore, it is impractical for the widespread application of fungal spores. Zimmer et al. (2013) assessed and recorded several substrates, which serve as suitable breeding sites and microhabitats for the larval development of midges, for example, maize silage residues, cattle dung, ground of flooded meadow, green filamentous algae and underlying substrate, silt from a pond, and ground of hollows. Carpenter et al. (2008b) found high levels of lichen, moss, and liverwort as commonly resting sites of midge adults near downy birch trees. Breeding sites such as cattle dung could provide a means of exposing midge larvae to M. anisopliae in the field through treated dung (Nicholas and McCorkell 2014). However, targeting newly emerged adults before their initiation of blood feeding would be preferable to achieve significant reduction in disease transmission rates.
Another factor is temperature, which is particularly important as the best time to treat the vector populations would be earlier in the season when midge density is still relatively low and few within the population are infected. The limitation of our studies is that the colonized line is adapted to higher temperatures than wild caught midges, which are cold tolerant; therefore, further studies are required to be conducted with the application testing using wild, field-caught midge populations. Thus, currently, control studies have demonstrated for a colonized midge species, which has a limited vector capacity in the potential use of entomopathogenic fungi for the reduction of midge-borne disease in livestock (Ansari et al. 2010, 2011, Nicholas and McCorkell 2014). The success of midge control programs using these fungi requires large-scale field trials in different microclimate conditions to establish the most effective formulations and application methods for the fungal spores.
Footnotes
Acknowledgment
Authors want to acknowledge Eric Denison, Vector-borne Disease Programme, Institute for Animal Health, Pirbright, Surrey, United Kingdom for providing adult C. nubeculosus.
Authors' Contribution
M.A. conceptualized and wrote the article. P.D. and M.W. provided logistic support and additional information. All authors revised the article and approved the final version.
Author Disclosure Statement
No conflicting financial interests exist.